海洋酸度变化下孔石莼对农药硫丹的生态响应研究❋
2019-02-21李晓东谭海丽
李晓东, 谭海丽, 尤 宏,2❋❋
(1.哈尔滨工业大学(威海)海洋科学与技术学院,山东 威海 264200;2.哈尔滨工业大学市政环境工程学院环境科学与工程系,城市水资源与水环境国家重点实验室,黑龙江 哈尔滨 150090)
海洋酸化是指由于海洋吸收、释放大气中过量的二氧化碳,导致海水逐渐变酸的过程。联合国政府间气候变化委员会(IPCC)第五次评估报告指出:自工业革命以来,海水pH 值已下降了0.1 个单位,相当于氢离子质量分数增加了26%[1],海洋酸化成为继“温室效应”之后又一由CO2过量排放而引起的全球性环境问题。
随着农业生产的迅速发展,环境中农药残留问题日渐引起广泛关注。其中硫丹是一种广谱有机氯杀虫剂,1954年由德国Hoechst 公司研制开发,因具备良好的胃毒和触杀作用,曾被广泛的运用在农业谷物、蔬菜、水果、棉花、林木等病虫害防治[2],2011年4月29日硫丹被列入斯德哥尔摩公约禁用物质列表,目前世界上已有60多个国家禁止使用,但由于过去几十年硫丹的大量使用加上其潜在的迁移能力,全球范围内各种环境介质中,如大气、土壤、沉积物、水体均已普遍检测出硫丹,中国从2003年在部分水体中就检出硫丹[3-6]。
同时硫丹对水生生物有极高的毒性,并且随着海洋酸化的加剧,海水pH值的降低会导致海水中硫丹溶解度的增加,降解速度的减缓,对海洋生物的生态毒性效应将会进一步增强。2017年2月2日,我国环境保护部环境保护对外合作中心与联合国开发计划署(UNDP)共同开发的 “中国硫丹淘汰项目”获得全球环境基金(GEF)批准。表明硫丹对生物的毒害性已引起国际社会和政府的广泛关注。
目前国内外对硫丹的研究,主要集中在对动物的毒性效应,如在高等哺乳动物和部分鱼类[7-9],而对于水生浮游植物的毒性研究较少;硫丹有着较强的生态毒性,并且随着海洋酸化现象日趋严重,硫丹的理化性质也受到了影响,如硫丹的溶解度随海水 pH 值的降低而增大,并且在低 pH 环境下硫丹在水中的半衰期增大,降解速率变缓。大型海藻是海洋中初级生产力的重要组成部分,在不到海洋总面积1%的沿岸带构成海洋总初级生产力的10%,在近岸碳循环方面起着至关重要的作用,还可为海洋中的无脊椎动物及鱼类提供产卵孵化和栖息的场所,而人类活动导致的环境变化问题正逐渐加剧,因此研究硫丹和海水酸度变化耦合作用对大型海藻的影响有着深远的意义。本文以绿藻门的孔石莼(UlvapertusaKjellman)为研究对象,考察了硫丹单独胁迫及其与酸度联合胁迫下对孔石莼在个体及生理生化方面的影响,为今后减轻硫丹对大型海藻及海洋生态系统的危害提供科学依据。
1 材料与方法
1.1 实验材料
本实验中的大型海藻孔石莼均采自威海市三连岛潮间带海域(海水pH=8.1 ,温度16 ℃,盐度31)。采集时选择健康一致的藻体,放在盛有少量海水的塑料袋中于2 h内运至实验室,选择生长良好的藻体,洗净附着的泥沙和浮游生物,其中将孔石莼打成直径1.5 cm的圆片,在实验室温度(20 ℃)及光照条件(光照周期为L∶D = 12 h∶12 h)下放入5 L玻璃缸中驯养采用灭菌后的陈海水(pH=8.2盐度31),每天24 h曝气。驯养4 d后进行正式实验。
1.2 试剂与仪器
硫丹 EC 乳油,有效含量为 35%,购自于河北省冀州市凯明农药有限责任公司。实验前用蒸馏水配成有效含量为 10 mg·L-1的母液,实验时按所需质量浓度进行稀释。其余试剂均为分析纯,国药集团化学试剂有限公司生产。
电子天平(台衡MP200A,中国)、实验室pH计(METTLER TOLEDOFE20,瑞士)、立式自动压力蒸汽灭菌器(致微GI54T,中国)、洁净工作台(安泰SW-CJ,中国)、紫外-可见分光光度计(尤尼柯TU-1800S,中国)、台式高速冷冻离心机(利康Neofuge BR,中国香港)。
1.3 实验设计
根据海水实际酸度,设定2个pH梯度,分别为7.9和8.2;根据硫丹预实验,设置浓度梯度为0、50、150、250、350 μg/L的硫丹实验组及pH联合胁迫实验设置如表1所示,每个处理组中设置3个平行样,在超净工作台内每组接种孔石莼1.3 g左右,实验周期为7 d,每天用0.01 mol/L NaOH和0.01 mol/L HCl溶液调节各处理组培养液的pH。
