基于SSR标记的不同优势等级云南松遗传多样性分析
2019-02-14蔡年辉许玉兰王亚楠汪梦婷李根前
蔡年辉 许玉兰 王亚楠 陈 诗 汪梦婷 李根前*
(1.西南林业大学西南山地森林资源保育与利用教育部重点实验室,昆明 650224; 2.西南林业大学云南省高校林木遗传改良与繁育重点实验室,昆明 650224)
云南松(PinusyunnanensisFranch.)是我国西南地区重要的造林树种,在区域林业经济生产和生态环境建设中发挥着重要的作用。在林分中,个体的生长因遗传基础或环境因素(对有限的光、水、养分等资源的竞争)而产生了分化[1~2],生长表现参差不齐[3],存在优势度的差异,其相对大小或冠层位置常用优势木、中等木、被压木或劣势木等林木等级来描述[2,4~7]。在林木经营管理中,可能因优势度的不同采取不一样的经营管理措施。对于天然更新的异龄林,在空间上呈现一定的层次结构,可将林层分为林冠层和更新层,其中林冠层为大量生物提供栖息地并主导着森林生态系统功能的发挥,例如固碳增汇、雨量分配、涵养水源及调节区域气候变化等,并承载了森林生物多样性的主体部分[8~9];更新层是决定未来林分结构的重要因子,也是林分可持续循环的决定性因子[10],其物种的演替是森林群落结构发生变化的主导驱动因子[11],按林层分布探讨其遗传变异特征对于全面系统地估测遗传变异的动态变化有重要意义。
了解遗传多样性和遗传变异结构对于科学制定其遗传资源保护策略具有重要的指导作用,并有利于合理的开发利用该遗传资源[12]。因此,有必要对林木进行优势度划分,进而开展遗传多样性的研究。目前针对云南松遗传多样性的研究较少,且主要集中在天然群体遗传变异及其空间格局形成机制[13~15],研究揭示了云南松群体遗传多样性丰富、群体间遗传分化不明显,也不存在明显的遗传结构,地理隔离对云南松群体遗传距离的影响低于生态隔离的作用,但各生态因子对云南松群体的作用不一样,进一步依据云南松群体的遗传变异特点,初步提出云南松群体的遗传多样性的优先保护单元。但从林木优势度差异研究遗传多样未见报道。SSR分子标记技术具有重复性好、多态性高、共显性遗传等优点[16],在种质资源评价、图谱构建、基因挖掘等方面的研究发挥着重要作用[16~19]。为此,本试验以云南松林分为试验对象,根据树高和胸径(地径)优势度不同划分为优势木、中等木和劣势木,应用SSR分子标记技术,比较不同生长优势等级云南松种质的遗传多样性,研究结果可为深入了解种质遗传完整性的时空变化特征奠定基础,为云南松可持续生产营林措施制定提供依据,进而指导林业生产实践。
