仿生无机纳米材料改造生物体的研究进展
2019-01-14唐睿康马为民邹志刚
熊 威 唐睿康 马为民 邹志刚*,,4
(1南京大学物理学院,固体微结构物理国家重点实验室,人工微结构科学与技术协同创新中心,环境材料与再生能源研究中心,南京 210093)
(2浙江大学化学系,生物物质与信息调控研究中心,杭州 310027)
(3上海师范大学生命科学学院,上海 200234)
(4南京大学物理学院,江苏省纳米技术重点实验室,南京 210093)
0 引 言
在地球形成之后的数十亿年间,地球上的生命形式一直是原始和低等的,并且主要是以古细菌和原始藻类为主[1]。直到距今5.41亿至5.12亿年前的寒武纪早期,在2 000多万年的时间内地球上突然涌现出各种各样的动物,它们不约而同地迅速起源和出现[2-4]。海绵、腔肠、腕足、节肢、脊索动物等一系列与现代动物形态基本相同的动物在地球上 “集体亮相”,形成了多种门类动物同时存在的繁荣景象[3-4]。这就是地质学上所谓的 “寒武纪生命大爆发”。在这个生物大爆发时期,地球上的生命形式出现了质的变化,最具标志性的是以三叶虫为代表的甲壳类生物的出现,并且此后出现的更加高等的动物都或多或少具有硬组织结构[4-6]。说明在这个时期,生物体开始学会了利用材料来改造自身去适应环境,而生物体在与环境相互作用中形成自身需要的材料的过程实际上就是生物矿化的过程,因此生物矿化对生物进化和寒武纪生物大爆发起到了至关重要的作用。生物体可以通过生物矿化形成具有复杂精细结构并且生物性能优异的有机-无机复合材料,比如骨骼、牙齿和贝壳等[7-8]。这些生物材料从纳米尺度到宏观尺度都具有高度有序的层次结构,而且它们特殊的结构和性质,可以为生物体提供机械支撑、保护、运动、信号传感等功能[9-14]。例如,硅藻可以利用环境中的硅酸盐在细胞表面形成一层结构精致的二氧化硅外壳,并且这层二氧化硅外壳可以为它们提供机械保护、光子晶体和pH缓冲剂的作用[15-17]。还有我们生活中常见的鸡蛋,也是一个很典型的例子,鸡蛋壳不仅可以为里面的细胞提供机械保护,还能在保持内部细胞活性的同时防止外部细菌的入侵[18-19]。此外,蛋壳还能为内部胚胎的生长提供足够的钙源[20-21]。除了保护作用,还有一些细菌可以利用内部的磁铁矿晶体(Fe3O4或Fe3S4)作为磁传感器[11,22]。
自然界中的生物矿化现象,是生物体通过生物大分子的调控形成无机矿物的过程,其化学本质是有机分子与无机材料的界面相互作用。生物矿物根据其成分主要可分为两大类:钙化的生物矿物和非钙化的生物矿物[23]。在已发现的生物矿物中,钙化的生物矿物约占总数的一半,其中碳酸钙矿物最为广泛,磷酸钙矿物次之[7,10]。碳酸钙矿物主要出现在无脊椎动物中,比如软体动物的外壳和海胆的刺[7,10]。磷酸钙矿物则主要出现在有脊椎动物中,最有代表性的就是骨骼和牙齿中的主要成分羟基磷灰石[7,10]。非钙化的生物矿物种类也很多,最主要的是二氧化硅类矿物,主要存在于硅藻、海绵骨针和一些植物中[7],其次是在趋磁细菌中发现的铁的硫化物和氧化物[7,24]。