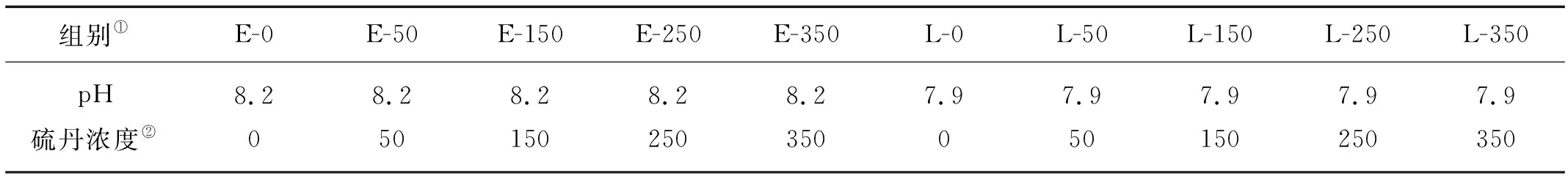
表1 硫丹及pH联合胁迫实验各处理组的设计Table 1 Design of endosulfan and pH stress experiment treatment groups
Note:①Groups;②Endosulfan concentration
1.4 孔石莼生长情况的测定
各实验组分别于暴露第1天、第3天及第7天测定孔石莼藻体鲜重的变化,孔石莼叶片称量前用吸水纸吸干。根据公式(1)计算孔石莼的相对生长率:
相对生长率(%) = [ ln(Wt/W0) /t]×100%。
(1)
其中:W0为初始湿重;Wt为td后的湿重。
1.5 叶绿素含量的测定
各实验组,分别于暴露第1、3、7天取出藻体0.2 g,放入预冷的研钵,加入少量石英砂和碳酸镁粉后在冰浴上研磨,之后加入10 mL 95%乙醇,在4 ℃冰箱中避光提取24 h,提取完毕在4 000 r/min 下离心 20 min,测定上清液在665、649和470 nm波长下的吸光度,并计算光合色素的含量。叶绿素a、叶绿素b和类胡萝卜素浓度计算公式如下:
Ca=13.95A665-6.88A649;
(2)
Cb=24.96×A649-7.32×A665;
(3)
Cx·c=4.08A470+3.31A665-11.64A649。
(4)
其中:Ca为叶绿素a的浓度;Cb为叶绿素b的浓度;Cx·c为类胡萝卜素的浓度;A665、A649和A470为665、649和470 nm的吸光度值。
1.6 抗氧化酶活力的测定
每个处理组分别于暴露第1天、第3天及第7天另取孔石莼叶片0.2 g,于预冷的研钵中加1 mL预冷的磷酸缓冲液(pH=7.8)在冰浴上研磨成浆,加磷酸缓冲液使体积为5 mL,于4 000 r/min下离心20 min后,取上清液测定蛋白质含量及酶活力。
SOD活性的测定采用Bewley等[10]改进的氮蓝四唑(NBT)光化学反应法,POD活性的测定采用 Srivestava[11]等改进的愈创木酚法。可溶性蛋白质含量测定采用考马斯亮蓝G-250染色法进行[12]。
1.7 数据处理
实验结果以平均值±标准差(Mean±SD) 表示,采用 SPSS 19.0软件对硫丹单独胁迫后的实验结果进行单因素方差分析,对硫丹胁迫后与pH耦合的实验结果进行多因素方差分析,分析硫丹、pH及硫丹于pH耦合后对孔石莼个体及生理生化方面的影响。设置显著水平为P<0.05,极其显著水平为P<0.01。
2 结果与分析
2.1 硫丹胁迫及与pH耦合对孔石莼生长的影响
2.1.1 硫丹对孔石莼生长的影响 图1为硫丹对孔石莼相对生长率的影响。由图可知,随着硫丹浓度的逐渐升高,孔石莼的相对生长率在第1天和第3天时均出现了显著的先升高后下降的趋势。对照组在第3天相对生长率升高了22.73%,而在第7天变化则不明显。与对照组相比,各浓度处理组的相对生长率在第1天分别变化了98.45%、61.56%、-49.85%、-124.15%,在第3天分别变化了86.25%、27.99%、-92.99%、-169.59%,在第7天则分别变化了13.96%、-14.37%、-132.92%、-195%。由此可以看出,50 μg/L的处理组在第1天和第3天相对生长率得到明显促进,而在第7天基本上没有促进作用;250和350 μg/L处理组孔石莼的相对生长率均出现明显的抑制作用(P<0.01),并且抑制作用随着暴露时间的延长而逐渐加剧。250 μg/L处理组孔石莼相对生长率从第1天至第3天下降了82.84%,第3天至第7天下降了551.33%,在第7天时出现了负增长现象,藻体叶片变薄并且表现出白化现象,而高浓度350 μg/L处理组的孔石莼在胁迫第1天就已出现了负增长,至第七天孔石莼叶片已表现出颜色变白、腐烂和非常易碎等症状。
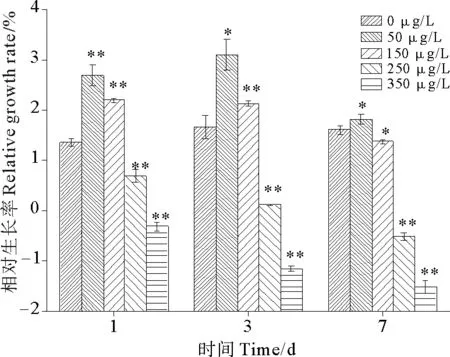
图1 硫丹对孔石莼相对生长率的影响Fig.1 Effects of treatments with different endosulfan concentrations on relative growth rate (RGR) of Ulva pertusa