1 材料与方法
1.1 样品采集
试验地林分为云南松天然更新林,林分生长良好,冠层郁闭。野外调查以样地法和每木检尺为基础,样地选择采用随机抽样法,设置3个30 m×30 m样地,样地位于云南省双江拉祜族佤族布朗族傣族自治县境内,各样地的地理坐标分别为:样地一:99°46′41.75″E~99°46′58.35″E、23°22′54.12″N~23°22′57.99″N;样地二:99°45′32.67″E~99°45′33.81″E、23°20′30.27″N~23°20′31.53″N;样地三:99°46′54.10″E~99°46′55.86″E、23°22′58.88″N~23°22′59.88″N。对样地内的每木进行检尺,测定时将各植株按生长状况划分为林冠层和更新层,胸径大于或等于5 cm的植株视为林冠层;将胸径小于5 cm的植株视为更新层[20]。林冠层测量树高和胸径,更新层测量树高和地径。试验地为云南松林,树种单一,结构简单,故按树高和胸径(地径)为主要分类因子,将样地内林冠层和更新层的各样株按树高和胸径(地径)进行优势等级分级。对3个样地内实测每木的树高和胸径(地径)整理,确定级差,将各样株归入相应的级别[21]即得到分级结果(表1)。

表1 各样地不同优势等级树高与胸径

表2 SSR扩增引物信息表
采集当年生针叶,每个样品采集的针叶分为两份,一份新鲜针叶直接进行DNA提取,另一份采用硅胶干燥贮藏备份。提取的DNA进行SSR分子标记分析,分别比较优势木、中等木和劣势木3个不同优势等级林木间的遗传多样性,探讨它们之间的遗传变异特征。
1.2 DNA提取与SSR-PCR扩增
云南松针叶DNA的提取采用CTAB提取法[22],由于云南松自身的生物特性,为了提高提取效果,本次研究增加了CTAB的用量,即采用4×CTAB[23]。研究中共使用引物15对,SSR引物信息见表2。SSR扩增反应体系和扩增程序参照前期研究[29],扩增产物由北京睿博兴科生物技术有限公司采用荧光分型检测。
1.3 数据分析
