2个小麦株高QTL位点验证及其对产量相关性状的效应分析
2019-04-11邓光兵张海莉梁俊俊余懋群杨武云
李 涛, 陆 炳, 李 俊,邓光兵,张海莉,梁俊俊,余懋群*,杨武云,龙 海
(1. 中国科学院成都生物研究所,四川 成都 610041;2. 中国科学院大学,北京 100049;3. 四川省农业科学院,四川 成都 610066)
【研究意义】小麦(TriticumaestivumL.)作为最重要的粮食作物之一,养活了全球35 %的人口,为人类提供了重要的能量摄入和蛋白质来源[1-2]。然而随着人口增长和可用耕地的减少,粮食危机正在变得越来越严重。因此实现小麦的高产、稳产对于解决全球粮食危机极其重要[3]。【前人研究进展】自20世纪60年代各国在小麦育种中通过引进小麦矮杆(Reduced height,Rht)基因降低小麦株高(Plant height,PH),增加抗倒伏能力使得小麦产量呈直线上升后[4],株高性状受到了国内外育种家的高度关注并做了大量的研究,旨在发现更多控制小麦株高的关键基因。迄今在小麦21条染色体上,已经有超过50个控制株高的数量性状位点(Quantitative trait locus,QTL)被报道[5-10]。其中Gao等基于高密度遗传图谱在周842B×中国春的重组自交系群体中鉴定到了5个株高QTL,分别位于染色体2A、4A、4B、4D和5A[11]。Zhang等在染色体1D、2B、3A、3D、4A、4B、5A和6B鉴定到了8个株高相关QTL[9]。Cui等利用3个中国小麦品种构建的2个重组自交系,在小麦21条染色体上都鉴定到了与株高相关的QTL[12]。此外还有24个能够降低株高7 % (Rht8)~55 % (Rht5)的关键基因被命名为Rht基因,分布于小麦染色体2AS、2BL、2DS、3BS、4BS、4DS、5AL、5DL、6A和7A[13-15]。株高的降低虽然能提高抗倒伏能力和种植密度,却与主要产量性状存在矛盾,既随着株高的大幅度下降,后代往往出现晚熟、早衰、粒瘪等特征,并最终导致产量下降。孙耀中等利用771份株高具有显著差异的材料探究株高与产量性状的关系发现,小麦植株的矮化会导致千粒重和穗粒重的显著降低[16]。Rht5基因被报道在降低株高的同时会严重影响籽粒数(-66 %)[13];Rht2矮化株系导致千粒重降低0.7~5.6 g[17]。此外,RhtB1、RhtD1和Rht12都被报道在降低株高时会对籽粒重产生不同程度的负效应[18]。【本研究切入点】因此,在小麦育种工作中,对于株高的调控必须兼顾其与产量性状的关系,注意综合因素的影响,使株高与产量性状有机结合,以达到高产稳产的目的[16]。陆炳等利用川麦42×川农16重组自交系,基于高密度遗传连锁图谱鉴定到了2个未曾报道的显著株高QTL[19]。