虽然生物矿化是自然界中广泛存在的现象,但是还有很多低等生物体不具备生物矿化的功能,而它们往往具有矿化生物体不具有的特殊功能,因此对于这些生物体的改造是非常有意义的。受到自然界生物矿化现象的启发,越来越多的科学家试图通过仿生矿化的策略为生物体合成与制备具有特定结构和特殊功能的纳米材料,从而达到改造生物体的目的。在过去的十年里,基于生物-材料界面复合技术的生物体改造研究已经逐渐形成一个研究领域,并且引起了来自化学、物理学、材料科学、生物学乃至生物医学等诸多领域的科学家们的强烈好奇和广泛兴趣。对于通过仿生矿化改造生物体以及基于单细胞壳化的功能化改造方面的研究,国内外已有多篇综述[19,25-32]。而近年来,通过材料改造生物体的研究已经从最初的单细胞矿化,发展到多细胞聚集,利用的材料体系从最初的生物矿物发展到其他无机纳米功能材料,改造的生物体从最初的大肠杆菌和酵母细胞拓展到病毒、藻类和动物细胞等,赋予的功能也从最初的生物保护延伸到能源、环境、医学、催化等诸多方面。因此,本文对近年来通过材料手段改造生物体的进展做了一个系统的分类和梳理,并对当前的问题和未来的挑战做了总结和分析。
1 材料在生物界面形成的调控机制
生物矿物的形成过程,比如晶体的成核、生长、形貌、同质异形体类型和组分,通常都受到生物体严格的控制,因此生物矿物在形貌、结晶行为和材料性质上与地质学上对应的矿物有着显著的不同。虽然理解每种矿物特有的形成机制并非易事,但是似乎存在一些对很多生物体都具有普适性的生物矿化调控策略。Mann把这些策略主要归纳为5种[33]:化学控制,空间控制,结构控制,形貌控制和构筑控制。
生物矿化区别于普通矿化最核心的特点就在于有机基质在矿化过程中的调控起着至关重要的作用。因此,有机基质的调控作用在仿生矿化中具有核心地位。仿生矿化中常见的有机添加剂类型主要有聚合物、表面活性剂和低分子量小分子和生物大分子。其中,一些生物大分子对于特定生物材料有特异性的识别能力,并且自身具有较强的可操作性,可以被用来制备具有多级结构的理想模板和用于控制功能复合材料构建。已有研究表明,调控钙矿化的生物大分子通常都含有羧基(-COO-)[34-39],而调控硅矿化的生物大分子通常带有-NH2或-NH-[40-44]。比如通过富含谷氨酸(Glu)、天冬氨酸(Asp)和磷酸化丝氨酸(pSer)的蛋白可以控制磷酸钙生物医学材料的制备[45-48],而通过富含精氨酸(Arg)、赖氨酸(Lys)或组氨酸(His)的阳性多肽和多胺(polyamine)类似物可以调控二氧化硅硅材料的合成[49]。
目前,仿生矿化的调控机制主要有:模板作用、限域的反应环境、非经典结晶、电/磁场诱导结晶[50]。在生物矿化过程中,有机基质的模板作用是有机分子与无机矿物最主要的一种相互作用。有机分子通过有机-无机界面的分子识别在晶体成核、生长及微结构的有序组装等方面起到的调控作用在仿生矿化中具有核心地位[45]。仿生矿化中的有机模板主要有2种类型[51-52]:一种是人工合成模板,例如Langmuir-Blodgett(LB)膜、自组装单层膜、囊泡、微囊、微乳液、胶束、反相胶束、单层分子膜;另一种是生物模板,主要是生物大分子或生物中的有机质,如胶原蛋白、磷脂、脂质体、甲壳素、胆固醇等。在限域的反应环境中的矿化主要涉及到2种策略[50,53]:一种是通过模板的途径构建限域的有机支架,另一种是利用限域的反应环境,比如纳米反应器。另外,中空的蛋白质结构也可以作为限域的反应环境用来合成各种纳米颗粒[54-60]。除了模板和限域反应环境中的经典结晶途径之外,仿生矿化还能够利用非经典的颗粒诱导的结晶途径[61]。非经典的结晶过程涉及到预先形成的纳米颗粒到一个有序的超级结构的自组装过程,并且在这个过程之后融合成单晶。电/磁场诱导结晶是指利用电场对带电离子的作用或磁场对磁性纳米颗粒的作用,诱导它们在结晶过程中沿着电场线或磁场线的方向定向排列组装[62-66]。电场主要影响结晶过程,而磁场还能够影响纳米颗粒的组装过程,比如控制纳米材料的一维、二维甚至三维的组装过程[63-66]。
2 纳米材料改造生物体的方法