数据的收集与分析同前期研究[15],数据收集采用GeneMarker V2.2.0,以GS500LIZ为标准收集数据,设置相应参数,人工逐一校对,Excel表格输出基因型分子量数据。利用Convert 1.3.1对数据进行转换,以满足不同分析软件的输入要求。采用GenALEx 6.4计算等位基因数(Na)、有效等位基因数(Ne)、Shannon’s信息指数(I)、观测杂合度(Ho)、期望杂合度(He)等遗传多样性参数,运用Arlequin 3.0进行分子方差分析(AMOVA,Analysis of Molecular Variance)。利用GenALEx 6.4计算遗传距离,运用NTSYS-pc 2.10s对不同生长优势等级进行聚类,聚类采用算术平均数非加权成组配对法(UPGMA,Unweighted Pair Group Method with Arithmetic average),生成聚类图。不同优势等级林木间遗传多样性的差异显著性采用单因素方差分析,由SPSS17.0软件进行运算。图表制作采用Excel软件。
2 结果与分析
2.1 遗传多样性分析
将采集的各样本植株进行SSR遗传多样性分析(图1)。结果表明,从林冠层来看,三个样地优势木、中等木和劣势木的Shannon’s信息指数平均值分别为0.762、0.759和0.741,以优势木的最高,观测杂合度也表现为优势木>中等木>劣势木;同样的三个样地优势木、中等木和劣势木的期望杂合度平均值分别为0.423、0.402和0.412,以优势木的最高,有效等位基因数也表现为优势木>劣势木>中等木。总体来看,不同优势等级林木间的遗传多样性差别较小,以优势木的略高。
更新层的遗传多样性变化表现有所不一样,三个样地的优势木、中等木和劣势木的Shannon’s信息指数平均值分别为0.774、0.661和0.736,以优势木的最高;同样地,三个样地的优势木、中等木和劣势木的期望杂合度平均值分别为0.410、0.374和0.403,以优势木的最高。等位基因数和有效等位基因数也表现为优势木>劣势木>中等木,观测杂合度表现为中等木>劣势木>优势木。相比较林冠层而言,更新层不同优势等级林木间的遗传多样性总体差别稍大些。
进一步对各遗传多样性指标进行差异显著性检验,结果表明(表3),无论在林冠层还是更新层,不同优势等级林木间的遗传多样性差异均不显著(P>0.05)。