其中来自于川麦42的Qph.cib-5A具有17.25 %的表型贡献率,来自川农16的Qph.cibb-7A具有13.18 %的表型贡献率[20]。【拟解决的关键问题】为了能够验证这两个株高显著位点的真实性并解析其与千粒重、穗粒数等产量性状的关系,从而为在小麦育种中的利用提供理论依据,本研究利用从川麦42×川农16重组自交系中衍生出的2个F2群体通过开发KASP标记的方法鉴定Qph.cib-5A和Qph.cibb-7A的真实性,并通过3个四川小麦品种川麦42、川麦39和川农16构建的2个重组自交系群体阐明这2个位点在不同遗传背景中对株高及重要产量性状的影响。
1 材料与方法
1.1 实验材料
本研究利用的实验材料包括2个重组自交系群体和2个F2群体:含127个家系的川麦42×川农16构建重组自交系[19]由四川省农科院杨武云研究员提供;含193个家系的川麦42×川麦39重组自交系(F9:F10)由中科院成都生物研究所农业生物技术中心分子育种实验室构建;川麦42×川农16重组自交系中筛选的株系(RIL-21和RIL-36)通过与2个亲本杂交得到的2个F2衍生群体,其中F2-1为RIL-21与川农16杂交得到的153个F2单株,F2-2为RIL-36与川麦42杂交得到的187个F2单株。
1.2 田间试验设计和性状评价
川麦42×川农16重组自交系群体和川麦42×川麦39重组自交系群体分别于2015-2016、2016-2017年种植于四川双流和什邡(共3个环境);F2-1和F2-2群体于2017种植于双流。田间试验采用完全随机区组设计,每个区组包括3行,每行1.5 m宽,2.5 m长,45粒种子均匀的种植于每一行。收获前进行田间性状调查,每行随机选取5株,调查株高(Plant height, PH)、穗长(Spike length, SL)、千粒重(Thousand kernel weight, TKW)、穗粒数(Grain number spike-1, GNS) 4个性状。其中株高和穗长利用尺子人工测量,千粒重和穗粒数通过万深SC-G拷种仪器得到。
1.3 表型数据分析
利用Microsoft Office excel 2013进行表型数据处理,包括显著性检验,频率分布直方图绘制,均值的计算等。利用R软件进行最佳线性无偏预测值(The best linear unbiased predictions, BLUP)的计算[21]。
1.4 DNA的提取
田间采集小麦幼嫩叶片组织,通过CTAB法提取基因组DNA[22],Thermo ND 2000微量分光光度计检测DNA的浓度和纯度。提取的DNA样本放置于-20 ℃保存备用。
1.5 KASP引物及KASP反应
