梨F1代群体果实性状的遗传分析及相关性研究
2019-01-29赵亚楠欧春青张艳杰姜淑苓
赵亚楠,欧春青,王 斐,张艳杰,马 力,姜淑苓
(中国农业科学院 果树研究所/农业部园艺作物种质资源利用重点实验室,辽宁兴城 125100)
苹果梨是中国优良梨品种之一, 已有百余年的栽培历史。‘八月红’梨为中国近年来培育的优质红色梨新品种,均作为梨杂交育种的亲本被广泛利用。国内外学者在梨果实性状遗传规律方面已有一些研究[1-2]。但有关‘苹果梨’与 ‘八月红’梨杂交后代的遗传规律还未有文献报道。为了填补理论上对其杂种后代遗传规律研究的空白, 本试验对‘苹果梨’与‘八月红’杂种后代果实主要经济性状的遗传规律进行初步的观察研究, 以期为梨遗传育种和果实性状的QTL定位提供参考依据。
1 材料与方法
1.1 试验材料
试验材料为2005年春播种的以‘苹果梨’为母本,‘八月红’为父本的210株杂交后代结果实生苗。苗木定植于中国农业科学院果树所试验基地,行株距为3.0 m×1.0 m,常规管理。2010年部分单株开始结果,2015年开始调查。
1.2 测定项目与方法
杂种实生苗果实主要经济性状连续调查记载3 a。调查项目有:果实大小、果形指数、果心大小、果肉硬度、可溶性固形物质量分数和可滴定酸质量分数6个果实性状。所有指标都在果实采后进行测定。
单果质量用天平(精度0.01 g)测定,取平均值。果实的最大纵径和横径以及果心直径用游标卡尺测量,硬度用GY-1型果实硬度计测定,可溶性固形物质量分数利用手持折糖仪测定,可滴定酸质量分数用905全自动电位滴定仪(Metrohm)测定[3]。
1.3 数据处理
采用IBM SPSS Statistics 20对梨果实性状数据进行统计分析,用 Kolmogorov-Smirnov Test和正态Q-Q图对调查的各性状进行正态分布检验,绘制频率分布直方图。相应的遗传指标用Excel 2013进行统计分析。
2 结果与分析
2.1 果实性状遗传分析
2.1.1 果实大小 试验结果表明,‘苹果梨’与‘八月红’组合杂交后代的果实大小偏度值偏大,呈偏正态分布,属于微效多基因控制的数量性状遗传(表1和图1)。杂交后代的平均果质量为116.04 g,小于中亲值,甚至小于低亲,低于低亲值的后代占91.90%,表现出低低亲遗传,呈现向小变异的趋势,可能是亲本的非加性效应比较强。遗传传递力为54.91%,变异系数为46.03%,说明后代分离广泛,这是由于在有性杂交中双亲的非加性效应解体,导致后代呈回归变异。母本苹果梨果体大小差异悬殊,一般大果质量267.5 g,小果质量66.15 g,最大果可达600 g, 最小果仅23.2 g[4]。从果实质量变化差异悬殊来看,此性状在群体中遗传潜力大,选择空间大,即通过选择改良的效果好。