对不同优势等级内各指标的变异波动进行分析(图2),在林冠层,优势木5个遗传多样性指标的变异系数为8.04%~18.14%,平均为12.55%;中等木为2.56%~19.32%,平均为8.58%;劣势木为2.68%~9.13%,平均4.71%,各指标平均值在不同分化等级林木之间表现为优势木>中等木>劣势木。更新层优势木5个遗传多样性指标的变异系数为3.78%~13.72%,平均为8.41%;中等木为7.64%~22.17%。平均为14.81%;劣势木为8.20%~22.00%,平均12.75%。更新层的总体表现与林冠层有所不同,以优势木最低,而以中等木最大。总体来看,不同生长优势等级各指标的变化格局不一。
2.2 遗传距离分析
为进一步比较不同优势等级云南松林木间的遗传关系,计算不同优势等级云南松林木间的遗传距离(表4)。从表中可以看出,林冠层三个样地各等级林木间遗传距离分别为0.016~0.043、
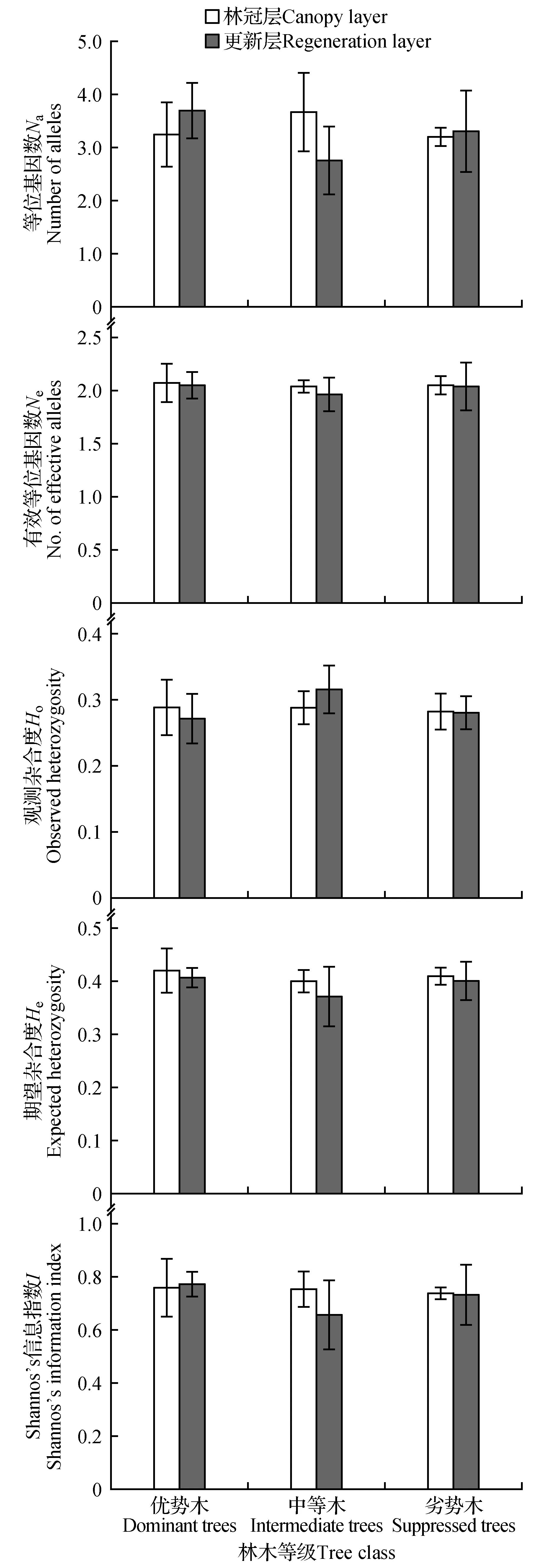
图1 不同优势等级云南松遗传多样性Fig.1 Genetic diversity in different dominance hierarchies of P.yunnanensis
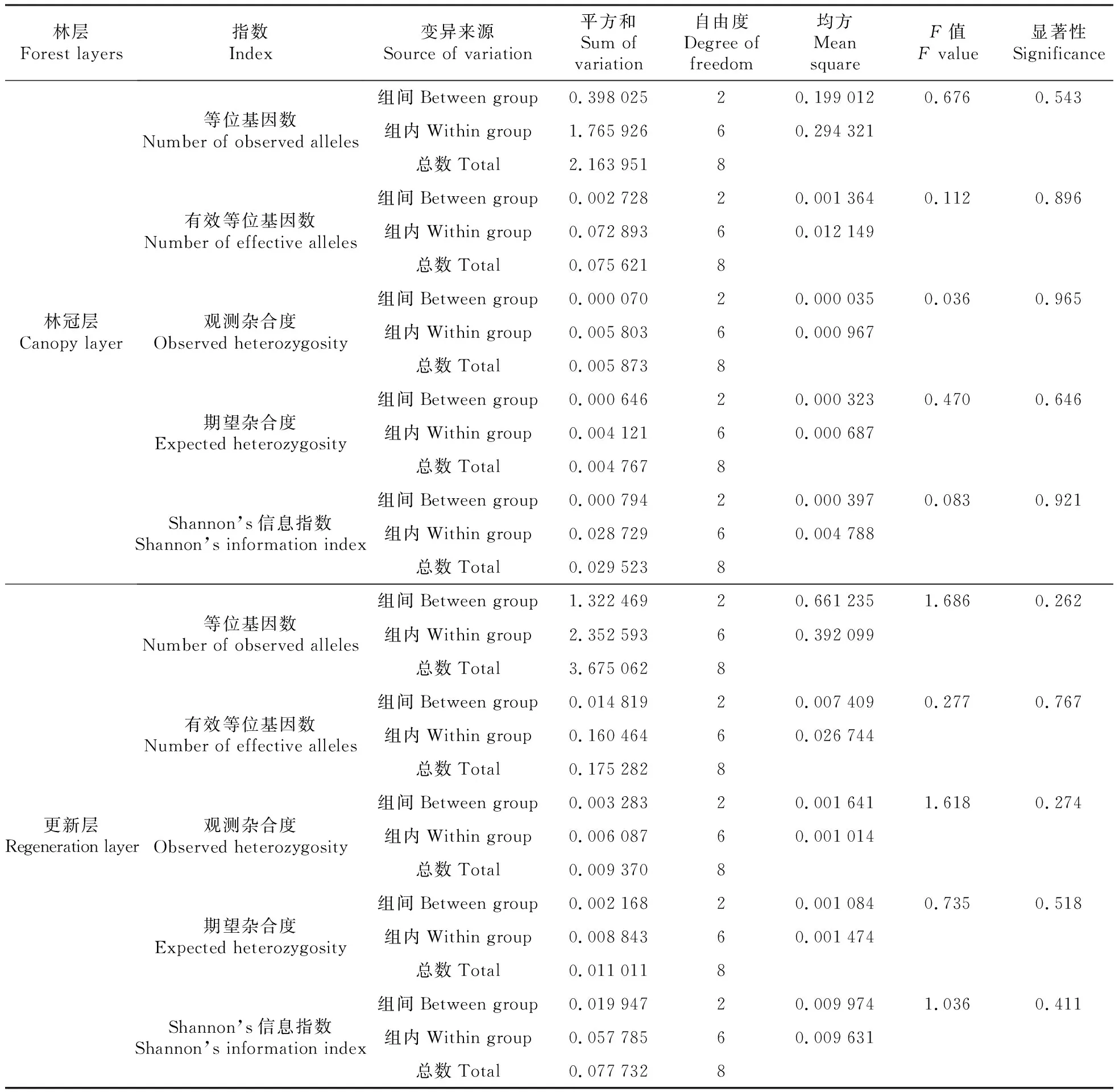
林层Forest layers指数Index变异来源Source of variation平方和Sum of variation自由度Degree of freedom均方Mean squareF值F value显著性Significance林冠层Canopy layer等位基因数Number of observed alleles有效等位基因数Number of effective alleles观测杂合度Observed heterozygosity期望杂合度Expected heterozygosityShannon’s信息指数Shannon’s information index组间Between group0.39802520.1990120.6760.543组内Within group1.76592660.294321总数Total2.1639518组间Between