根据陆炳等鉴定到的与Qph.cib-5A和Qph.cibb-7A紧密连锁的单核苷酸多态性标记(Single nucleotide polymorphism, SNP)开发KASP标记,通过KASP反应鉴定F2群体基因型。KASP反应体系由混合引物,Master Mix和样本DNA组成。其中混合引物由2条末端碱基不同的正向引物和1条共同的反向引物组成,其组分包括40 μl的dd H2O,15 μl的正向引物(100 μmol·L-1)和30 μl的反向引物(100 μmol·L-1);Master Mix来自于LGC(https://www.lgcgroup.com/);DNA通过CTAB法提取。PCR体系为10 μl,包括5 μl的DNA样本(10 ng·μl-1)、5 μl Master Mix和0.14 μl的混合引物。PCR程序为94 ℃预变性15 min;94 ℃变性20 s, 61 ℃退火和延伸1 min,10个循环,每个循环延伸温度降低0.6 ℃;94 ℃变性20 s,55 ℃退火和延伸1 min,26个循环;37 ℃荧光读取1 min。KASP反应所需组分加于96孔板,在CFX ConnectTMReal-Time System 中选取“FAM”和“HEX”2种荧光进行PCR反应并在反应结束后读取终端荧光信号。
1.6 基因分型
F2-1和F2-2群体利用KASP反应进行基因分型;川麦42×川农16重组自交系群体利用小麦90K SNP芯片进行基因分型(陆炳等2017);川麦42×川麦39重组自交系群体利用小麦35k育种家芯片进行基因分型,由北京中玉金标记生物技术有限公司完成。
2 结果与分析
2.1 表型分析
Qph.cib-5A和Qph.cibb-7A位点具有正向效应的等位基因分别来自川麦42和川农16。因此,我们从川麦42×川农16重组自交系中筛选出同时具有这2种基因型的家系RIL21和RIL36,并利用其分别与川农16和川麦42杂交,构建了2个衍生F2群体:F2-1和F2-2。F2-1群体中,Qph.cibb-7A位点与川麦42基因型一致,Qph.cib-5A存在分离;F2-2群体中,Qph.cib-5A位点与川农16一致,Qph.cibb-7A存在分离。亲本和群体种植在同样的环境,以同样的方式进行性状数据测定。表型数据分析显示在F2群体中,亲本之间株高差异大,群体中分布范围广,符合正态分布,具有典型的数量遗传特征。在两个重组自交系群体中,株高、穗长、穗粒数、千粒重也呈正态分布(表1,图1)。根据相关性分析结果(表2),株高、穗长和千粒重在2个重组自交系群体中彼此呈正相关,因此株高的增加对千粒重和穗长的增加具有正效应。此外在川麦42×川麦39重组自交系群体中穗粒数与株高和千粒重呈负相关,与穗长不具有相关性。在川麦42×川农16重组自交系群体中穗粒数与株高和穗长呈正相关,与千粒重不具有相关性。
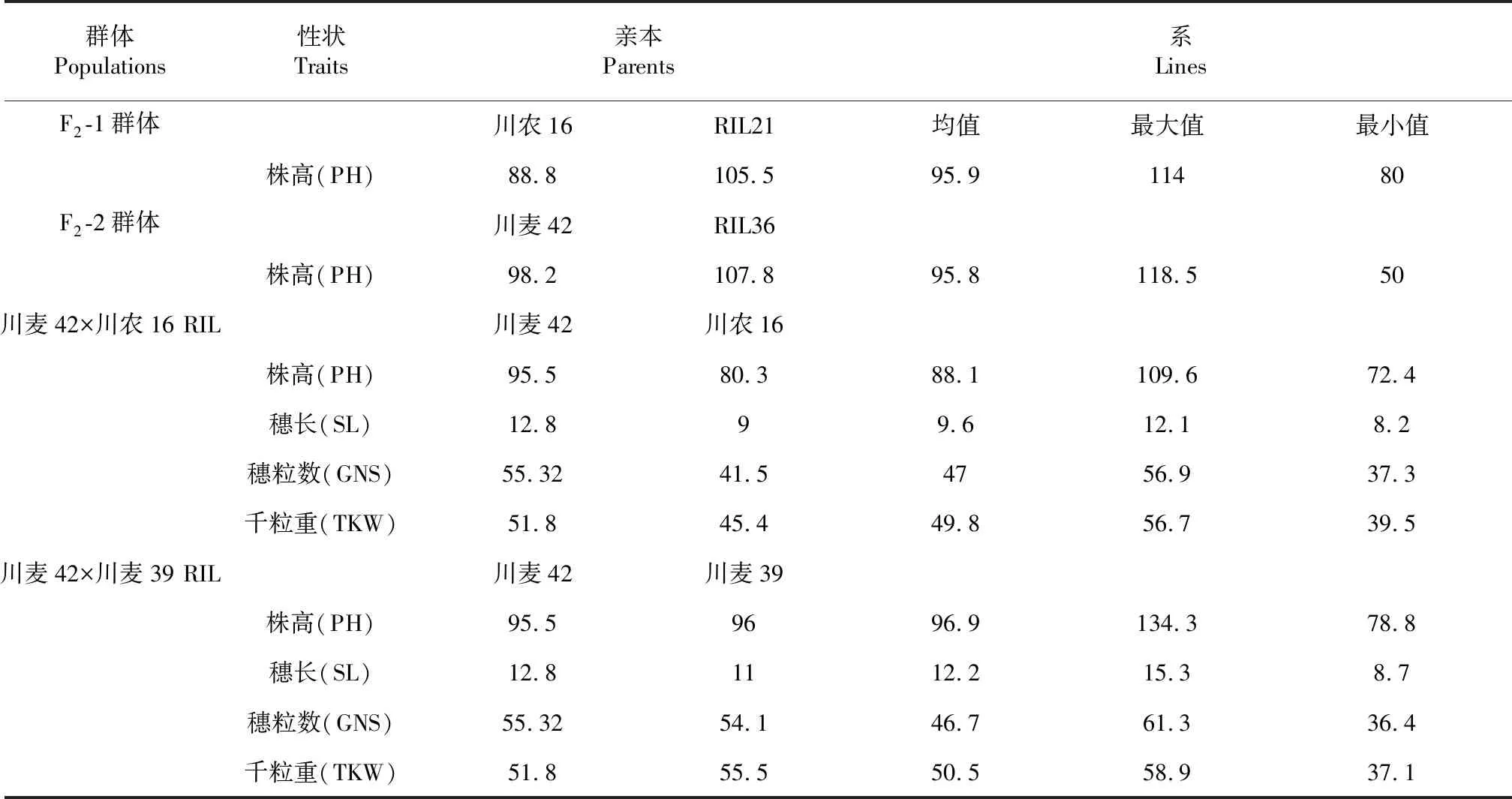
表1 表型性状在群体和亲本中的分布
表22个重组自交系中株高、穗长、穗粒数和千粒重基于BLUP值的相关性