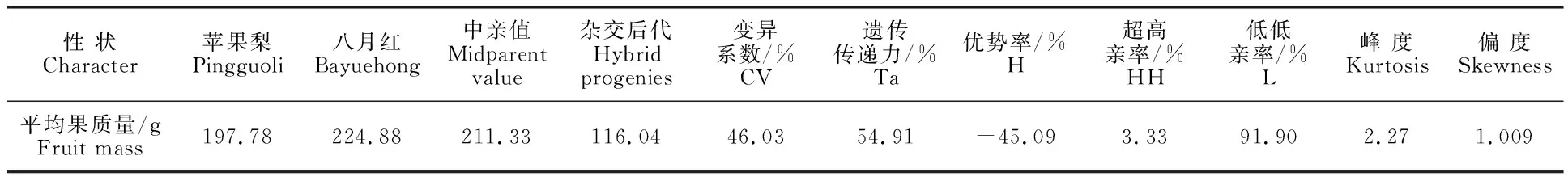
表1 ‘苹果梨’ב八月红’杂交后代果实大小的遗传变异Table 1 Genetic variation of fruit size in ‘Pingguoli’ × ‘Bayuehong’ hybrid progenies

图1 ‘苹果梨’ב八月红’杂交后代果实大小次数分布Fig.1 Frequency distribution of fruit size in ‘Pingguoli’ × ‘Bayuehong’ hybrid progenies
2.1.2 果形指数 从表2可以看出,杂交后代的果形指数平均值为0.93,与中亲值一致。变异系数为9.13%,遗传传递力为99.63%。超高亲率(5.71%)和低低亲率(1.43%)都较低,后代中有92.86%的果形指数介于双亲之间,总体表现出趋中遗传变异,呈较好正态分布,属于微效多基因控制的数量性状遗传(图2)。
2.1.3 果心大小 从表3可以看出,杂交后代的平均果心大小为38%,与中亲值相等。后代中有79.05%的果心大小介于双亲之间,超高亲率(6.19%)和低低亲率(14.76%)都较低。遗传传递力为96.45%,变异系数为17.36%,后代分离不广泛,总体表现为趋中偏小遗传变异,偏度值偏大,呈偏正态分布(图3)。
2.1.4 果肉硬度 试验结果表明,‘苹果梨’与‘八月红’组合杂交后代的果肉硬度为连续性分布,但不满足正态或偏正态分布,而是呈现主基因-多基因模型的多峰分布(表4和图4)。推测在该遗传群体中,果肉硬度是由主效基因和微效多基因共同控制的复杂遗传。
后代平均果肉硬度为7.12 kg/cm2,大于中亲值,甚至高于高亲值,出现较多的超高亲植株,平均超高亲率为59.0%,总体表现出超高亲遗传的倾向,说明此组合的高硬度性状遗传给后代的能力强。变异系数为27.68%,后代分离较广泛,极差值达9.32 kg/cm2。