经过长期的自然选择和生物进化,自然界中的大部分生物体学会了利用材料来改造自己,但是很多简单的生命形式,例如绝大多数的微生物,还不具备利用材料改造自身功能的能力,因而它们更容易受到环境胁迫的干扰。受到自然界生物矿化现象的启发,越来越多的化学家和材料学家们选择利用纳米材料对这些生物体进行改造[28,31-32]。纳米材料改造生物体的关键在于材料与生物的结合,核心是生物与材料界面的相互作用对生物体功能的影响。经过近十年的研究和探索,已经发展出了一系列实现材料与生物体结合的方法,但是目前实现纳米材料与生物体的结合,主要还是基于仿生矿化的策略。
2.1 表面矿化基团改性诱导矿化
2.1.1 层层自组装修饰
层层自组装(layer by layer assembly,LbL)是一种通过交替沉积相反带电荷的聚电解质分子的表面修饰方法。通过层层自组装,也能够赋予表面特殊的活性基团。这种方法最初利用带电基板在带相反电荷的聚电解质溶液中交替沉积制备聚电解质自组装多层膜[67]。由于绝大多数的细胞,表面不具备诱导矿化的活性位点,所以通常需要对细胞进行表面改性,通过层层自组装可以改变细胞的表面电荷及赋予细胞表面特定的矿化位点,从而实现细胞表面的矿化,并且通过调节聚电解质的吸附时间和层数,可以调控矿化外壳的厚度。对细胞的表面矿化改造,最早是在酵母细胞上实现了磷酸钙矿化改造[68],这也是仿生矿化改造生物体的第1个尝试,并且随后很多工作都是以酵母细胞为模型细胞开展的。酵母是真核细胞的典型代表,常作为研究的模式生物[69-71]。酵母的细胞壁主要由弱电负性的多聚糖、甘露糖、N-乙酰氨基葡萄糖等组成,缺乏诱导矿化相关的残基和官能团,因此需要对酵母细胞进行表面修饰[72]。聚二甲基己二烯氯化铵(PDADMAC)是一种富含季胺基并且带正电的聚电解质,它们可以吸附在酵母细胞表面,当细胞表面带有一定量的负电荷后,再吸附富含羧基(-COOH)且带负电的聚丙烯酸(PAA)分子,经过多次交替吸附之后,细胞表面的活性位点就增多了(图1)。已有的研究已经证明,羧基是诱导钙矿化的活性位点[34-39],当细胞表面富含羧基时,可以富集钙离子,从而诱导表面钙矿化的发生(图1),最终形成一层纳米尺度的磷酸钙外壳[68]。
利用层层自组装技术不仅能够赋予细胞表面钙矿化的位点,通过改变最外层的聚电解质,也能将其他的纳米材料引入细胞的表面。比如将富含季胺基的PDADMAC暴露在层层自组装的最外层,就可以诱导细胞表面的硅矿化发生或者吸附纳米二氧化硅颗粒,从而在细胞表面形成纳米级的二氧化硅外壳[73]。利用类似的方法,选用具有活性位点的多肽替代聚电解质,可以实现对小球藻细胞的纳米二氧化钛包裹[74]。此外,直接利用聚电解质和无机 纳米材料或者电荷改性的纳米材料进行层层自组装[75-76],也能实现对细胞与纳米材料的结合。
目前,通过层层自组装对细胞进行表面改性并实现纳米材料在表面的复合是最为常用的细胞功能化改造方法之一,但是这种方法通常需要多次的离心操作,容易对细胞造成机械损伤,并且细胞反复浸在聚电解质溶液中,也会受到聚电解质的毒性影响。因此这种方法对于细胞壁厚、带电性强的细胞有比较好的适用性,但是对于无细胞壁或者带电性弱的细胞就不具有很好的适用性,尤其是对无细胞壁的细胞,离心操作会破坏细胞膜结构,从而使改造失去意义。

图1 酵母细胞仿生钙矿化示意图[68]Fig.1 Scheme for biomimetic calcification of yeast cell[68]
2.1.2 直接吸附修饰