Table 2 Correlation coefficient of plant height (PH), spike length (SL), thousand kernel weight (TKW) and grain number spike-1(GNS) based on the BLUP value in two recombinant inbred line populations
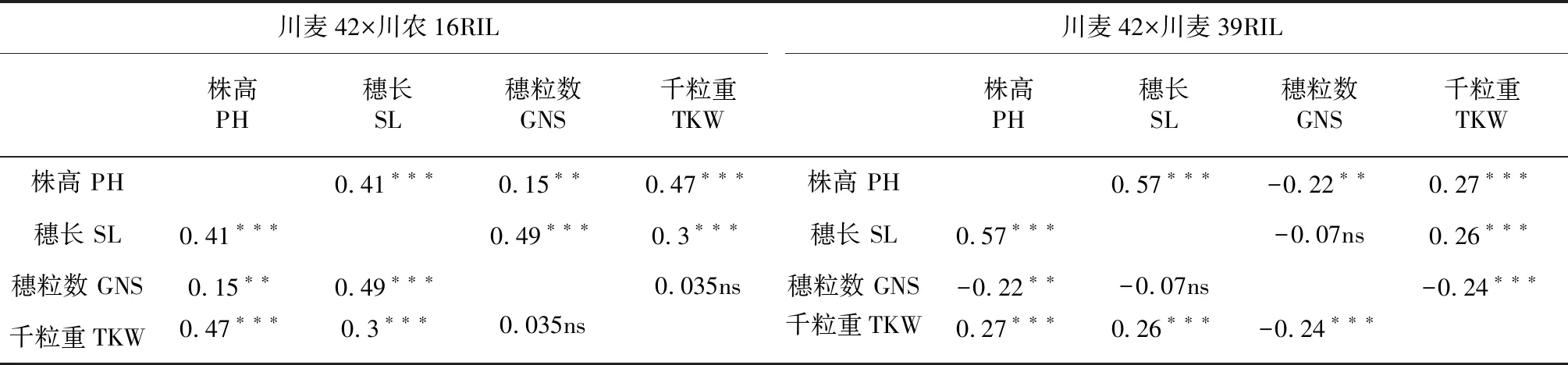
川麦42×川农16RIL川麦42×川麦39RIL株高 PH穗长 SL穗粒数 GNS千粒重TKW株高 PH穗长 SL穗粒数 GNS千粒重TKW株高 PH0.41∗∗∗0.15∗∗0.47∗∗∗株高 PH0.57∗∗∗-0.22∗∗0.27∗∗∗穗长 SL0.41∗∗∗0.49∗∗∗0.3∗∗∗穗长 SL0.57∗∗∗-0.07ns0.26∗∗∗穗粒数 GNS0.15∗∗0.49∗∗∗0.035ns穗粒数 GNS-0.22∗∗-0.07ns-0.24∗∗∗千粒重TKW0.47∗∗∗0.3∗∗∗0.035ns千粒重TKW0.27∗∗∗0.26∗∗∗-0.24∗∗∗
注:*,**和***分别表示在P=0.05、P=0.01和P=0.001水平上显著相关。
Note:*, ** and *** indicate significance at the 0.05,0.01 and 0.001 levels, respectively.
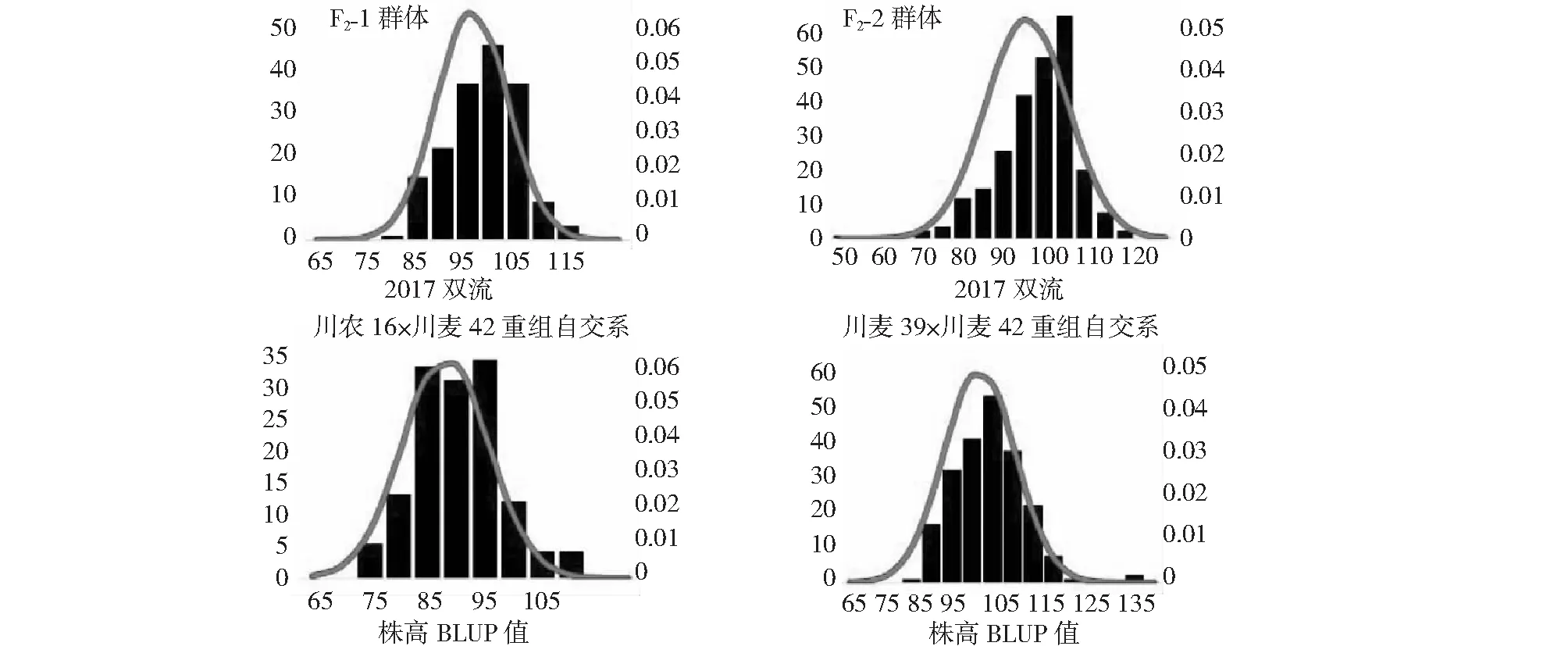
图1 株高在4个群体中的频率分布Fig.1 Frequency distribution of plant height (PH) in four populations
2.2 利用F2群体验证Qph.cib-5A和Qph.cibb-7A的真实性
2.2.1 KASP标记开发及F2群体筛选 将陆炳等鉴定到的与Qph.cib-5A和Qph.cibb-7A紧密连锁的SNP标记,开发成了多个可直接用于PCR检测的KASP标记。通过检测川麦42和川农16之间的差异,最终在5A和7A上各得到1条在两亲本之间具有显著差异的KASP标记,分别命名为“PH-5A”和“PHb-7A”。其中PH-5A位于染色体5A长臂461.49 Mb,PHb-7A位于染色体7A长臂562.10 Mb。由于PH-5A和PHb-7A是根据与Qph.cib-5A和Qph.cibb-7A紧密连锁的SNP设计而来,因此这2个KASP标记也与Qph.cib-5A和Qph.cibb-7A存在紧密连锁关系,可在其他群体中对Qph.cib-5A和Qph.cibb-7A进行追踪定位和分型(表3,图2)。
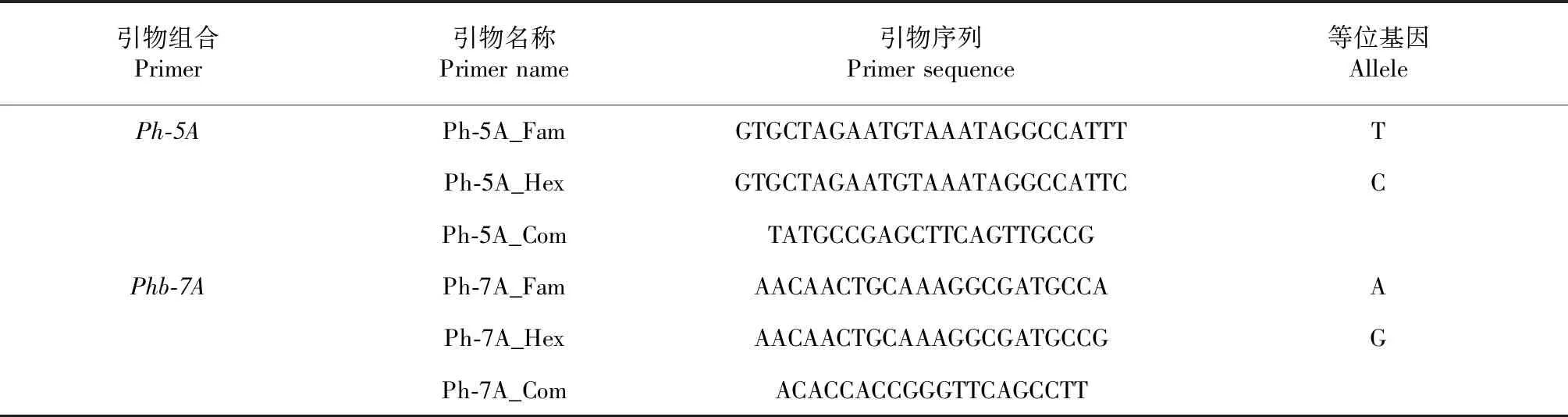
表3 Qph.cib-5A和Qph.cibb-7A位点开发的KASP标记引物