表2 ‘苹果梨’ב八月红’杂交后代果形指数的遗传变异Table 2 Genetic variation of fruit shape index in ‘Pingguoli’ × ‘Bayuehong’ hybrid progenies

图2 ‘苹果梨’ב八月红’杂交后代果形指数次数分布Fig.2 Frequency distribution of fruit shape index in ‘Pingguoli’ × ‘Bayuehong’ hybrid progenies

表3 ‘苹果梨’ב八月红’杂交后代果心大小的遗传变异Table 3 Genetic variation of core size in ‘Pingguoli’ × ‘Bayuehong’ hybrid progenies
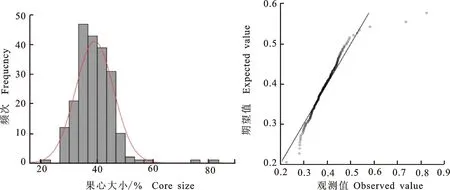
图3 ‘苹果梨’ב八月红’杂交后代果心大小次数分布Fig.3 Frequency distribution of core size in ‘Pingguoli’ × ‘Bayuehong’ hybrid progenies

表4 ‘苹果梨’ב八月红’杂交后代果肉硬度的遗传变异Table 4 Genetic variation of flesh hardness in ‘Pingguoli’ × ‘Bayuehong’ hybrid progenies
2.1.5 可溶性固形物质量分数 从表5可以看出,杂交后代的平均可溶性固形物质量分数(SSC)大于中亲值。遗传传递力强,达到105.98%。后代中超高亲率为44.00%,并且在后代中出现SSC达21.99%的高亲株,具有很强的杂种优势。后代分离广泛,极差值达14.59%,变异系数为14.53%,总体表现为超高亲遗传的倾向,呈较好的正态分布(图5)。
2.1.6 可滴定酸质量分数 从图6可以看出,‘苹果梨’和‘八月红’组合杂交后代的可滴定酸质量分数(TAC)分3个区段,呈不连续性分布。不满足正态或偏正态分布。但在3个不同区段内,株系间又表现出明显连续性变异和多峰分布,呈主基因-多基因模型。推测TAC这一性状可能是由多个主效基因和微效多基因共同控制的复杂遗传。从表6可以看出,杂交后代的TAC平均值大于中亲值,遗传传递力强(114.73%),出现较多的超高亲株(38.00%)。后代分离广泛,变异系数为58.41%,极差值高达1.099,选择空间大,总体表现出高亲遗传。
2.2 果实性状间的相关性分析
