8种绿化树种光合特性及叶片解剖结构比较
2019-02-14冮慧欣王嘉琪黄春岩王秀伟
冮慧欣 王嘉琪 黄春岩 王秀伟
(东北林业大学林学院,哈尔滨 150040)
城市绿化作为城市生态建设中不可缺少的重要组成部分之一,生态园林不仅包含了园林绿化的美化作用,其更重要意义在于充分利用植物的释氧和吸收CO2的能力,及降温增湿、滞尘、杀菌、抗污、降噪等生态功能[1~2]。这些生态功能是通过植物的生理活动实现的,但是光合能力在种间和基因型间的变化很大,这些差异通常与代谢和(或)叶片的解剖结构的性质有关[3]。种内光合速率差异是由测定方法和环境对叶片解剖结构的先决影响造成的[4]。叶片是对环境变化较敏感且变异性和可塑性较大的器官,不同环境中植物叶片厚度、角质层、表皮细胞、栅栏组织和海绵组织等均发生相应变化,不同种植物叶片解剖结构存在着很大的差异[5~9]。已有的研究表明叶片厚度对生理指标影响显著,小麦旗叶厚度与净光合速率、气孔导度和蒸腾速率呈显著正相关[10],大豆叶片光合速率取决于叶片厚度和单位面积栅栏细胞数[11]。表皮和表面角质层蜡质通过增加扩散的阻力来降低蒸腾[12]。气孔是植物赖以调节自身和外界环境发生关系的一种重要结构,是植物体内和环境气体交换与水分蒸腾的一个主要通道,叶表面气孔密度、大小和分布受植物种类和环境影响[13]。结构是功能的基础,植物结构的变化必然影响到生理生态功能的改变[14]。土壤含水量、不同光照对蒸腾速率和光合速率的影响显著[15~16]。环境条件的差异会导致植物叶片解剖结构的变化[8],阳生植物和阴生植物或阳生叶片和阴生叶片解剖结构与生理指标差异显著[17],除了环境条件差异外,是否城市绿化树种叶片解剖结构的差异导致其光合生理参数的差异还不清楚。为此,本研究比较测定了哈尔滨市8个树种的叶片解剖结构和光合生理相关因子,旨在比较分析不同树种间叶片解剖结构变化与其光合生理特征的关系。
1 材料和方法
1.1 实验材料
选择东北林业大学校园内的8种生长状态良好的树木:暴马丁香(Syringareticulata(Blume)Hara var.mandshurica(Maxim.)Hara)、东北溲疏(Deutziaamurensis(Rgl) Airy-Shaw)、紫丁香(SyringaoblataLindl.)、山楂叶悬钩子(RubuscrataegifoliusBge.)、榛(CorylusheterophyllaFisch. ex Trautv.)、金银忍冬(Loniceramaackii(Rupr.)Maxim.)、梓树(CatalpaovataG.Don)和银中杨(Populusalba×P.berolinensisDippel)为研究对象,每树种选择3株,每株选取3片生长状态良好的叶片进行光合生理指标测定和解剖结构观察。
1.2 生理指标测定
在生长旺季的2012年8月中旬,在天气晴朗的上午,平均温度30.97℃,采用便携式光合仪(Model6400;Li-Cor Inc.,Lincoln,NE,USA)测定8个树种叶片的净光合速率、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)等参数。采用LED红/蓝光源(6400-02B)控制叶室内的光强梯度为1200、1000、800、600、400、200、100、50、20、0μmol·photons·m-2·s-1。