KASP检测结果显示,153个F2-1个体中在Qph.cib-5A位点纯合AA基因型(即具有正向增加株高的等位基因型)的株系有33株,纯合aa基因型的株系有51株,杂合Aa基因型的株系有69株;187份F2-2株系中,在Qph.cib-7Aa位点,纯合BB基因型的株系50株,纯合bb基因型的株系46株,杂合Bb基因型的株系91株(图2)。
2.2.2Qph.cib-5A和Qph.cibb-7A在F2群体中对株高的效应 根据基因型将F2-1和F2-2群体分成3组,比较3组不同基因型单株高度差异。在F2-1群体中,Qph.cib-5A位点纯合AA基因型株系的株高为100.36 cm;纯合aa基因型株系的株高为92.58 cm;杂合Aa基因型株系株高为93.8 cm。其中纯合AA基因型株系株高显著高于纯合aa基因型株系株高7.78 cm(7.75 %)和杂合Aa基因型株系株高6.56 cm(6.54 %);杂合Aa基因型和与纯合aa基因型株系之间株高没有显著性差异(图3)。

“A”和“B”代表增加株高的基因型;“a”和“b”代表降低株高基因型;t检验(P<0.05)检测差异显著性;“1”表示AA与Aa或BB与Bb之间差异显著性;“2”代表Aa与aa或Bb与bb之间的差异显著性;“3”代表AA与aa或BB与bb之间差异显著性;*,**和***分别表示在P = 0.05、P = 0.01和P=0.001水平上显著相关‘A’ and ‘B’ represent the genotype of increasing plant height; ‘a’ and ‘b’ represent the genotype of reducing plant height; Student’s t test (P<0.05) was used to identify differences; ‘1’ represents the difference of AA/Aa or BB/Bb; ‘2’ represents the difference of Aa/aa or Bb/bb; ‘3’ represents the difference of AA/aa or BB/bb; *, ** and *** indicate significance at the 0.05,0.01 and 0.001 level, respectively图3 Qph.cib-5A和Qph.cibb-7A在F2群体中对株高的遗传效应Fig.3 The effect of Qph.cib-5A and Qph.cibb-7A for plant height in F2 population
Table 4 The effect ofQph.cib-5AandQph.cibb-7Ain the Chuannong 16×Chuanmai42 and Chuanmai 39 × Chuanmai42 recombinant inbred lines