由表7可知,果实大小与果心大小、果肉硬度呈极显著负相关,而与果形指数、SSC和TAC相 关性均不显著;TAC与果形指数呈极显著正相关,与TAC呈显著正相关;其余性状间相关性不显著。在这些相关性较强的性状中,其QTL可能在连锁群所处的位置较近。

图4 ‘苹果梨’ב八月红’杂交后代果肉硬度次数分布Fig.4 Frequency distribution of flesh hardness in ‘Pingguoli’ × ‘Bayuehong’ hybrid progenies

表5 ‘苹果梨’ב八月红’杂交后代SSC的遗传变异Table 5 Genetic variation of SSC in ‘Pingguoli’ × ‘Bayuehong’ hybrid progenies
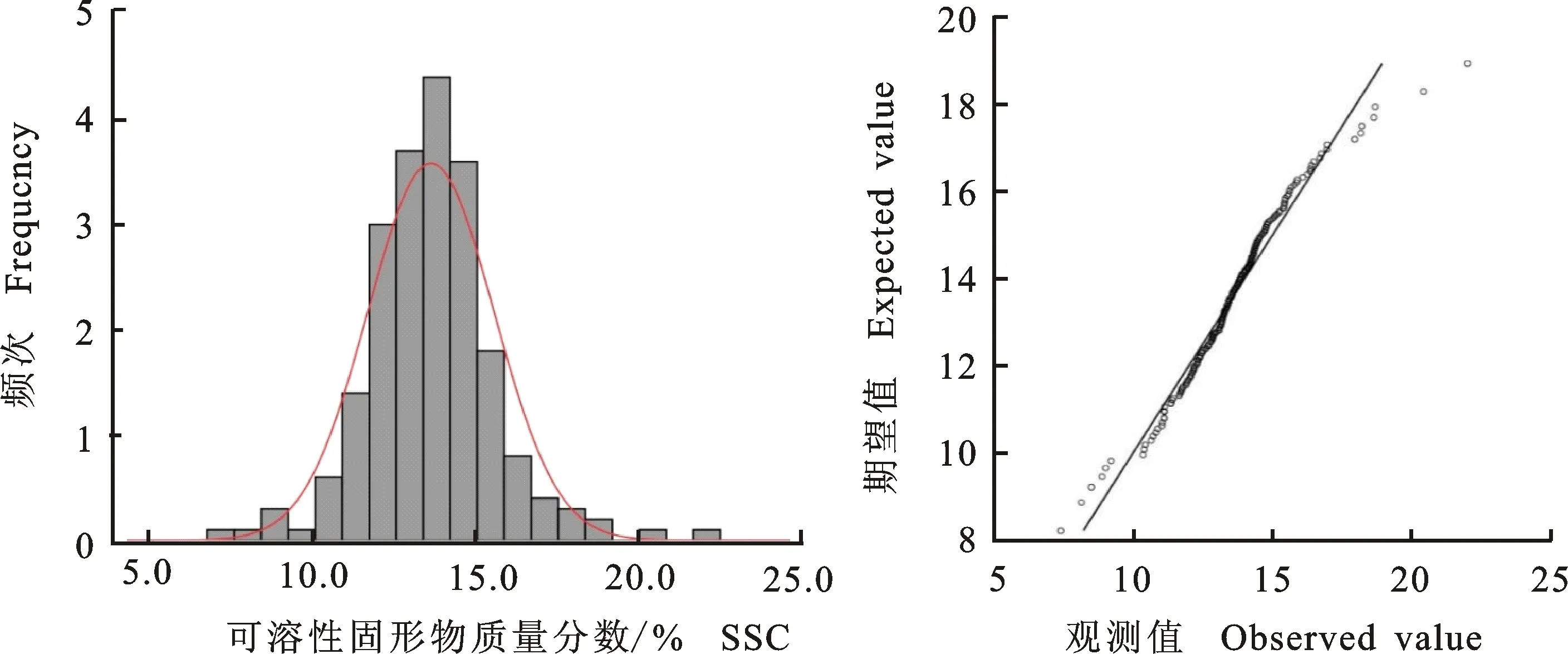
图5 ‘苹果梨’ב八月红’杂交后代SSC次数分布Fig.5 Frequency distribution of SSC in ‘Pingguoli’ × ‘Bayuehong’ hybrid progenies
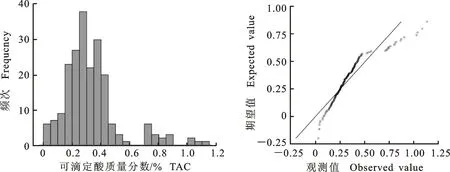
图6 ‘苹果梨’ב八月红’杂交后代TAC次数分布Fig.6 Frequency distribution of TAC in ‘Pingguoli’ × ‘Bayuehong’ hybrid progenies

表6 ‘苹果梨’ב八月红’杂交后代TAC的遗传变异Table 6 Genetic variation of TAC in ‘Pingguoli’ × ‘Bayuehong’ hybrid progenies
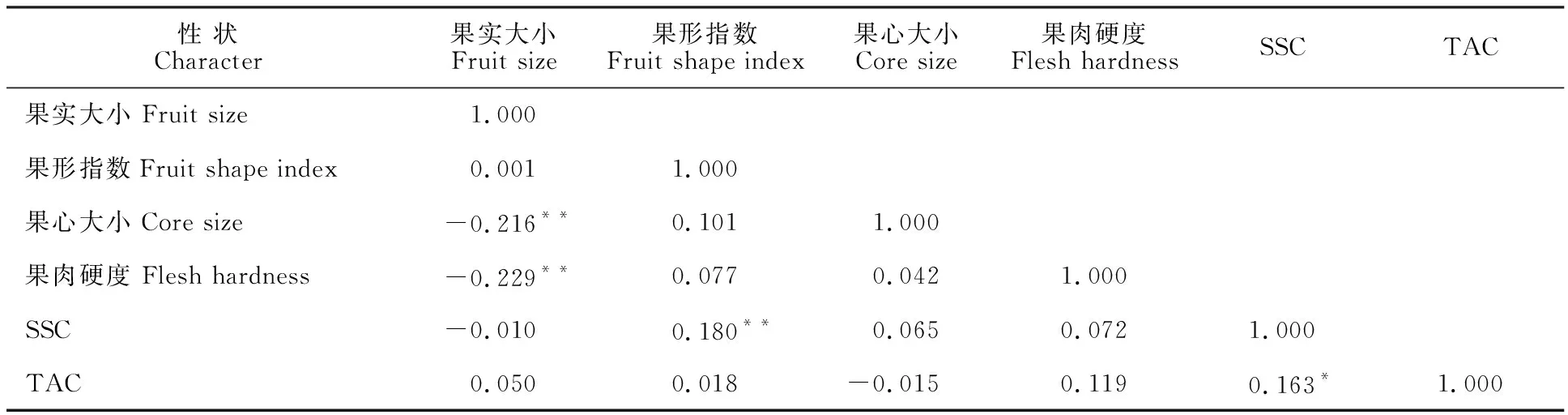
表7 ‘苹果梨’ב八月红’杂交后代果实性状的相关性分析Table 7 Correlation analysis on fruit traits of ‘Pingguoli’ × ‘Bayuehong’ hybrid progenies
注:**表示在 0.01 水平(双侧)上极显著相关;*表示在 0.05 水平(双侧)上显著相关。
Note:**presents significant correlation at 0.01 level;* presents correlation at 0.05 level.
