浙麦冬根际及非根际细菌菌群结构与多样性研究
2019-03-26石森林指导
陈 宸 石森林(指导)
浙江中医药大学 浙江 杭州 310053
本研究以慈溪和桐庐两个产地浙麦冬为研究对象,通过高通量测序手段研究浙麦冬根际与非根际土壤微生物的结构与多样性差异,为浙麦冬根际微生态的研究提供基础数据,同时为进一步研究浙麦冬对根际微生态的影响机制奠定基础。
1 材料与方法
1.1 样本采集:于2018年9月在慈溪(CX)和桐庐(TL)浙麦冬产区进行样品采集,在二年生浙麦冬种植区域均匀选取3个采样点,每个采样点通过五点取样法采集样本。将浙麦冬植株连根拔起,抖落根系外围的大块土壤,利用1L磷酸缓冲液清洗得到粘附在根部的土壤作为根际土壤(G),在植株附近无植物生长的地方取20cm深处土壤作为非根际土壤(FG),将从两个产区采集到的各两种土壤样本分别标记为:慈溪根际土壤(CXG)、慈溪非根际土壤(CXFG)、桐庐根际土壤(TLG)和桐庐非根际土壤(TLFG)。将同类样本混合均匀,使用qigen DNAeasy powerSoil kit试剂盒(qigen公司,美国)按说明书提取获得DNA,使用Nanodrop2000紫外分光光度计检测浓度,通过琼脂糖凝胶电泳分析DNA完整性。
1.2 16SrDNA基因扩增及Ion Torrent S5测序:采用引物341F/806R(341F:CCTAYGGGRBGCASCAG;806R:GGACTACNNGGGTATCTAAT)扩增16SrDNA V3-V4可变区片段。PCR扩增程序如下:95℃for 3 minutes;35cycles of:-95℃ for 30 seconds;-58℃ for 30 seconds;-72℃ for 30 seconds;-72℃for 5 minutes;获得的PCR产物进行磁珠筛选纯化,得到PCR产物。将每个样品等比例混合后,依托Thermo Fisher Scientific公司Ion Torrent S5平台进行测序(由浙江天科高新技术发展有限有限公司提供)。
1.3 数据分析:测序得到的raw data,通过生物信息分析去除接头和barcode序列;然后使用QIIME软件对数据进行过滤,过滤掉含N较多或者低质量序列,最后进行嵌合体过滤,得到可用于后续分析的有效数据(序列平均长度在420bp左右),利用Uparse软件对所有样品的全部Effective Tags进行聚类,默认以97%的一致性将序列聚类成为OTUs,同时会选取OTUs的代表性序列,依据其算法原则,筛选的是OTUs中出现频数最高的序列作为OTUs的代表序列。