2.1.2 硫丹与pH耦合对孔石莼生长的影响 图2为pH与不同浓度硫丹耦合对孔石莼相对生长率的影响,所有低pH的处理组与对照组相比都具有极显著性(P<0.01)的差异。在环境pH和低pH条件下孔石莼受硫丹胁迫相对生长率都表现为低浓度促进高浓度抑制效应,并且均在50 μg/L处理组时相对生长率值达到最高峰。硫丹胁迫第1天,与对照组相比,环境pH和低pH的各浓度处理组分别变化了98.45%、61.56%、-49.85%、-124.15%和84.70%、222.43%、117.42%、-51.63%、-123.94% 。说明在硫丹暴露第1天时,孔石莼的相对生长率在低pH条件下受到的促进作用更明显,并且在L-0 μg/L处理组的相对生长率与对照组相比也表现为上升趋势。在暴露第3天时,各处理组分别变化了86.25%、27.99%、-92.99%、-169.59%和81.60%、262.55%、106.95%、-92.55%、-177.07%。说明在低pH条件下暴露第3天时,孔石莼的相对生长率受到硫丹的胁迫作用更显著,低浓度的刺激作用和高浓度的抑制作用更加明显。至暴露第7天,各处理组分别变化了13.96%、-14.37%、-132.92%、-195%和69.61%、187.64%、43.42%、-140.37%、-201.95% 。随着时间的延长,暴露第七天所有处理组孔石莼的相对生长率相对于第1天和第3天均有所下降。
pH与不同浓度硫丹耦合对孔石莼生长主体间效应的检验后发现,暴露第1天、第3天及第7天硫丹、pH以及两者的交互作用对孔石莼的生长都发挥了作用,并且具有极其显著的影响(P<0.01)。
2.2 硫丹胁迫及与pH耦合对孔石莼光合色素的影响
2.2.1 硫丹对孔石莼光合色素的影响 图3为硫丹对孔石莼叶绿素a含量的影响。在硫丹暴露第1天、第3天和第7天,低浓度50 μg/L处理组的叶绿素a含量都表现出一致的促进作用,随着硫丹浓度的增加,其他各浓度处理组呈现不同程度的抑制作用;随着硫丹胁迫时间的延长,各浓度处理组的下降幅度不同,第1天至第3天,50和150 μg/L处理组叶绿素a的含量与对照组一样无明显变化趋势。250和350 μg/L处理组在整个暴露期间表现出持续的下降趋势,与对照组相比均表现出极显著性差异(P<0.01),说明高浓度硫丹对孔石莼叶绿素a有较大的损伤。在第3天,50 μg/L浓度处理组出现叶绿素a含量的最高峰值,至第7天后则出现了明显下降,但与对照组相比,没有显著性差异(P>0.05)。

图2 pH与不同浓度硫丹耦合对孔石莼相对生长率(RGR)的影响Fig.2 Effects of treatments coupling with different endosulfan concentrations and pH on relative growth rate (RGR) of Ulva pertusa
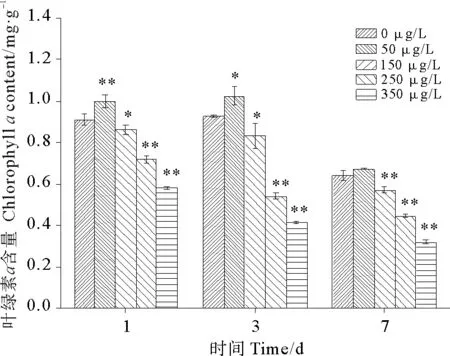
图3 硫丹对孔石莼叶绿素a含量的影响Fig. 3 Effects of treatments with different endosulfan concentrations on Chlorophyll a content of Ulva pertusa
图4为硫丹对孔石莼叶绿素b含量的影响。与第1天相比,各浓度组第3天叶绿素b的含量分别变化了2.8%、2.26%、1.15%、-4.73%、-15.68%,至第7天叶绿素b的含量分别变化了-13.29%、-13.95%、-6.88%、-17.66%、-30.46%。随着硫丹浓度的增加和暴露时间的延长,叶绿素b受到的抑制作用越来越明显,具有明显的剂量-时间效应。50 μg/L处理组的孔石莼在3个阶段均表现出明显的促进作用,表明一定浓度的硫丹农药会促进孔石莼叶绿素b的合成和积累,之后叶绿素b含量开始回落,随着硫丹浓度的增加而逐渐降低。