2.3 果实性状间的通径分析
相关分析表明,果实性状间存在一定的相关性,要进一步研究性状间的数量关系,需要进行通径分析。以果实大小为因变量,另外5个性状为自变量,进行逐步回归分析,建立果实大小的最优回归方程为:Y=222.712-5.975X1-162.619X2+ε(X1:果肉硬度,X2:果心大小,R=0.309,F=10.888**)。同理,以SSC为因变量,建立最优回归方程为:Y=9.254+4.076X3+1.608X4+ε(X3:果形指数,X4:TAC,R=0.241,F=6.376**)(结果与表7一致)。经通径分析可知,果肉硬度和果心大小对果实大小的直接通径系数分别为-0.221和-0.206,表明果肉硬度对果实大小直接作用最大,决定系数(R2)=0.095,剩余通径系数(Pe)=0.951,该值较大,说明还有对果实大小影响较大的因素没有考虑到。果形指数和TAC对SSC的直接通径系数分别为0.177和0.160,表明果形指数对SSC直接作用最大,R2=0.058,Pe=0.970,同样也有对SSC影响较大的因素没有考虑到。
3 讨论与结论
梨不同种或品种间进行有性杂交过程中,受双亲基因间的加性效应以及非加性效应的影响,后代性状往往出现广泛分离的现象,形成不同的类型[5]。其中大多数是劣质性状变异,但也会出现将双亲中控制同一性状的不同微效基因积累起来,出现超高亲的类型[6]。正是因为基因重组可以将双亲控制不同性状的优良基因结为一体,杂交育种才更有意义。本研究结果表明,‘苹果梨’与‘八月红’组合杂交后代群体中多个果实性状存在不同的遗传变异倾向和数量性状混合遗传模型。
果实性状遗传规律分析表明,果实大小、果心大小和果形指数这3个性状的后代果实平均值都小于或等于亲中值,且遗传传递力都低于100%,优势率等于0或为负值,杂种优势不明显。果实大小为低低亲遗传变异,普遍出现经济性状退化现象,存在较明显的非加性效应解体的影响,与前人的研究结论一致[7-9]。果形指数为趋中偏大遗传变异,与方成泉等[10]趋中偏小遗传的结论有些许差异,这可能与本组合的双亲果形指数相差较大、遗传传递力高及受亲本影响大而趋向高亲本遗传有关[11]。果心大小与果形指数类似,但总体表现为趋中偏小遗传,非加性效应较弱,这与滕美贞[11]的结论一致,但与大多数研究者认为的梨后代果心大小趋中偏大的趋势有差别[12],这可能与苹果梨小果心遗传传递力较强有关[13-14]。
果肉硬度、SSC和TAC的后代果实平均值都大于亲中值,遗传传递力都超过100%,优势率都为正值,具有很强的杂种优势。说明这3个性状遗传以加性效应为主,并且存在着正向的非加性效应。果肉硬度表现为超高亲遗传倾向,有明显增大的趋势,表明后代的耐贮性有增强的趋势,对梨果实的储存有重要意义[15]。这一结果与崔艳波等[16]的研究结果一致,在草莓[17]和番茄[18]杂交后代中发现果肉硬度也有此规律,但在桃[19]杂交后代中却出现相反结果,因此不同梨品种亲本杂交后代是否都符合此规律,还有待进一步研究。TAC为趋向高亲遗传,变异系数最大(58.41%),呈现较高的变异程度,遗传潜力最大,选择改良效果最好,在苹果中也有类似报道[20-21]。SSC为超高亲遗传,与前人研究结论相同[22-23],这可能与后代果质量普遍偏小、硬度普遍偏高及肉质紧密有关[24]。
果实大小、果形指数、果心大小和SSC这4个性状在杂交后代中均呈正态或偏正态分布分布,为微效多基因控制的数量性状。该结果与崔艳波等[16]结果一致。其余2个性状次数分布图呈多峰分布,呈现主基因-多基因模型,可能是由多个主效基因和微效多基因共同控制的复杂遗传,这一结果与番茄[18]和西瓜[25]中的研究结果类似。
经相关分析和通径分析,梨果实大小与果心大小、果肉硬度呈极显著负相关,且果肉硬度对果实大小直接作用最大,最优回归方程为Y1=222.712-5.975X1-162.619X2+ε(Y1为果实大小,X1为果肉硬度,X2为果心大小)。SSC与果形指数呈极显著正相关,与TAC呈显著正相关,最优回归方程为Y2=9.254+4.076X3+1.608X4+ε(Y2为SSC,X3为果形指数,X4为TAC)。
本研究梨F1代群体果实性状中,变异系数大小顺序为TAC>果实大小>果心大小>果肉硬度>果形指数>SSC,变异系数越大,说明性状在群体中的遗传潜能和选择空间越大,通过选择改良的效果就越好;遗传传递力大小顺序为果肉硬度>TAC>SSC>果形指数>果心大小>果实大小,遗传传递力越大,杂种优势越明显。在今后的杂交育种中,可根据育种目标和上述规律合理配置亲本组合,以增加优株入选率。