对OTUs代表序列进行物种注释,用uclust方法与Silva数据库进行物种注释分析,并分别在各个分类水平:kingdom(界),phylum(门),class(纲),order(目),family(科),genus(属),species(种)统计各样本的群落组成。使用PyNAST软件与Silva数据库中的“Core Set”数据信息进行快速多序列比对,得到所有OTUs代表序列的系统发生关系。最后对各样品的数据进行均一化处理,以样品中数据量最少的为标准进行均一化处理,后续的Alpha多样性分析和Beta多样性分析都是基于均一化处理后的数据。使用QIIME 软件计算 Observed-species,Chao1,Shannon,Simpson,ACE,Goods-coverage指数,使用R软件进行Alpha多样性指数组间差异分析;Alpha多样性指数组间差异分析选用T-test检验。
2 结果
2.1 根际与非根际土壤细菌多样性指数比较:将4种土壤样本的Effective Tags进行聚类,以97%的一致性将序列聚类成为OTUs。对OTU数量进行比较,未发现显著差异。通过Shannon指数、Simpson指数、ACE指数和Chao1指数比较不同土壤微生物Alpha多样性,无显著差异。详见表1。
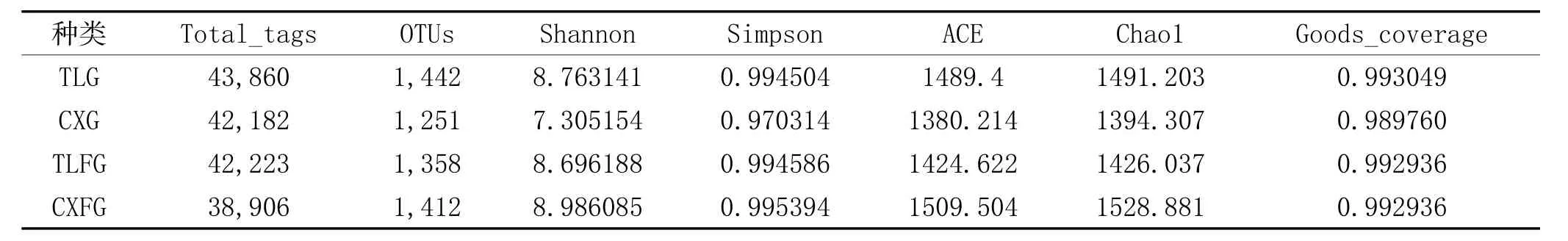
表1 四种土壤样本细菌多样性指数
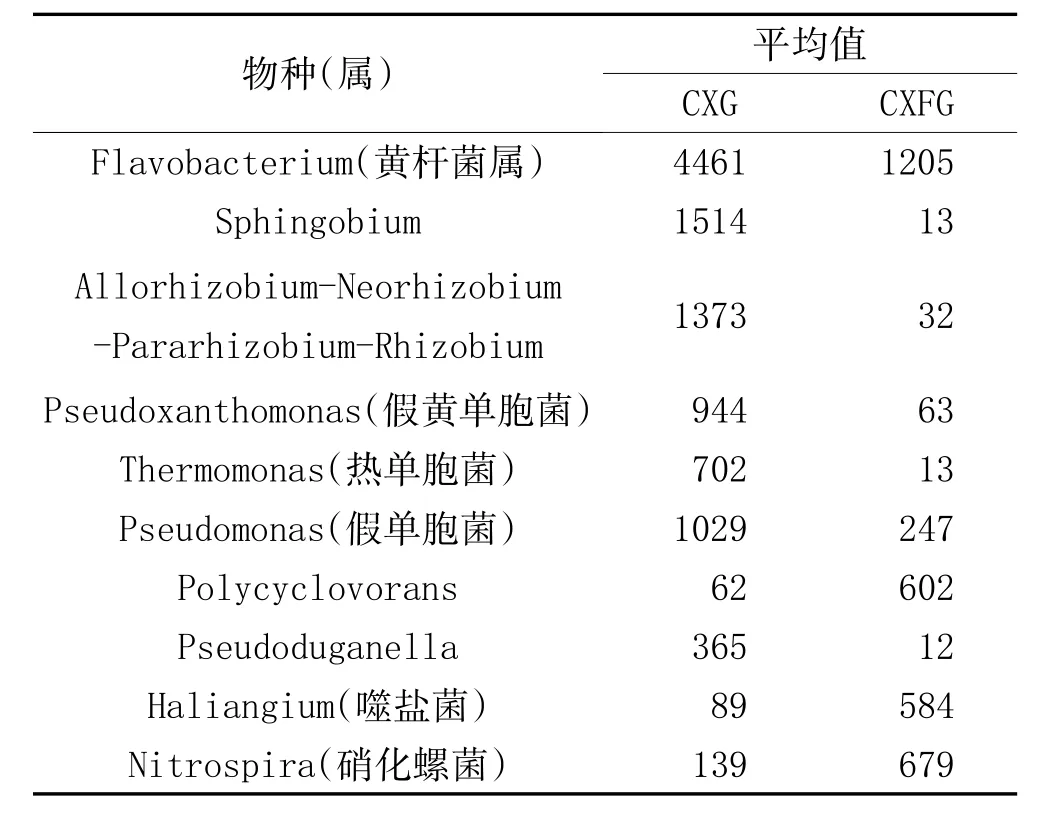
表2 CXG与CXFG的Fisher样本间物种差异分析

表3 TLG与TLFG的Fisher样本间物种差异分析
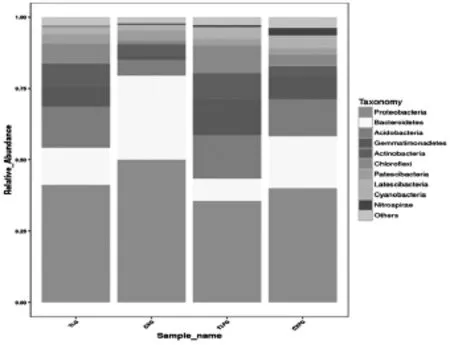
图1 门水平上的物种相对丰度柱形图
2.2 细菌群落组成及多样性分析:CXG、CXFG、TLG、TLFG四种土壤分别有27、27、24和27个门类菌群。根据物种注释结果,四种土壤中丰度最大的十个门类相同,分别是:Proteobacteria(变形菌门)、Bacteroidetes(拟杆菌门)、Acidobacteria(酸杆菌门)、Gemmatimonadetes(芽单胞菌门)、Actinobacteria(放线菌门)、Chloroflexi(绿弯菌门)、Patescibacteria、Latescibacteria、Cyanobacteria(蓝藻门)和Nitrospirae(硝化螺旋菌门)。