类胡萝卜素不仅是藻类的主要捕光色素,也是生物体中的一种抗氧化剂,对细胞具有保护作用,使生物细胞免受外界不良因子的伤害。由图5可知,在硫丹胁迫后的第1天,与对照组相比,仅350 μg/L处理组的类胡萝卜素含量表现出极显著性差异(P<0.01),其余处理组没有显著性差异(P>0.05)。然而随着硫丹胁迫时间加长,各浓度处理组逐渐表现出明显的差异性,至暴露第7天时,各硫丹处理组均出现极显著性差异(P<0.01)。说明硫丹胁迫下,孔石莼中类胡萝卜素的响应具有明显的时间依赖效应。
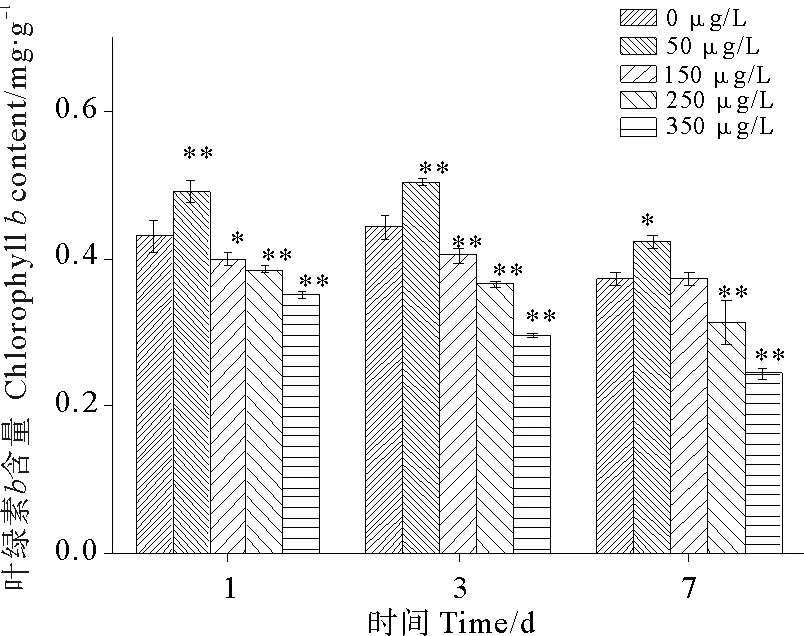
图4 硫丹对孔石莼叶绿素b含量的影响Fig.4 Effects of treatments with different endosulfan concentrations on Chlorophyll b content of Ulva pertusa

图5 硫丹对孔石莼类胡萝卜素含量的影响Fig.5 Effects of treatments with different endosulfan concentrations on Carotenoids content of Ulva pertusa
2.2.2 硫丹与pH 耦合对孔石莼光合色素的影响 pH与不同浓度硫丹耦合对孔石莼叶绿素a含量的影响如图6所示,环境pH与低pH条件下,各处理组孔石莼叶绿素a含量都表现为先升高后下降的趋势,并且低pH的各处理组与环境pH相比,孔石莼叶绿素a的含量相对下降。环境pH与低pH处理组均在暴露第3天50 μg/L处理组出现叶绿素a含量的峰值,并且与对照组相比,环境pH下的叶绿素a的含量升高了12.54%,然而低pH条件下的叶绿素a的含量是相对下降的。在低pH硫丹处理组,除了第7天的L-50 μg/L处理组之外,其余各处理组与对照组相比都显现出显著的差异性(P<0.05)。L-0 μg/L处理组与对照组相比,叶绿素a的含量在第1天、第3天和第7天分别下降了21.66%、20.92%、52.69% 。表明在pH为7.9时孔石莼叶绿素a的含量是下降的。
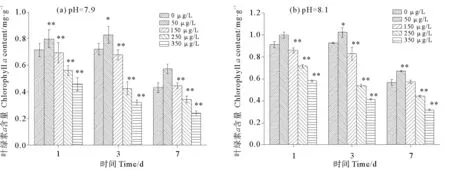
图6 pH与不同浓度硫丹耦合对孔石莼叶绿素a的影响Fig.6 Effects of treatments coupling with different endosulfan concentrations and pH on Chlorophyll a contentof Ulvapertusa
pH与不同浓度硫丹耦合对孔石莼叶绿素a主体间效应的检验后发现,暴露第1天、第3天和第7天,硫丹、pH对孔石莼叶绿素a含量均具有显著性影响(P<0.05),而硫丹与pH的交互作用对孔石莼叶绿素a的影响较小,仅在胁迫后的第3天发挥显著性效应(P<0.05),第1天和第7天均无显著性差异(P>0.05)。
图7为pH与不同浓度的硫丹耦合对孔石莼叶绿素b含量的影响。环境pH与低pH条件下,各处理组孔石莼叶绿素b含量都表现为先升高后下降的趋势,均在50 μg/L硫丹浓度时上升,环境pH的50 μg/L浓度是在第3天出现叶绿素b含量的峰值,低pH的50 μg/L浓度是在第1天出现叶绿素b含量的峰值,并且低pH的各处理组与环境pH相比,孔石莼叶绿素b的含量相对下降。在低pH硫丹处理组,除暴露第1天L-0和L-50 μg/L处理组之外,其他各处理组与对照组相比都有显著的差异性(P<0.05)。L-0 μg/L处理组与对照组相比,叶绿素b的含量在第1天、第3天和第7天分别下降了24.98%、21.20%、30.29% ,说明在pH为7.9的条件下孔石莼叶绿素b的含量是下降的。
pH与不同浓度硫丹耦合对孔石莼叶绿素b主体间效应的检验后发现,暴露第1天、第3天和第7天,硫丹、pH对孔石莼叶绿素b含量都具有显著性影响(P<0.05)。硫丹与pH的交互作用仅在胁迫后的第3天发挥显著性效应(P<0.05),第1天和第7天均无显著性差异(P>0.05)。

图7 pH与不同浓度硫丹耦合对孔石莼叶绿素b的影响Fig.7 Effects of treatments coupling with different endosulfan concentrations and pH on Chlorophyll b contentof Ulvapertusa