group0.00272820.0013640.1120.896组内Within group0.07289360.012149总数Total0.0756218组间Between group0.00007020.0000350.0360.965组内Within group0.00580360.000967总数Total0.0058738组间Between group0.00064620.0003230.4700.646组内Within group0.00412160.000687总数Total0.0047678组间Between group0.00079420.0003970.0830.921组内Within group0.02872960.004788总数Total0.0295238更新层Regeneration layer等位基因数Number of observed alleles有效等位基因数Number of effective alleles观测杂合度Observed heterozygosity期望杂合度Expected heterozygosityShannon’s信息指数Shannon’s information index组间Between group1.32246920.6612351.6860.262组内Within group2.35259360.392099总数Total3.6750628组间Between group0.01481920.0074090.2770.767组内Within group0.16046460.026744总数Total0.1752828组间Between group0.00328320.0016411.6180.274组内Within group0.00608760.001014总数Total0.0093708组间Between group0.00216820.0010840.7350.518组内Within group0.00884360.001474总数Total0.0110118组间Between group0.01994720.0099741.0360.411组内Within group0.05778560.009631总数Total0.0777328
0.013~0.051和0.026~0.048,其高值在样地一和样地三均表现为优势木与劣势木之间、样地二表现为中等木与劣势木之间,三个样地平均分别为0.032、0.036和0.037,总体平均为0.035。更新层三个样地遗传距离分别为0.016~0.020、0.026~0.028和0.033~0.116,其高值在样地一和样地三均表现为劣势木与中等木之间、样地二表现在中等木与优势木之间,三个样地平均分别为0.019、0.027和0.079,总体平均为0.041。综合来看,无论是林冠层还是更新层,其各等级林木间的遗传距离均较低,且出现无一致的规律性,不同等级林木间的遗传距离较近。
2.3 聚类分析
为确定各等级林木间的遗传关系,根据遗传距离,采用NTSYS-pc 2.10s软件中的算数平均数非加权成组配对法(UPGMA)构建聚类图。从图3中看出,三个样地林冠层的聚类呈现随机性,分别以优势木与中等木、优势木与劣势木、中等木与劣势木先聚再与另一优势等级林木聚类。同样地,在三个样地更新层的聚类也呈现较大的随机性,其中样地一和样地三以优势木和劣势木相近,而样地二以中等木与劣势木相近。但总体来看,各个样地不同等级林木间的聚类均比较随机,聚类与林木生长优势等级间无明显联系。
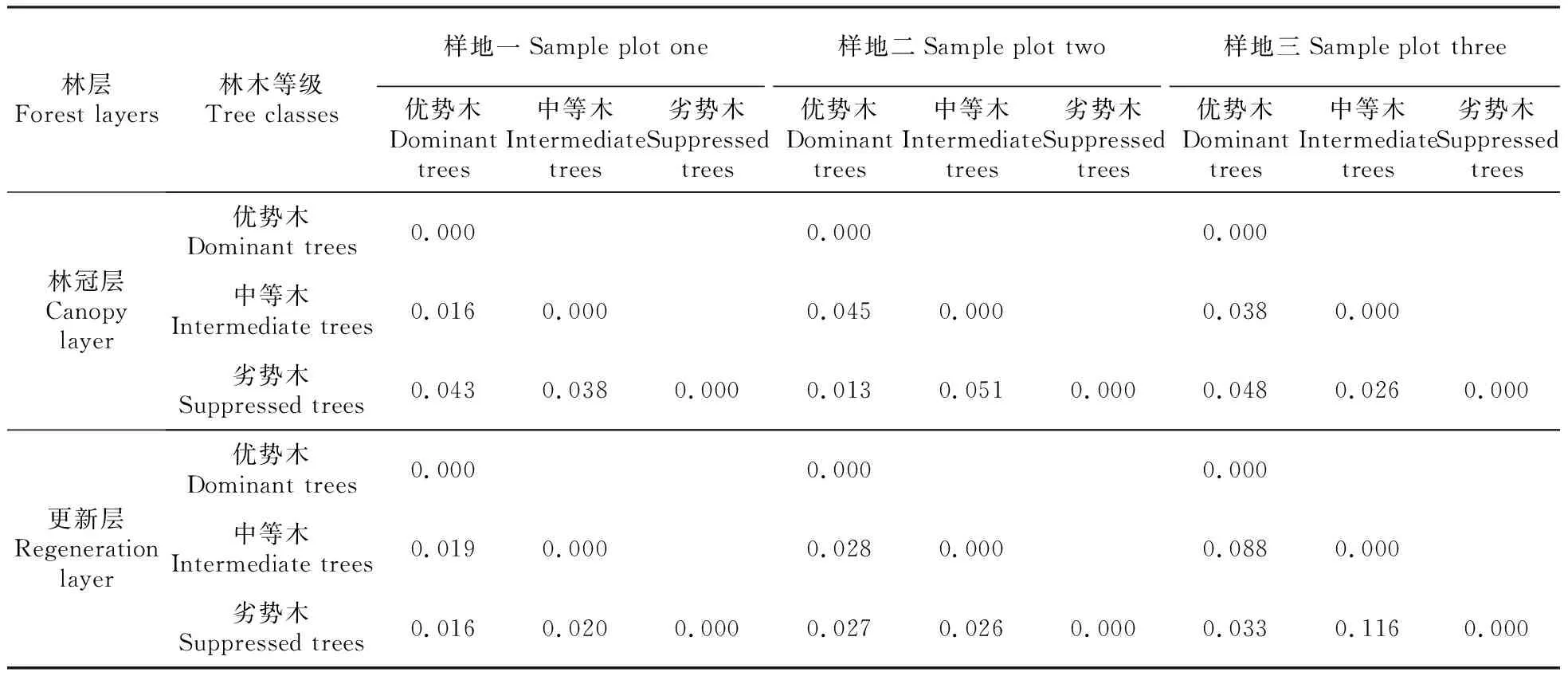
表4 不同优势等级云南松遗传距离分析
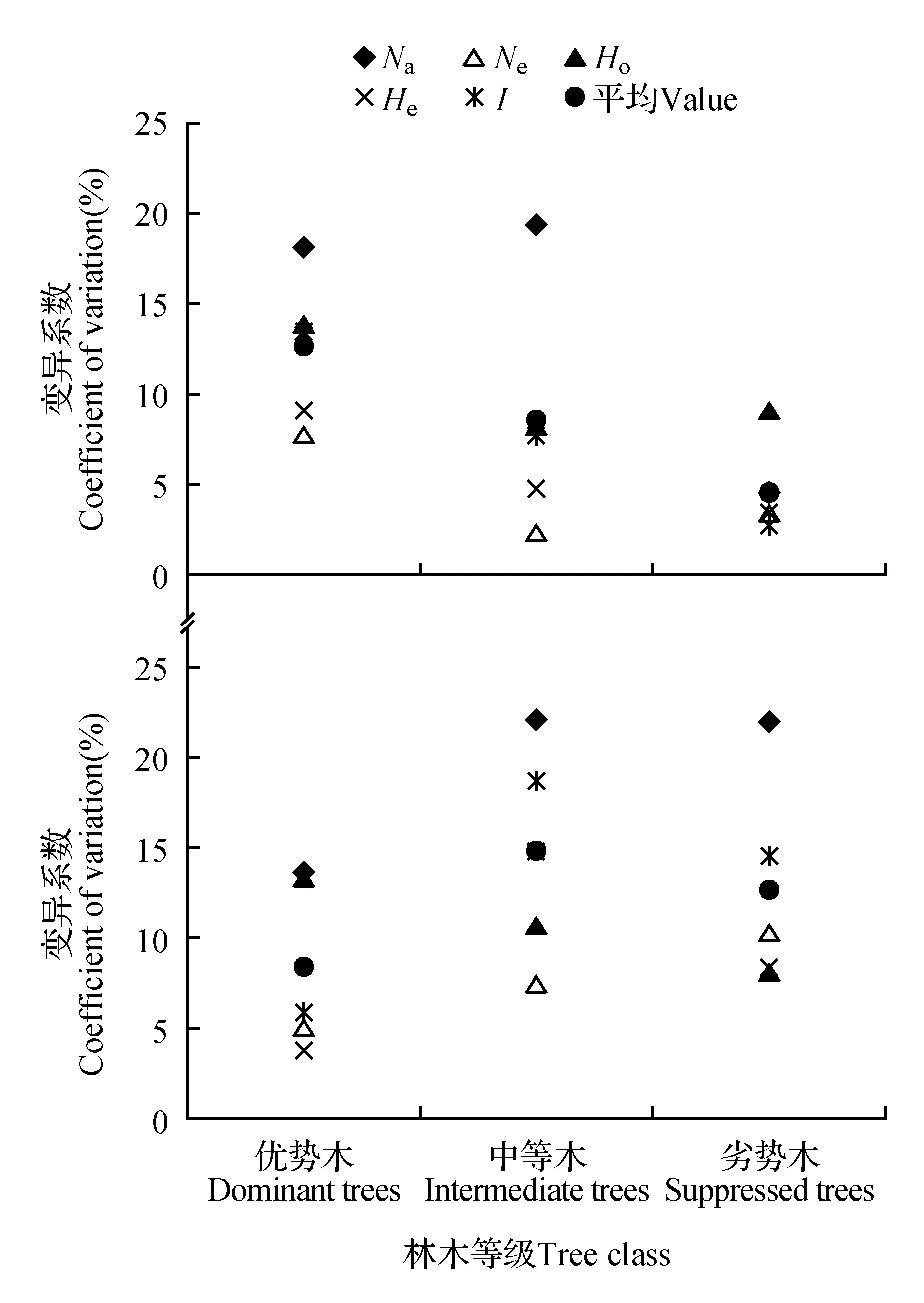
图2 不同优势等级云南松遗传多样性变异分析 A.林冠层;B.更新层 Na.等位基因数;Ne.有效等位基因数;Ho.观测杂合度;He.期望杂合度;I. Shannon’s信息指数Fig.2 Variance coefficient of genetic diversity in different dominance hierarchies of P.yunnanensisA.Canopy layer; B.Regeneration layer Na.Number of alleles; Ne.No. of effective alleles; Ho.Observed heterozygosity; He.Expected heterozygosity; I.Shannon’s information index