对于一些没有细胞壁结构的细胞,比如哺乳动物细胞,它们的表面功能化改造需要更加温和的手段和细胞相容性更好的材料,所以一直是细胞功能化改造的难题。2014年,韩国的Choi课题组报道了一种单层聚电解质修饰哺乳动物细胞表面诱导硅矿化的方法[77]。他们将HeLa细胞浸在阳离子聚电解质聚乙烯亚胺(PEI)溶液中,让细胞表面吸附上一层PEI分子,目的是表面胺基化,然后在正硅酸甲酯(TMOS)的溶液中成功诱导细胞表面的硅矿化(图2a)将二氧化硅引入到了哺乳动物细胞表面,得益于PEI分子的高正电荷密度和胺基化程度,PEI能够很好地吸附于HeLa细胞表面并诱导硅矿化的发生,但正因如此,PEI对细胞的毒性也很大,因此还需要寻找细胞相容性更好的聚电解质或其他修饰分子才能使这种方法更加具有应用前景。随后,Choi课题组又合成了具有硅矿化基团的多肽链[78],通过吸附于酵母细胞表面,赋予其硅矿化的活性位点,实现了硅矿化改造(图2b)。但是这个改造过程中,需要将表面改性后的酵母细胞置于TEOS溶液中搅拌6 h,这说明人工合成的多肽链与细胞的吸附结合能力不强,表面矿化位点不多,并且长时间置于TEOS溶液中也会对细胞活性有影响。2016年,赵瑞波等[79]利用HeLa细胞表面的叶酸受体吸附叶酸分子,再通过叶酸分子对钙离子的螯合作用,实现了肿瘤细胞的靶向磷酸钙矿化。此外,苏宝连课题组也利用L-半胱氨酸结合的金纳米颗粒为酵母细胞构筑了一层可自修复的纳米金外壳[80]。还有一些研究者基于分子识别原理,通过特异性配体改性纳米颗粒,实现与细胞表面的结合[81]。
受到贻贝中的粘合蛋白启发,多巴胺已经成为界面修饰的可靠媒介[82]。在酵母细胞表面实现多巴胺的聚合是Choi课题组开发的另一种细胞表面改性方法[83],如果在聚多巴胺外壳上接枝PEI分子,可继续诱导硅矿化在表面发生[84]。随后,王本等[85]将多巴胺表面聚合成功应用到红细胞改造中。但是由于多巴胺在发生聚合时,会发生氧化反应对细胞产生毒性,并且聚合后的多巴胺呈现黑色,因此这个过程对细胞的损伤比表面吸附聚电解质可能更大,所以这种方法的应用有待继续探究。

图2 (a)PEI分子改性诱导HeLa细胞表硅矿化[77];(b)R4C12R4短肽改性诱导酵母细胞表面硅矿化[78]Fig.2 (a)Silicification on HeLa cell surface induced by PEImodification[77];(b)Silicification on yeast cell surface induced by R4C12R4 modification[78]
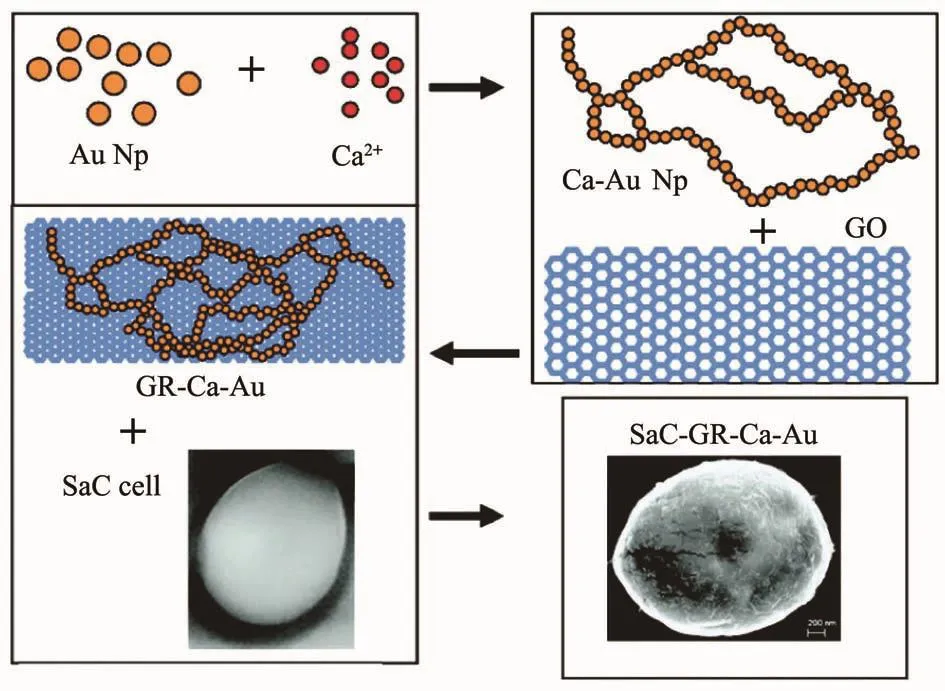
图3 钙离子功能化的纳米金颗粒辅助石墨烯片与酵母细胞结合[80]Fig.3 Graphene sheets are interfaced with yeast cells,using calcium ion functionalized Au nanoparticles[89]
2.2 金属离子表面富集诱导纳米材料沉积
在一些特殊的生物-材料复合体系中,生物体表面通过富集特殊的金属阳离子也可以诱导纳米材料在其表面形成。很多细胞可以通过细胞膜上的离子受体从环境中吸收过量的钙离子到细胞内部,因此这些钙离子的受体可以视为钙矿化的成核位点和离子结合位点[86-87]。由于钙离子正二价的特性,钙离子可以在2个带负电的分子间起到 “桥联”的作用。利用钙离子的“桥联”作用,柠檬酸盐改性的金纳米颗粒与氯化钙溶液共混可以引发金纳米颗粒在酵母细胞壁外表面形成一层金纳米壳[88]。