由图8可知,环境pH以及低pH条件下,各处理组孔石莼类胡萝卜素含量都表现为先升高后下降的趋势,均在50 μg/L处理组上升,并且在第3天达到最大值。低pH的各处理组与环境pH相比,孔石莼类胡萝卜素的含量相对下降。在低pH条件下,除了L-50 μg/L和暴露第1天的L-150 μg/L处理组之外,其他各处理组与对照组相比呈现出显著性差异(P<0.05)。L-0 μg/L处理组与对照组相比,类胡萝卜素的含量在第1天、第3天和第7天分别下降了19.45%、11.62%、38.54% 。说明孔石莼体内类胡萝卜素的含量在pH为7.9时总体呈下降趋势,但相对于第1天和第7天,第3天时类胡萝卜素的含量有所上升。

图8 pH与不同浓度硫丹耦合对孔石莼类胡萝卜素的影响Fig.8 Effects of treatments coupling with different endosulfan concentrations and pH on Carotenoids content of Ulvapertusa
pH与不同浓度硫丹耦合对孔石莼类胡萝卜素主体间效应的检验后发现,硫丹、pH对孔石莼类胡萝卜素含量都具有显著性影响(P<0.05),并且在第1天、第3天及第7天均发挥了显著作用(P<0.05)。反而硫丹与pH的交互作用在第1天、第3天和第7天均没有发挥显著性效应(P>0.05)。
2.3 硫丹胁迫及与pH耦合对孔石莼抗氧化酶活性的影响
2.3.1 硫丹对孔石莼抗氧化酶活性的影响 硫丹会诱导藻体细胞产生大量活性氧物质,导致细胞内活性氧的生成和清除平衡被破坏,从而造成氧化损伤。本实验中主要考察了硫丹对孔石莼体内两种抗氧化酶,超氧化物歧化酶(SOD)和过氧化物酶(POD)的影响。硫丹对孔石莼SOD总活性的影响如图9所示。硫丹暴露第1天,50 μg/L处理组的SOD总活性最高,与对照组相比变化了129.36%,之后随着硫丹浓度的增加,各浓度处理组SOD总活性逐渐降低并且下降幅度基本上一致。暴露第3天,SOD总活性不再存在促进作用,但在150 μg/L出现一个转折点,之后的各浓度组SOD总活性的下降幅度增大。至第7天,各处理组一致表现出抑制作用,与对照组相比,差异性极显著(P<0.01),基本上呈现一种线性下降趋势,且下降幅度达到最大。除了对照组SOD总活性上升之外,各处理组SOD总活性随着暴露时间的延长均表现为抑制作用;至第7天,与对照组相比,各浓度处理组SOD总活性分别下降了13.80%、24.75%、35.96%、47.82%。
图10为硫丹对SOD比活性的影响,由图可知,孔石莼SOD比活性的变化趋势于总活性完全不同。各浓度组SOD比活性在实验周期内均受到不同程度的促进作用,并且在第7天时250 μg/L处理组受到的促进作用最明显,达到一个极显著的高峰值,与对照组相比升高了47.29%(P<0.01)。除了第3天各浓度组SOD比活性变化不明显之外,第1天和第7天各处理组与对照组相比,均表现出极显著的差异性(P<0.01)。第1天和第3天,随着硫丹浓度的增加,SOD比活性有些许下降,变化幅度不明显;至第七天,随着浓度的升高SOD比活性出现先升高后下降的趋势,与对照组相比,各浓度组分别上升了5.52%、13.38%、27.57%、17.88%,在350 μg/l处理组时开始呈现下降趋势,但仍然高于其他浓度组的比活性。
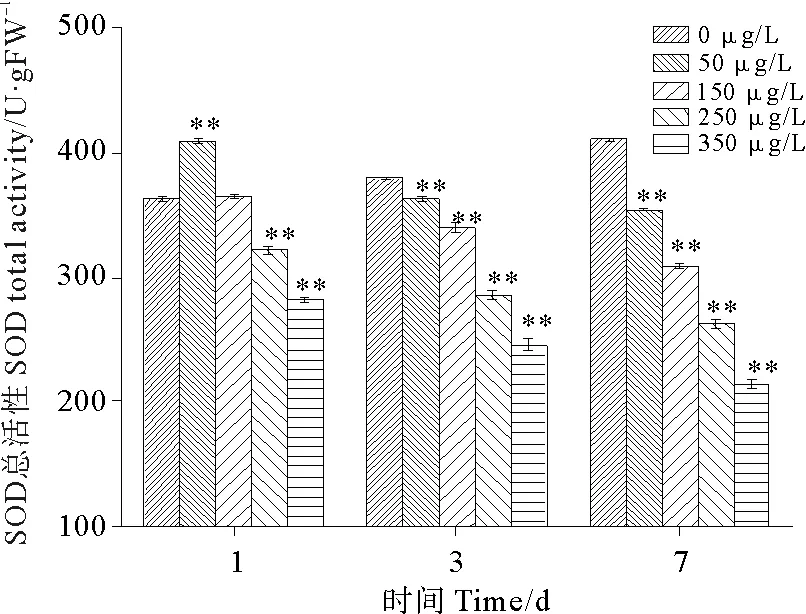
图9 硫丹对孔石莼SOD总活性的影响Fig.9 Effects of treatments with different endosulfan concentrations on SOD total activityof Ulvapertusa
硫丹对孔石莼POD总活性的影响如图11所示,暴露第1天和第3天,孔石莼POD总活性均是出现先升高后降低的趋势,而在第7天为持续下降趋势,且各处理组之间具有显著性差异(P<0.05)。50 μg/L处理组在第1天POD总活性的达到最高峰值,与对照组相比上升了45.53%,说明低浓度的硫丹可以刺激POD酶的活性升高。而第3天则是在150 μg/L处理组出现轻微上升,之后活性立即下降,下降幅度很明显。至第七天与对照组相比,各浓度组POD总活性分别下降了18.80%、31.61%、60.48%、70.04%。高浓度处理组250和350 μg/L的POD总活性具有显著的剂量-时间效应关系,说明高浓度的硫丹胁迫下,会造成孔石莼的POD酶失活。