群体Populationtraits性状Qph.cib-5AAAaa差异(%)DifferenceQph.cibb-7ABBbb差异(%)DifferenceQph.cib-5A/Qph.cibb-7AAABBaabb差异(%)DifferenceCM42×CN16株高(PH)92.1±6.482.9±5.510∗∗∗90.2±7.484.7±6.76.1∗∗∗93.7±5.781.4±5.513.1∗∗∗穗长(SL)9.9±0.79±0.59.1∗∗∗9.6±0.79.5±0.7110±0.69.2±0.58∗∗∗穗粒数(GNS)46.8±4.547±3.9-0.446.8±4.147.7±4.3-1.946.2±4.346.8±3.7-1.3千粒重(TKW)50.9±3.248.6±3.44.5∗∗50.5±3.348.5±3.54∗∗52±348.1±3.97.5∗∗CM42×CM39株高(PH)98.7±7.794.6±74.2∗∗∗98.4±8.795.2±6.13.3∗102.3±8.794.7±67.4∗∗∗穗长(SL)12.5±1.612±1.24∗12.3±1.612.4±1.3-0.812.5±1.812.1±1.23.2穗粒数(GNS)46.4±3.347±5-1.347.5±4.246.7±4.71.746.8±3.847.4±5.6-1.3千粒重(TKW)51±3.849.7±4.42.5∗51.3±4.250±3.52.5∗51.5±3.549.1±3.94.7∗∗
注:“A”和“B”代表增加株高的基因型;“a”和“b”代表降低株高基因型;值表示为均值±标准偏差;差异性=(高值-低值)/高值× 100 %,t检验(P<0.05)检测差异显著性; *,**和***分别表示在P=0.05、P=0.01和P=0.001水平上显著相关。
Note:‘A’and ‘B’ represent the genotype of increasing plant height; ‘a’ and ‘b’ represent the genotype of reducing plant height; Values are the mean ± SD (standard deviation);Student’s t test (P< 0.05) was used to identify differences, Difference (%)=(Mean tall-Mean dwarf)/Mean tall × 100 %; *, ** and *** indicate significance at the 0.05,0.01 and 0.001 level, respectively.