根据这个策略,将酵母细胞浸入还原石墨烯和钙离子结合的金纳米颗粒混合溶液中,可以赋予酵母细胞一层还原石墨烯材料的外壳(图3)[89]。此外,铬离子在热醋穆尔氏菌细胞表面富集,在光照和半胱氨酸的存在下,可以诱发硫化镉纳米颗粒在细胞表面形成[90]。此外,一些表面带负电的病毒纳米颗粒表面,也能够直接吸附阳离子,使局部过饱和度增加,从而引发表面纳米材料沉积[91]。例如,日本脑炎疫苗在富含钙离子的模拟体液中表面可以富集大量的钙离子从而引发原位的钙矿化,形成一层磷酸钙纳米外壳[92]。与前面提到的层层自组装改性诱导纳米材料沉积不同,这种方法操作步骤更加简单,对生物体的机械损伤更小,但是这种方法通常需要生物体具有相对高的比表面积和表面电荷密度,适用性不如层层自组装改性的方法。
2.3 基因改造
自然界中的生物矿化现象都是受到基因调控的,是生物体可遗传的自发行为。但是目前人工对生物体的纳米材料改造都是不可遗传的,而且由于是非自发的行为,所以对生物体本身是有损伤的。因此,将与矿化蛋白相关的基因导入到生物体中,让其表达并进行自发的矿化是一种更为理想的策略。
科学家们已经对一些生物体做了矿化基因改造的尝试,例如在毕赤酵母(pichiapastoris)中过表达溶菌素酶和在大肠杆菌中过表达硅矿化蛋白都可以诱导硅矿化在表面发生[93-94]。与细胞的基因工程改造相比,病毒的基因工程改造相对更加容易。Belcher课题组长期利用基因改造的M13噬菌体诱导无机纳米材料的生长,并且得到了许多形貌结构特殊的纳米材料[95-98]。王广川等[99]在多年病毒矿化的研究基础上,通过对减活的人类肠道病毒71型(EV71)的 cDNA 进行基因改造(图 4a),获得了可以表达成核多肽诱导磷酸钙矿化的病毒(图4b),并且这种基因改造对病毒的活性影响很小,而矿化后的病毒却有了更好的热稳定性。通过基因改造赋予生物体矿化能力,相比于单纯通过化学与材料手段的改造,最大的优势是这种改造是可遗传的,但是对于其他功能的影响也是不可逆的。目前基因工程改造生物体的矿化功能,只在病毒、大肠杆菌和酵母等一些模式生物中实现,更加广泛的应用还有待于基因功能的研究及基因工程和材料技术的发展。
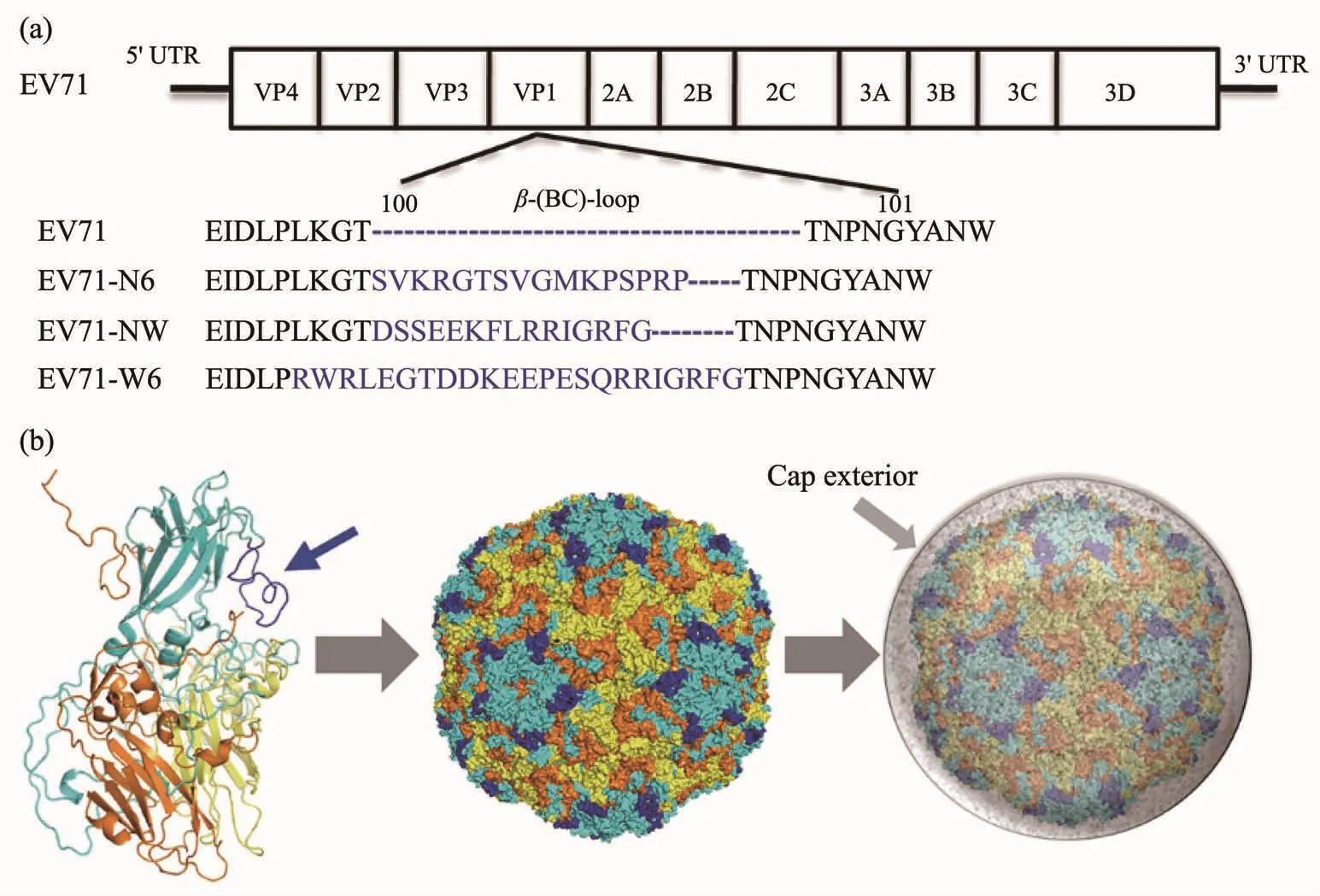
图4 基因改造诱导病毒自矿化[99]:(a)EV71基因组及在EV71基因组中的插入位点(VP1 蛋白上 β-(B-C)-loop);(b)改造后的病毒原位钙矿化Fig.4 Genetic modification-induced virus self-biomineralization[99]:(a)EV71 genome and the insertion site of the β-(B-C)-loop of VP1;(b)Gene engineered virus calcium mineralization in situ
2.4 矿化诱导聚集
在生物进化的过程中,单细胞到多细胞的转变是一个十分重要的变化,是生命由低等向高等进化的第一步。自然界中许多高等的生命体,都是以多细胞形式存在的,并且存在细胞的功能分化,而这种现象是单细胞生物所不具有的。在从单细胞到多细胞的转变过程中,首先需要细胞在空间上相互接近,形成细胞团簇,最后发生聚集,细胞空间位置和微环境的差别诱导细胞发生功能差异[100-102]。受到生物进化的启发,如果可以实现人工诱导和调控细胞的聚集,那么就有望发现生物体的新功能。目前,利用阳离子聚电解质PDADMAC的“桥联”作用,在诱导仿生硅矿化的同时,可以实现小球藻细胞的聚集(图5),并且发现聚集的小球藻在有氧条件下可以持续产生氢气[103]。