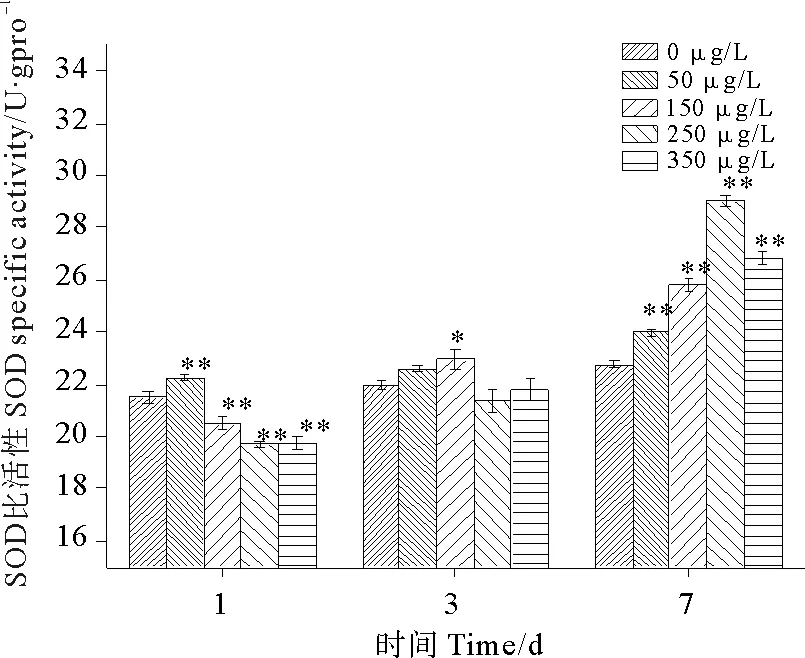
图10 硫丹对孔石莼SOD比活性的影响Fig.10 Effects of treatments with different endosulfan concentrations on SOD specific activity of Ulvapertusa
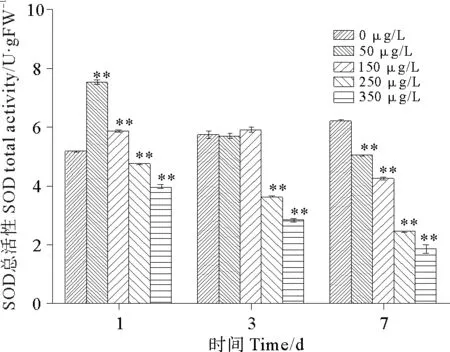
图11 硫丹对孔石莼POD总活性的影响Fig.11 Effects of treatments with different endosulfan concentrations on POD total activityof Ulvapertusa
图12为硫丹对孔石莼POD比活性的影响。孔石莼POD比活性也呈现出低浓度促进作用,高浓度抑制作用。50 μg/L处理组在暴露第一天出现促进作用,与对照组相比,POD的比活性上升了33.24%。在硫丹暴露第2天,50和150 μg/L浓度组均表现出促进作用,与对照组相比分别升高了6.94%、20.23%。在暴露第7天,50和150 μg/L浓度组与对照组之间无显著性差异(P>0.05)。随着硫丹暴露时间的延长,150 μg/L浓度组的POD比活性在第三天先升高之后到第7天又下降,但仍然比第1天的比活性高。

图12 硫丹对孔石莼POD比活性的影响Fig.12 Effects of treatments with different endosulfan concentrations on POD specific activityof Ulvapertusa
2.3.2 硫丹与pH耦合对孔石莼抗氧化酶活性的影响 图14 为pH与不同浓度硫丹耦合对孔石莼SOD比活性的影响。在环境pH和低pH条件下,孔石莼SOD比活性的变化趋势表现较为一致,均为先升高再降低。环境pH条件下,暴露第7天时250 μg/L处理组出现SOD比活性的峰值,相对于对照组增加了27.58%。低pH条件下,暴露第1天时50 μg/L处理组的SOD比活性先上升随后开始下降;在暴露第3天和第7天都是在150 μg/L处理组出现峰值,相对于对照组分别增加了18.35%和37.18%。在第7天低pH的各处理组除了L-0处理组之外,其他各浓度组与对照组均有极显著性差异(P<0.01)。
从pH与不同浓度硫丹耦合对孔石莼SOD活性主体间效应分析来看,硫丹对孔石莼SOD比活性有作用,且在第1天、第3天和第7天均具有极显著性影响(P<0.01)。pH仅在第3天和第7天对SOD比活性产生影响。硫丹与pH的交互作用仅在第七天发挥显著作用(P<0.05),第1天和第3天均无显著影响(P>0.05)。
图14为pH与不同浓度硫丹耦合对孔石莼POD比活性的影响。在环境pH和低pH条件下,孔石莼POD的比活性的变化趋势一致表现为先升高再降低。且POD对硫丹胁迫的响应相较于SOD更加敏感。环境pH条件下,在暴露第1天时50 μg/L处理组POD比活性达到峰值,相对于对照组增加了33.22% ;150 μg/L处理组在暴露第3天时出现峰值,与对照组相比增加了20.12% ;至第7天各处理组POD比活性没有明显升高,高浓度时显著降低。低pH条件下,暴露第1天、第3天和第7天均是50 μg/L处理组出现峰值,之后开始下降,相对于对照组分别增加了24.07%、33.74%和29.32% 。低pH条件下,除L-0、L-150 μg/L (7 d)、L-250 μg/L(1 d)3个处理组之外,其他处理组与对照组相比均有极显著性差异(P<0.01)。

图13 pH与不同浓度硫丹耦合对孔石莼SOD比活性的影响Fig.13 Effects of treatments coupling with different endosulfan concentrations and pH on SOD activity of Ulvapertusa
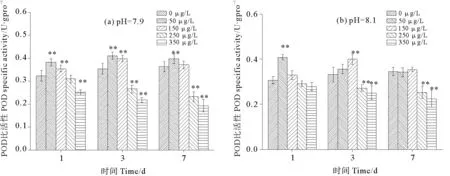