3 讨论
林木生长受遗传物质或环境的影响而呈现不一样的表型性状,出现不同的优势程度,然而,这些不同优势等级林木间的遗传分化和遗传多样性差异等均未明确。本研究对不同生长优势等级林木间的遗传多样性进行分析,结果表明,无论在林冠层还是更新层,不同生长优势等级云南松林木的遗传多样性均表现为观测杂合度低于期望杂合度,表现为杂合子缺失。与前期不同群体遗传多样性的研究相比[15],各遗传多样性指标有所降低,其中以观测杂合度差异稍大。在林冠层,三个样地优势木的有效等位基因数、Shannon’s信息指数、观测杂合度和期望杂合度分别为2.083、0.762、0.290和0.423,均表现为优势木略高于中等木和劣势木。在更新层,有效等位基因数、Shannon’s信息指数、观测杂合度和期望杂合度分别为2.063、0.774、0.272和0.410,除观测杂合度外,其余3个指标也均表现为优势木略高于中等木和劣势木。这在其它树种的研究中有类似的报道,如华北落叶松的遗传多样性比较中结果发现优树群体的杂合度或多样性水平要高于天然林,可能是杂种优势的存在使得基因杂合性高的个体更容易被选中成为优树[12]。从总体平均值来看,不同优势等级云南松林木间遗传多样性以优势木较高,但各等级间的差异不显著。
遗传距离及其聚类分析表明,不同优势等级林木间的聚类随机性较大,其聚类与林木生长优势等级间无必然联系,这也进一步表明遗传变异对林木生长优势等级差异无明显响应,即遗传多样性受林木生长分化影响不明显,这可能与林木的天然繁育系统相关。然而,其它的研究表明,不同生长优势等级间在臭氧吸收特征与环境调控[5]、树干液流差异及其对土壤水分与气象因子的响应[4,6~7]等方面存在差异,可能与光照、温度、光合有效辐射等有关[5]。在光竞争的林分中,优势度(相对树高)是最直接反映树木对自然资源利用能力的树形因子[30],水力限制作用尚不明显下,随树高增加,优势度增大,树木接受更多光照,导致蒸腾和液流速率提高[31];优势木具有较强的蒸腾能力,可能是其占据林内有利空间位置,更为充分的获取光辐射能量以驱动树干液流[4,7];优势木在林分中占据空间优势,容易从环境中获得较多的资源,保证了高大树木维持较高的液流通量和对环境干扰后的恢复能力[32];优势木的树干储水能力较强[33~34],其对环境因子的改变具有较强的缓冲能力[6]。