F2-2群体中,在Qph.cibb-7A位点纯合BB基因型株系株高为99.47 cm,纯合bb基因型株系株高为93.31 cm,杂合Bb基因型株系株高为96.52 cm。显著性差异分析显示,纯合BB基因型株系的株高显著高于纯合bb基因型株系株高6.16 cm(6.19 %)和杂合Bb基因型株系的株高2.95 cm(2.97 %)。纯合bb基因型株系和杂合Bb基因型株系之间株高同样具有显著性差异且Bb基因型株系株高高于bb基因型株系3.21 cm(3.33 %,图3)。
2.3 Qph.cib-5A和Qph.cibb-7A在不同遗传背景中的效应及与主要产量性状的关系
分析川麦39、川麦42和川农16在Qph.cib-5A和Qph.cibb-7A位点的基因型显示,在这2个株高位点川麦39与川农16具有相同的基因型。因此用川麦42和川麦39构建的重组自交系群体在这2个位点中存在分离。基于此为了进一步鉴定Qph.cib-5A和Qph.cibb-7A在不同遗传背景中的效应,根据Qph.cib-5A(461.49 Mb)和Qph.cibb-7A(562.10 Mb)在染色体上的位置,结合90K SNP芯片和35K SNP芯片对川麦42×川农16重组自交系和川麦42×川麦39重组自交系的基因分型结果,将2个群体分为6组。通过比较6个组别之间株高、穗长、穗粒数和千粒重的差异,解析Qph.cib-5A和Qph.cibb-7A在不同遗传背景中对株高和产量性状的影响。
由表4可知,在川麦42×川农16重组自交系群体中Qph.cib-5A能显著的增加株高10 %,穗长9.1 %和千粒重4.5 %,对小麦的穗粒数没有影响。在川麦42×川麦39重组自交系中Qph.cib-5A显著增加株高4.2 %,穗长4 %和千粒重2.5 %。Qph.cibb-7A对株高和千粒重的显著影响在2个群体中也同时存在。在川麦42×川农16重组自交系群体中,Qph.cibb-7A株高和千粒重的增加效应分别为6.1 %和4 %;在川麦42×川麦39重组自交系中,Qph.cibb-7A对株高和千粒重的增加3.3 %和2.5 %,而对穗长和穗粒数基本没影响。此外,当同时聚合了Qph.cib-5A和Qph.cibb-7A时,在川麦42×川农16重组自交系群体中株高增加13.1 %,穗长增加8 %,千粒重增加7.5 %,对穗粒数基本无影响;而在在川麦42×川麦39重组自交系中,株高增加7.4 %,千粒重增加4.7 %,穗长和穗粒数基本不受影响。
3 讨 论
3.1 Qph.cib-5A和Qph.cibb-7A与已报道QTL位点比较
国内外育种家在不同的遗传群体,不同的种植环境,不同的遗传背景下定位到了许多株高相关QTL。Yu等利用W7984×Opata85构建了重组自交系,基于高密度遗传图谱在染色体5A的104.22 Mb处鉴定到一个株高相关QTL[23]。Gao等利用90K芯片,在周8425B×中国春重组自交系中鉴定到2个株高相关QTL,分别位于染色体5A短臂和7A短臂[11]。叶亚琼等利用2个冬小麦品种陇鉴19与Q9086杂交创建的重组自交系群体在染色体5A染色体Xbarc141~Xcfd2121区间和7A染色体Xpsp3001~Xgwm63区间各检测到一个多环境稳定表达的株高QTL位点[24],BLAST结果显示,Xbarc141位于5A长臂468.94 Mb,Xpsp3001位于7A染色体94.43 Mb,Xgwm63位于7A染色体133.13 Mb。此外在已经报道的Rht基因中,Rht12和Rht22分别位于染色体5A和7A,其中Rht12位于在染色体5A长臂678.29 Mb[25]。Rht22位于染色体7A短臂[26]。本实验室陆炳等利用2个四川麦品种川农16和川麦42创制的RIL群体[20],基于90K芯片鉴定到的两个主效株高QTL,Qph.cib-5A和Qph.cib-7A,分别与本研究开发的位于染色体5A长臂461.49 Mb和染色体7A长臂562.10 Mb处的KASP标记紧密连锁。与先前报道的株高相关QTL和Rht基因比较发现,Qph.cib-5A与叶亚琼等报道的一个株高QTL和Rht12都位于染色体长臂,因此需要进一步验证是否为同一个株高位点;Qph.cibb-7A与先前报道的QTL和Rht基因都不同,可能是一个新的主效株高QTL。