1.3 叶片解剖结构的观察
生理指标测定后,摘取的叶片切成5mm长的小段,用FAA固定,参照李和平方法制作石蜡切片[18]。用显微镜观察石蜡切片并应用显微图像处理分析软件PHMIAS2008Cs Ver2.2测量表皮细胞厚度、叶片厚度、栅栏组织和海绵组织的厚度。
采用指甲油印迹法,在叶片上、下表皮中部靠近主脉的两侧快速涂上一层薄薄的透明指甲油,约1cm2,待其风干结成膜后,然后轻轻剥下,把所有叶表皮膜的指甲油层粘在透明胶带上,制成临时装片,在显微镜下对单位面积气孔进行计数,对所测得的各树种气孔数量进行统计,然后计算气孔密度。
1.4 统计分析
(1)
式中:Pn为净光合速率,α是表观量子效率,I为光通量密度;θ为曲角,Pmax是最大净光合速率,Rd是光下呼吸速率[19]。
P=Pmax[1-exp(1-αI/Pmax)]
(2)
并采用公式2、3分别求出光补偿点(LCP)与光饱和点(LSP)。
LCP=Pmaxln(C0)/α
(3)
LSP=Pmaxln(100C0)/α
(4)
式中:C0为度量弱光下净光合速率趋于零的指标。
采用Excel2010(Microsoft Inc,USA)对数据进行整理,使用SPSS16.0for windows(SPSS Inc,USA)统计分析系统进行光响应曲线拟合,对最大光合速率、呼吸速率,表观量子效率进行估计,树种间生理指标和差异和解剖结构指标的差异采用单因素方差分析(One-Way ANOVA),对方差分析差异有统计学意义的指标,用Duncan法进行组间多重比较。
2 结果与分析
2.1 树种间生理指标的差异
树种间叶片最大光合速率、气孔导度、胞间CO2浓度、蒸腾速率、光饱和点差异显著(P<0.05)(表1)。8个树种叶片光合速率的变异系数为31.02%,梓树叶片最大光合速率在大,为19.81μmol·m-2·s-1,金银忍冬和榛叶片最大光合速率最低,分别为9.77和9.70μmol·m-2·s-1。胞间CO2浓度的变异系数为13.45%,暴马丁香的胞间CO2浓度最大,为293.7μmol·mol-1,紫丁香胞间CO2浓度最低,为196.0μmol·mol-1,蒸腾速率的变异系数为44.50%,梓树叶片的蒸腾速率最大,为7.9mmol·m-2·s-1,暴马丁香和金银忍冬叶片的蒸腾速率最低,为2.84和2.89mmol·m-2·s-1。光饱和点的变异系数为24.62%,梓树叶片的光饱和点最大,为1643.89μmol·m-2·s-1,暴马丁香叶片的光饱和点最低,为904.8μmol·m-2·s-1。梓树叶片的气孔导度最大,为0.53mol·m-2·s-1,其余7个树种叶片气孔导度差异不显著。8个树种叶片的光补偿点(LCP)和呼吸速率(Rd)差异不显著(P<0.05)(表1)。
2.2 树种间叶解剖结构的差异
8个树种叶片的横切面的基本结构由叶肉细胞、上下表皮、栅栏组织、海绵组织、维管束等组成,栅栏组织细胞呈长柱状,海绵组织由排列疏松的不规则细胞组成。8个树种叶片表皮厚度、栅栏组织厚度、上表皮气孔密度和下表皮气孔密度差异显著(P<0.05)(表2)。8个树种表皮厚度的变异系数为27.03%,梓树叶片的表皮厚度最大,为24.65 μm,银中杨叶片的表皮厚度最小,为12.82 μm。8个树种栅栏组织厚度的变异系数为30.33%,紫丁香的栅栏组织厚度最大,为108.6 μm,东北溲疏栅栏组织厚度最小,为50.0 μm。8个树种叶片厚度和海绵组织厚度差异不显著(P<0.05)。8个树种下表皮气孔密度差异显著(P<0.05),变异系数为42.33%,梓树叶片下表皮气孔密度最大,为60.2 个·mm-2,东北溲疏叶片下表皮气孔密度最小,为15.0 个·mm-2。8个树种中仅有紫丁香和银中杨叶片上表皮具有气孔,紫丁香叶片上表皮气孔密度为3.2 个·mm-2,银中杨上表皮气孔密度为22.8 个·mm-2。