不仅如此,直接使用二氧化硅纳米颗粒,在PDADMAC的“桥联”作用下,也能诱导蓝藻细胞的聚集[104]。2种方式都是利用仿生硅矿化的策略,各有优缺点,前者可以获得更加稳定的聚集体结构,但操作更加复杂,不适合大规模应用,后者操作简便有大规模应用的前景,但是获得的聚集体结构不如前者紧致和稳定。最近,通过仿生硅矿化诱导细胞聚集改性的策略也应用到了基因改造后的大肠杆菌[105],说明这种方法具有一定的普适性,可以拓展到其他生物体的改造中。如果能解决生物相容性和聚集体结构可控性的问题,将是一种非常有前景的生物改造策略。

图5 仿生硅矿化诱导小球藻细胞聚集[103]Fig.5 Cell aggregation of Chlorella induced by biomimetic silicification[103]
2.5 其他方法
以上介绍的方法都是基于生物体表面的改性实现的纳米材料复合,实际上在一些特殊的生物-材料体系中,一些生物体也可以直接与材料结合。比如,早在1989年,就有科学家利用溶胶-凝胶法实现了二氧化硅对酵母的包埋[106],随后这种方法也应用到了细菌、蓝藻等生物体系[107-109],但是这种方法对细胞的改造很难做到单细胞化或者纳米化,容易使很多细胞被包埋在成块的材料中,同时也会释放出醇类物质毒害细胞。王广川等[110]发现EV71病毒纳米颗粒可以在硅酸中实现原位硅矿化(图6),为病毒的矿化改造提供了一种新的便捷途径。此外近期的研究还发现,小球藻细胞可以自发地吸附带正电的二氧化铈纳米颗粒,形成外壳包裹在细胞表面[111]。这些生物-材料直接复合的方法,简便快捷,有利于大规模应用,但是需要生物界面与材料特性的良好匹配。例如,大规模的大肠杆菌硅矿化包裹,可以通过喷雾干燥技术来实现,并且包裹后的细胞活性得以保持,是一种有前景的大规模实现生物体包裹的技术[112]。

图6 EV71病毒纳米颗粒原位硅矿化[110]Fig.6 In situ silicification of EV71 virus nanoparticle[110]
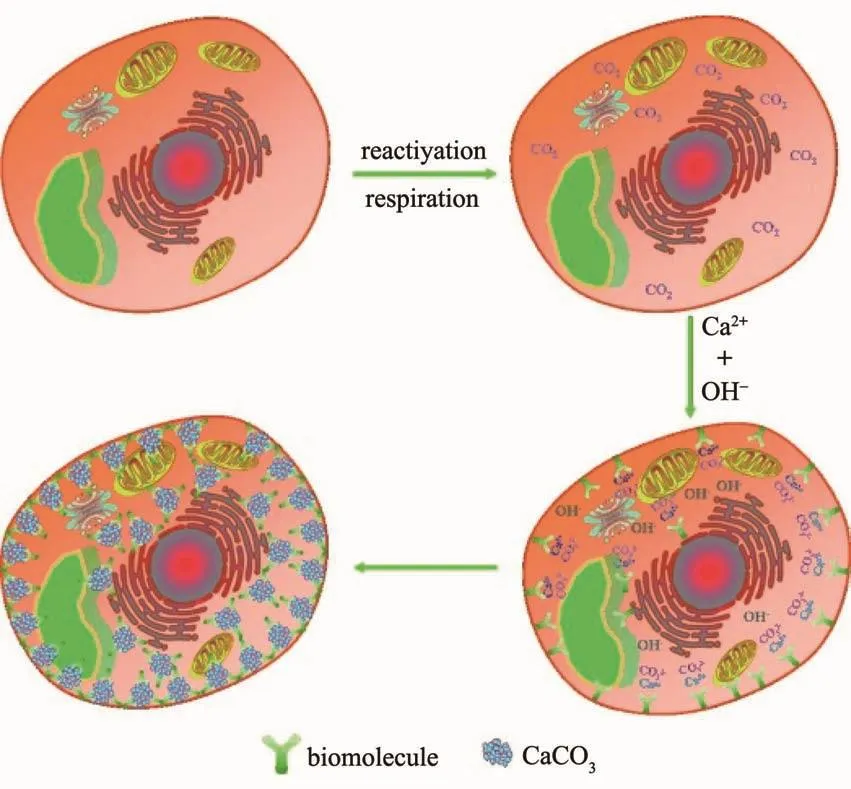
图7 酵母细胞内部碳酸钙纳米壳形成机制[113]Fig.7 Formation mechanism of CaCO3 nanoshell in yeast cell[113]
在自然界中,趋磁细菌能够在其细胞内部合成由磁铁矿(Fe3O4或 Fe3S4)组成的“指南针”,为它们定向运动提供保障[1,22]。这些在内部通过矿化形成的具有特殊结构的纳米生物材料对生物体的功能也起到十分重要的作用。受到自然界生物体内部矿化的启发,研究者也开始思考如何能够在生物体内部合成纳米材料从而对生物体实现功能化改造。例如,杨林课题组[113]通过用麦芽糖促进酵母的呼吸作用,在饱和氢氧化钙溶液中,利用酵母呼吸作用放出的二氧化碳与氢氧化钙反应,最终在酵母细胞内部形成一层5 nm的碳酸钙矿化层(图7)。随后,又有一些研究报道,在其他微生物细胞内部合成了不同形貌结构和组成的金属纳米颗粒,大致机制是利用微生物细胞内部特有的还原能力将进入到内部的金属离子还原成金属单质。相比于对细胞表面的改造,对细胞内部的改造需要对细胞生理代谢途径有更深刻的理解,并且细胞内部的纳米材料对细胞的影响也有待深入探究,因此这是一种既新颖又有挑战性的方式。
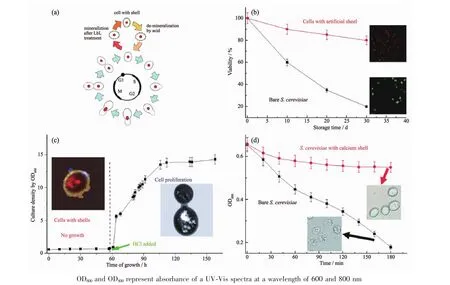