也有研究表明,不同生长优势等级间木材材性或生物量差异不明显,木材材性相对量对林木分化不太敏感[32];生物量分配研究表明,除根茎生物量之外,不同林木分化等级的生物量组分(包括枝、叶、树干和根系)的相对分配比例无显著差异,并保持根冠比相对稳定,其相对分配格局较为保守[2]。不同的研究从各自角度提示了不同生长优势等级间的差异,总体表现为易受环境影响的指标其波动较大,而不易受环境影响的相对比较保守。
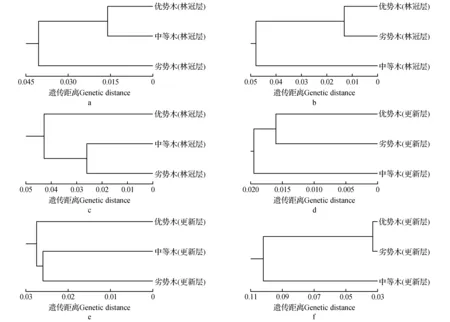
图3 不同优势等级云南松林木聚类图 a.样地一林冠层;b.样地二林冠层;c.样地三林冠层;d.样地一更新层;e.样地二更新层;f.样地三更新层Fig.3 UPGMA cluster in different dominance hierarchies of P.yunnanensis a.The forest canopy layer at the sample plot one; b.The forest canopy layer at the sample plot two; c.The forest canopy layer at the sample plot three; d.The regeneration layer at the sample plot one; e.The regeneration layer at the sample plot two; f.The regeneration layer at the sample plot three
本研究中,优势木的遗传多样性略高,但它们差异不显著,表明研究区域内云南松遗传多样性与不同生长优势等级林木间关系不明显。但是,调查中观察到,林分中作为优势木的,其长势或干型比中等木或劣势木好,同时对环境改变的缓冲能力也较强[6]。此外,遗传多样性高其适应环境的能力也越强,森林生态系统也更加稳固[12]。因此,在林分管理或抚育间伐中,建议保留优势木,给采伐迹地保留生长势好的植株,以其作为天然更新的母树,提高遗传增益的潜力很大,同时亦有望保持群体有较高遗传多样性和相对稳固的生态系统,但其在较长时间尺度上的表现仍有待进一步研究。