3.2 Qph.cib-5A和Qph.cibb-7A与Rht基因对株高效应的比较
目前,已命名的株高相关Rht基因有24个,分布于小麦染色体2AS、2BL、2DS、3BS、4BS、4DS、5AL、5DL、6A和7A上[27-31]。其中只有Rht-B1b、Rht-B1c、Rht-D1b和Rht-D1c被克隆。在前人对Rht基因的研究中,许琦等在210份冬小麦品系中发现Rht-B1b、Rht-D1b和Rht8对小麦平均株高的降秆作用分别为9.7、16.7和2.5 cm[32]。赤霉素敏感基因Rht4、Rht8、Rht12和Rht13可以通过缩短小麦不同节间长度分别降低株高17 %、7 %、40 %和34 %[14]。Rebetzke等报道Rht5能够显著的降低株高55 %[13];Wurschum等报道Rht24能够降低小麦株高8 cm[15]。在本研究中,Qph.cib-5A和Qph.cibb-7A在不同群体对株高都具有显著效应。F2群体中单个Qph.cib-5A和Qph.cibb-7A分别增加株高7.78和6.16 cm,对株高的遗传效应与Rht8相似。相同的效应在其他2个重组自交系群体中也能被观察到,只是在川麦42×川麦39重组自交系群体中对株高的影响相对较弱,这可能是由于在该群体中还有其他主效基因控制株高,使得Qph.cib-5A和Qph.cibb-7A的遗传效应不能充分体现导致的。此外在Qph.cib-5A位点,杂合基因型株系的株高显著低于纯合AA基因型株系的株高,而与纯合aa基因型株系株高没有差异,说明在Qph.cib-5A位点降低株高的aa基因型具有较强的显性效应。
3.3 Qph.cib-5A和Qph.cibb-7A在不同遗传背景中的效应及其与主要产量性状的关系
过矮的植株,标志着营养器官的消弱,不利于产量的增加[16]。而过高的植株又会导致植株抗倒伏能力减弱,种植密度降低,最终也会使小麦减产。因此在育种过程中对于株高的调控,一定要协调其与主要产量性状之间的关系,只有这样才能达到最大收获指数的育种目标。基于此,本研究通过比较Qph.cib-5A和Qph.cibb-7A在2个重组自交系群体中对株高、千粒重、穗粒数和穗长的影响,解析其在调控株高的同时与主要产量性状之间的关系。研究发现,Qph.cib-5A和Qph.cibb-7A在增加株高4.1~9.2和3.2~5.5 cm的同时能显著增加千粒重1.3~2.3和1.3~2 g,而对穗粒数无明显影响。此外,当聚合了Qph.cib-5A和Qph.cibb-7A时,植株的株高和千粒重分别增加7.6~12.3 cm和2.4~3.9 g,表现出效应的叠加。虽然Qph.cib-5A和Qph.cibb-7A对株高的影响不及多数Rht基因,但这种叠加效应能够在较大范围(3.2~12.3 cm)内对株高进行调控。下一步将继续开发与Qph.cib-5A和Qph.cibb-7A连锁更加紧密的标记,以便于其在育种中更好的利用。
4 结 论
Qph.cib-5A和Qph.cibb-7A在不同的遗传背景中对小麦株高都具有显著的调控作用,此外在调控株高的同时会影响千粒重和穗长,而不会作用于穗粒数。同时聚合2个位点时对株高、穗长、千粒重的效应值大于只有单个QTL或没有QTL位点时的效应。该研究可为株高遗传机制的解析和控制株高基因的精细定位提供理论依据,也能为小麦育种提供可用的分子标记和理论基础。