图8 (a)正常酵母和磷酸钙矿化酵母的生命循环示意图;(b)正常酵母细胞与磷酸钙矿化的酵母细胞在纯水中随时间的存活率,插图部分为存放30 d后相应的酵母活性图(酵母live/dead活性染色试剂盒,红色为活细胞,绿色为死细胞);(c)磷酸钙矿化的啤酒酵母脱壳前后生长曲线。矿化酵母不能分裂增殖,在t=60 h脱壳处理(1 mmol·L-1盐酸)后,酵母恢复繁殖;(d)正常酵母与矿化酵母在溶菌酶存在条件下的存活情况[68]Fig.8 (a)Life cycle of normal yeast cells and the encapsulated cells;(b)Curves showing the percentage of living cells in pure water against time;The insets are the corresponding fluorescent images after 30 d(the red spots imply that the cells are still alive,and green spots represent the dead cells);(c)Growth curve of S.cerevisiae cells with and without the mineral shells;Mineralized yeast cells cannot divide and proliferate;1 mmol·L-1 HCl was added at t=60 h to induce the dissolution of the mineral shell,and then yeast cell recovered proliferating;(d)Survival of S.cerevisiae cells in the presence of zymolyase[68]
3 纳米材料对生物体的功能改造及其应用
本文所综述的仿生无机纳米材料改造生物体研究进展,主要是针对细胞和病毒的改造,因为这些低等的生物体通常有一些特有的功能,而且在尺寸上适合做纳米材料的改造,结合纳米材料的尺寸效应,就有可能产生特殊生物-材料相互作用,从而影响或改变生物体的功能,同时生物-材料的界面也对纳米材料的形成造成影响。以下将分类介绍近十年来纳米材料在生物体的功能改造及其应用中的进展。
3.1 生物保护
自然界中的很多生物体,都会受到外界胁迫的影响,尤其是低等的生物体,更加容易受到外界因素影响,所以纳米材料对生物体的保护作用是其他功能改造的基础。目前纳米材料在生物体改造中的应用,有很大一部分是基于对生物的保护作用。由于不同的保护作用,从而产生了不同的功能和应用。
3.1.1 细胞保存
受到鸡蛋壳的启发,王本等[68]通过层层自组装表面改性的方法首次实现了酵母细胞的表面钙矿化,人工赋予了酵母细胞一层700 nm厚的磷酸钙外壳。磷酸钙单细胞包裹后的酵母细胞依然能保持其活性,并且能够控制细胞的分裂,将细胞维持在休眠状态(图8a),因此磷酸钙包裹的酵母细胞比普通的酵母细胞在纯水中的保存时间更长[68]。在1个月的时间内,普通酵母细胞的死亡率接近80%,而磷酸钙包裹的酵母细胞的死亡率约为15%(图8b)。这可能是因为磷酸钙外壳起到了阻碍细胞与外界环境之间的物质交换和细胞之间信息交流的作用。当磷酸钙外壳在pH=5.5的弱酸环境中溶解后,酵母细胞又可以重新分裂(图8c)。这层磷酸钙外壳还能帮助酵母细胞抵抗外来侵害,比如在细胞裂解酶存在下,普通的酵母细胞在3 h就几乎全部发生裂解,而有磷酸钙外壳包裹的酵母细胞大部分保持完好的形态(图8d)。