图14 pH与不同浓度硫丹耦合对孔石莼POD比活性的影响Fig.14 Effects of treatments coupling with different endosulfan concentrations and pH on POD activity of Ulvapertusa
pH与不同浓度硫丹耦合对孔石莼POD活性主体间效应的检验后发现,硫丹对孔石莼POD比活性发挥作用,且在第1天、第3天和第7天均具有极显著性差异(P<0.01)。pH在上述3个时间段对POD比活性均没有显著影响(P>0.05)。硫丹与pH的交互作用在第1天、第3天和第7天均发挥了作用,对POD比活性具有极显著影响(P<0.01)。
3 讨论
3.1 硫丹胁迫及与pH耦合对孔石莼生长的影响
植物的生长状况会受诸多外界环境因素的制约与影响[13],而生长的响应则是一系列生理生化参数受到胁迫的综合表现[14]。本实验中发现在高浓度硫丹胁迫下,孔石莼叶片出现严重白化现象,硫丹单独胁迫时,孔石莼的相对生长率均随硫丹浓度的升高而呈现先升高后下降的趋势,相对生长率在硫丹浓度为50 μg/L 时达到最高,而暴露时间进一步延长后,硫丹对孔石莼的生长的促进作用逐渐减弱而抑制作用逐渐加深,这也与许多研究者发现低浓度有机污染物对藻类不但没有毒害作用,反而有促进作用的结论一致。Stebbing[15]认为这种增益现象是生物的一种“毒物的兴奋效应”,是自我保护的一种机制有机污染物促使藻类产生“兴奋效应”的原因在于低浓度下,毒害和降解两个过程同时存在,其中降解过程占主导地位,因而污染物对藻类伤害小,而在高浓度硫丹胁迫下,孔石莼的相对生长率远远低于对照组并且出现负增长。这可能是因为硫丹具有较高脂溶性,能渗透到藻体细胞壁并溶解于细胞膜中,破坏细胞的内含物组成,进而引起细胞萎缩甚至破裂,宏观上表现为生长上受到抑制,藻体颜色出现发白。
pH是影响藻类生长繁殖重要的环境因子。其对藻类生长产生的影响取决于光合作用与呼吸作用间的碳收支平衡,本实验中发现低pH环境时孔石莼的相对生长率高于 pH 为8.2的对照组;而光合色素含量相对于对照组有所下降。可能是在低 pH 下藻类细胞的渗透压调节受到干扰,通过细胞的离子(如K+、Na+)流动失控,从而导致细胞功能的紊乱,细胞色素含量下降,光合作用速率降低的同时呼吸作用受到抑制影响更大。这与文献中提到的海洋酸化导致海洋生物量的增加,但初级生产力可能并未有相应的增长相一致。
3.2 硫丹胁迫及与pH耦合对孔石莼光合色素的影响
光合作用色素是植物进行光合作用的物质基础,也是间接反映进行光合作用生物的生物量指标,其含量变化能较好地反映生物各阶段生长发育正常与否。目前的研究表明,藻类在遭受环境胁迫时,反映其光合强度的叶绿素、类胡萝卜素等色素会发生变化[16],这可能与色素前体的周转和转化有关。硫丹单独胁迫时,50 μg/L处理组能够刺激孔石莼体内光合色素含量增加,而高浓度硫丹则会降低光合色素含量,且叶绿素a受到的抑制作用尤其显著,藻体的光合作用也受到了明显的抑制作用;而低 pH 与硫丹耦合胁迫的各处理组相对于仅受硫丹胁迫的各处理组光合色素的含量下降幅度更大,表明 pH 加重了硫丹对孔石莼的毒性效应,两者关系表现为协同作用。其可能的原因是硫丹的理化性质受环境因子pH的影响,在低 pH 环境下硫丹在水中的半衰期会增加,降解的更缓慢。因此在低 pH 硫丹处理组中光合色素的含量整体比环境pH条件下各处理组色素含量低。Alberte[17]认为逆境胁迫下叶绿素含量降低的主要原因是叶绿体片层中捕光 Chla/b- Pro 复合体合成受到抑制。在低浓度硫丹单独胁迫时,孔石莼中3种色素含量都会增加,这可能是光合色素合成系统对硫丹毒害的一种应激反应,可以作为一种有效保护机制抵御硫丹对藻体的毒害,光合色素含量的增加使藻体能够吸收更多的光能进行光合作用,从而为藻体生长代谢提供能量;同时,类胡萝卜素可以作为抗氧化剂清除自由基和单线态氧,防止膜脂的过氧化,维持膜结构的稳定。
光合器官是植物细胞内超氧自由基等的主要来源之一,而光合色素及与之结合的类囊体膜均具有不饱和多烯结构,极易受 ROS 的攻击。本实验中, 350 μg/L的硫丹单独胁迫下孔石莼叶绿素a和类胡萝卜素的含量均有较为明显的下降,可能是由于孔石莼受硫丹胁迫后,细胞内活性氧的产生和清除间的平衡被破坏,导致活性氧积累,损伤了细胞膜的结构和功能。唐学玺等的研究表明,久效磷胁迫下扁藻叶绿素a的降解与活性氧的损伤有关[18]。