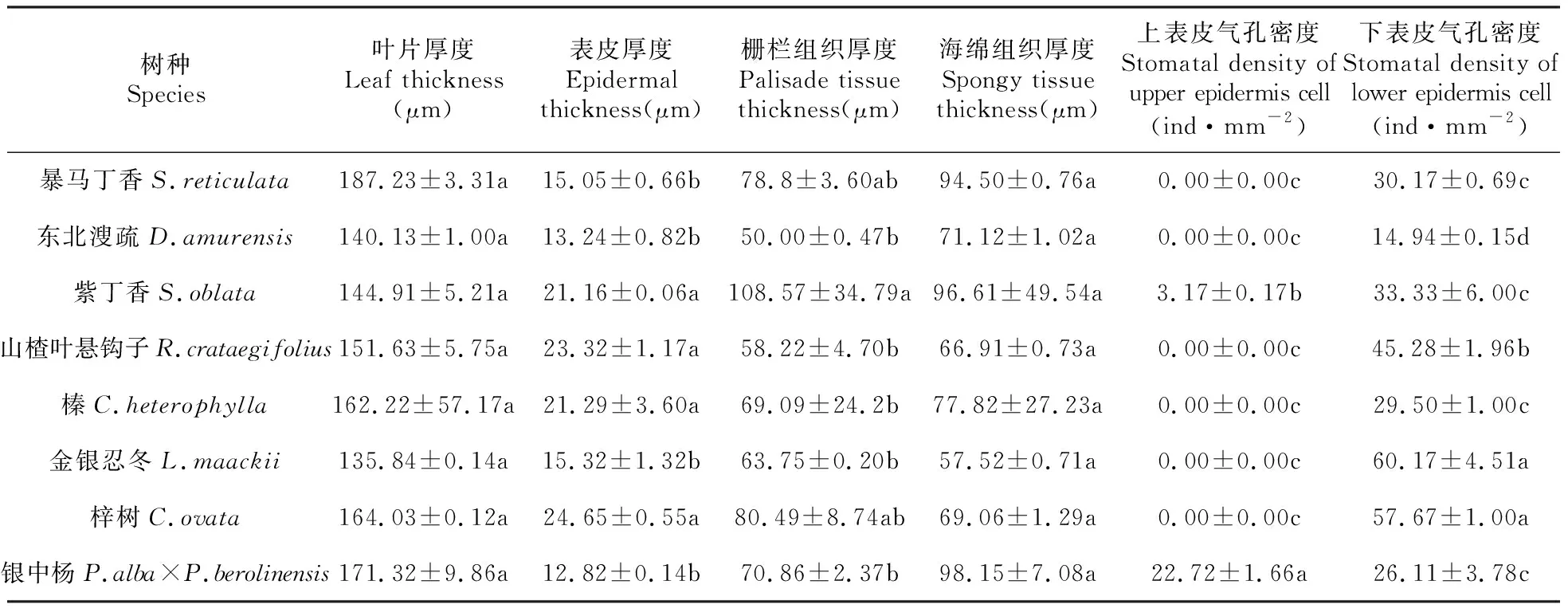
表2 8个树种叶片叶解剖结构差异
2.3 叶片解剖结构与生理指标的关系
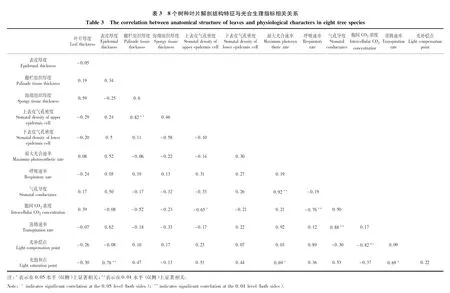
由表3可见,8个树种光饱和点与叶表皮厚度显著正相关(P<0.01),相关系数为0.78。胞间CO2浓度与上表皮气孔密度显著负相关(P<0.05),相关系数为-0.65。最大光合速率、呼吸速率、蒸腾速率和光补偿点与表皮厚度、栅栏组织厚度、海绵组织厚度、上表皮气孔密度和下表皮气孔密度相关均不显著(P>0.05)。胞间CO2浓度与表皮厚度、栅栏组织厚度、海绵组织厚度和下表皮气孔密度相关均不显著(P>0.05)。光饱和点与栅栏组织厚度、海绵组织厚度、上表皮气孔密度和下表皮气孔密度相关均不显著(P>0.05)。
3 讨论
8个树种间叶片解剖结构(栅栏组织厚度、海绵组织厚度、上表皮气孔密度和下表皮气孔密度)和光合生理指标(最大光合速率、气孔导度、胞间CO2浓度、蒸腾速率和光饱和点)均存在显著差异。光合速率受到许多植物因素的影响,包括叶肉固碳能力的遗传差异、叶片解剖结构、叶龄(有时为树龄)、气孔体积、气孔密度和气孔开度调控[3]。本研究中光饱和点与叶表皮厚度显著正相关,已有的研究表明叶片表皮毛和厚角质膜发达能够适应或抵御外界环境(水分和光照)的胁迫,维持植物正常的生理生态机能[20],因此表皮较厚能增强对太阳辐射的反射,即减少对太阳辐射能的吸收,可能间接促进了光饱和点的升高。本研究中胞间CO2浓度与上表皮气孔密度显著负相关,气孔是植物与外界环境进行CO2和H2O等气体交换的重要通道,直接决定着植物的蒸腾和光合作用[21]。已有研究表明大气CO2浓度与土壤氮素供应增加可影响叶片表皮组织分化,增加叶片上表皮气孔出现的比例,促进叶片的光合速率树木生长[22]。单提波研究表明高气孔密度水稻的最大光合速率为比中、低气孔密度水稻分别高4.3%和19.6%[23]。
已有研究表明油橄榄(OleaeuropaeaL.)栅栏组织厚度、 栅栏/海绵组织与净光合速率极显著正相关,在一定时期和范围内可作为判断光合能力的重要指标[24],但是本研究中未发现光合生理指标与栅栏组织厚度、海绵组织厚度的相关关系。光合特征存在种和基因型的差异,但是也受到叶片类型、叶龄、空气CO2浓度和温度的影响[3]。已有叶片结构与生理指标间关系研究是针对同一种植物进行研究,本研究选择了8个树种进行研究,所以呈现出更复杂的趋势。本研究的结果表明,在树种之间,叶片解剖结构能够影响生理活动。但是值得注意的是,本研究只是选择成熟叶片进行研究,我们不清楚生理指标是如何随着叶片结构的发育而变化的,以及在不同的发育阶段,树种在生理和叶片结构上的差异是否会更小或更大。虽然仍有许多关于叶片结构对生理的影响的机理需要研究,但我们认为,对叶片结构的研究,可以更好地理解光合速率、呼吸速率和蒸腾速率等生理指标的变化。