从图1可以看出,Proteobacteria(变形菌门)、Bacteroidetes(拟杆菌门)、Patescibacteria和Cyanobacteria(蓝藻门)在根际土壤中的相对丰度明显高于在非根际土壤中,而Acidobacteria(酸杆菌门)和Actinobacteria(放线菌门)的相对丰度在两者间基本持平,Gemmatimonadetes(芽单胞菌门)、Chloroflexi(绿弯菌门)、LatescibacteriaNitrospirae(硝化螺旋菌门)在非根际土壤中的相对丰度则明显高于在根际土壤中。
2.3 细菌群落结构差异分析:将4种土壤样本的OTU分布进行Venn图分析,4种土壤中均有分布的OTU数量为600,TLG与TLFG共有OTU数量为1093,CXG与CXFG共有OTU数量为967,TLG与CXG共有OTU数量为827,TLFG与CXFG共有OUT数量为903,4种土壤依独有OUT数量由高到低次序为:CXFG(155)、TLG(117),TLFG(93),CXG(68)。在属分类水平上对四种土壤样本进行CXG与CXFG、TLG与TLFG样本间Fisher检验,找出差异显著的物种。在CXG与CXFG可鉴定到属的266个属中有显著差异的(P<0.05)有147个,在TLG与TLFG可鉴定到属的254个属中有显著差异的(P<0.05)有112个。取差异最显著的前10个属展示,见表2、表3。
3 讨论
麦冬作为重要的常用中药材,最早在《神农本草经》中就有记载,曰其“气味甘、平,无毒”“久服轻身不老,不饥”。有临床报道,沙参麦冬汤能够延长非小细胞肺癌患者无进展生存期,并能显著改善患者生活质量[1]。而麦冬的药效往往与其产地有关。植物通过根系向土壤分泌有机物质影响土壤微生物,受到这种影响的土壤区域称为根际。以往研究表明,在麦冬生长期间,麦冬根系向根外不断分泌有机物质,这些有机物质对根际土壤微生物的物种结构会产生影响[2]。而根际土壤微生物的结构变化反过来会对土壤肥力和植物生长产生重要影响[3]。本研究发现,根际土壤细菌与非根际土壤细菌在门和属水平上结构存在明显差异,特别是属水平上物种差异明显。麦冬根系对黄杆菌属、假单胞杆菌属、不动细菌属、亚杆菌属和肠杆菌属这些反硝化细菌有明显选择性,与水稻情况基本相似[4]。值得注意的是,慈溪与桐庐两地非根际土壤可能因受自然条件、土壤理化性质的影响,细菌物种数量和群落结构有明显差异,但两地根际土壤根际土壤细菌物种数量和群落结构明显趋近。这表明浙麦冬根系分泌物对根际土壤细菌的种类和群落结构有明显调控作用。