3.3 硫丹胁迫及与pH耦合对孔石莼抗氧化酶活性的影响
藻类抗氧化酶保护系统主要包括SOD、CAT和POD等,超氧化物歧化酶(SOD)是细胞内 ROS 清除系统的主要成分,是需氧生物长期进化过程中形成的一套复杂的抗氧化保护系统的成分之一,与 CAT、POD 等酶抗性系统一起协同防御活性氧或其他过氧自由基对细胞膜系统的伤害。Wang等通过实验发现壬基酚暴露处理 12 天的铜绿微囊藻的GST和SOD 活力均有升高,但是 GST 活力变化比 SOD 活力变化更明显,认为 GST 在消除毒物毒性上贡献比 SOD 要大[19]。本实验结果显示,硫丹单独胁迫时孔石莼体内SOD和POD酶活性都是随着丹浓度的递增呈现先升高后下降的趋势,在50 μg/L硫丹胁迫下,SOD的总活性升高;50和150 μg/L处理组的POD总活性均有不同程度的上升,并且SOD比活性变化的更显著。在350 μg/L硫丹胁迫下,SOD和POD的总活性明显下降,但POD在高浓度胁迫环境下活性比SOD下降的更低,说明POD对硫丹胁迫更加敏感。
岳文杰等研究显示,高浓度的氯氰菊酯胁迫后龙须菜的可溶性蛋白含量受到抑制,本实验中,孔石莼在高浓度硫丹胁迫下生长受到严重胁迫,可溶性蛋白含量也表现出显著降低, SOD和POD活性是相对的增加[20]。Prasad等研究结果显示,在硫丹刺激下,丝状蓝菌的抗氧化酶SOD、CAT、POD的比活性都显著增强[21]。硫丹浓度为250 μg/L时,孔石莼SOD的比活性出现上升趋势,而150 μg/L硫丹处理组的POD活性上升幅度更加明显。
本实验发现低浓度硫丹能使藻体产生诱导反应,增加抗氧化酶的活性,以抵抗氧化胁迫。而高浓度硫丹胁迫下孔石莼体内的抗氧化系统发生紊乱,抗氧化酶活性降低,使得藻体对硫丹胁迫的抗性机制丧失。这些在微藻中也有相同报道,沈忱等用苯并芘胁迫三种海洋微藻,发现随着胁迫时间的延长 BaP 的诱导作用逐渐消失,表现出抗氧化酶活力先升高后降低的总趋势[22]。这种现象在其他植物中也有广泛的报道。实验结果表明孔石莼对硫丹的胁迫具有一定的耐受能力,轻度胁迫下,孔石莼的抗氧化防御系统产生适应性的诱导反应;而当外界胁迫加重时,会对抗氧化系统产生抑制效应。
当硫丹和pH共同胁迫时,低 pH 硫丹处理组的抗氧化酶的活性对硫丹的敏感程度与环境 pH 处理组不同。实验结果显示环境 pH 处理组SOD 的比活性在250 μg/L时上升幅度最大,而低 pH 处理组在150 μg/L时已经达到最大峰值,说明低 pH 条件下SOD 的比活性对硫丹更加敏感。POD 的比活性在环境 pH 处理组在硫丹暴露第1天达到顶峰,低 pH 处理组则在硫丹暴露第3天达到最大值,说明相对于环境 pH 条件,POD 对硫丹胁迫的敏感性在低 pH 条件下减小。
4 结论
(1) 硫丹对孔石莼单独胁迫时,低浓度的硫丹对孔石莼的生长、光合色素以及抗氧化酶系统都表现出“毒物兴奋效应”,硫丹浓度为50 μg/L时促进效应最为显著,这是藻类的一种自我保护机制。具体表现为相对生长率升高、光合色素含量增加以及抗氧化酶活性的上升;高浓度硫丹胁迫下,由于活性氧的过量累积,孔石莼的生长代谢过程均受到了不同程度的损害。孔石莼的生长受到抑制,相对生长率下降甚至出现负增长,藻体颜色发白,光合色素含量出现明显下降,抗氧化酶活性受到显著抑制。
(2) 硫丹与环境因子pH耦合实验中,低pH条件会促进孔石莼的生长,但降低了孔石莼光合色素的含量。在低pH条件下,硫丹对孔石莼的刺激作用更加显著,表现为低浓度更加促进,高浓度时抑制作用加剧。低 pH 条件下各处理组抗氧化酶对硫丹的敏感程度与环境 pH 处理组不同,SOD对硫丹更为敏感而POD对硫丹的敏感性则有所下降。硫丹与pH的交互作用在孔石莼不同生理过程中发挥的效应程度不同,考虑到pH对硫丹的一些理化性质也会产生影响,因此在暴露的各阶段,孔石莼对胁迫的响应也有所不同。
另外针对本实验中硫丹对大型海藻在个体和生理生化上的生态毒理影响,硫丹分子层面的作用机理和调控方式将是本文下一步的研究内容。