随后Choi课题组[73,84]先后利用仿生硅矿化实现了酵母细胞和HeLa细胞的二氧化硅包裹。硅矿化后的酵母细胞表面有一层30 nm厚的二氧化硅外壳,也具有长期保持细胞活性的作用,在纯水中放置30 d,矿化细胞存活率为56%,而普通酵母细胞仅为24%,并且二氧化硅包裹几乎完全限制了细胞分裂。对HeLa细胞的表面硅矿化虽然会对细胞活性有损伤,但是随着培养时间的延长,未矿化处理的HeLa细胞存活率显著下降,而矿化后的细胞存活率下降得明显更加缓慢。这些工作表明细胞的纳米材料包裹是实现细胞保存的途径,Choi课题组[27,90]还在一些文章中提出了“人工孢子”的概念,也是对细胞保存这个应用的一个印证。

图9 (a)正常酵母细胞未经热处理的切片TEM图片;(b)正常酵母细胞52℃热处理2 h后的切片TEM图片;(c)二氧化硅包裹的酵母细胞未经热处理的切片TEM图片;(d)二氧化硅包裹的酵母细胞52℃热处理2 h的切片TEM图片[75]Fig.9 (a)TEM image of native yeast cell without heat treatment;(b)TEM image of native yeast cell after 2 h treatment at 52 ℃;(c)TEM image of silica-coated yeast cell without heat treatment;(d)TEM image of silica-coated yeast cell after 2 h treatment at 52℃[75]
3.1.2 热保护
热稳定性是生物保存的一个关键因素,因为高温会使蛋白质的构象发生变化,甚至导致蛋白质失活[114-115],而且也会对生物体产生热胁迫,所以提高生物和蛋白制品的热稳定性是十分必要的。自然界中的许多植物如水稻、马尾草和仙人掌等的表面具有二氧化硅矿物层,而且相关研究表明这种矿物层除了提供机械支持和预防病原菌入侵等功能外还具有保持水分和增强其耐热性的功能[116-118]。受到自然现象的启发,王广川等[75]通过层层自组装改性和静电吸附的方式,赋予酵母细胞一层二氧化硅纳米外壳,并且发现二氧化硅包裹的酵母细胞可以在49~53℃范围内保持活性,而普通的酵母细胞在52℃时仅有50%的存活率。在透射电镜下观察细胞切片,二氧化硅纳米外壳在热处理下并没有发生明显的结构变化,而普通酵母细胞壁在热处理时则有明显的形变,这说明纳米二氧化硅的外壳起到了保持细胞结构和限制形变的作用(图9)。此外,Choi课题组[119]通过二氧化硅和二氧化钛的复合纳米外壳也能为小球藻细胞提供热保护的功能。
减毒活疫苗是一种非常典型的生物制品,对于人类对抗传染病具有十分重要的意义,但是它具有温度敏感性,它的运输和保藏过程必须采用低温环境(如冷冻)[120-121]。据统计,由于储藏或运输不当,每年约有50%的疫苗被浪费[122]。因此,改善疫苗的热稳定性对于减少冷链开支和疫苗浪费都具有重要的意义。王广川等[92]通过原位矿化的方式成功制备出磷酸钙矿化的乙脑病毒(JEV)疫苗(图 10a),且矿化形成的磷酸钙外壳在不影响疫苗本身基本生物学和不损失其免疫原性的条件下显著提高了疫苗的热稳定性(图10b)。在室温下,磷酸钙矿化的疫苗可以至少保存1周,而非矿化的疫苗最多保存2 d(图10c)[92]。并且在室温下存放1周后,矿化的疫苗活性和新鲜的疫苗活性相当(图10d)[92]。但是,很多疫苗由于缺乏诱导磷酸钙矿化成核的官能团,难以通过直接矿化的方式制备出这种具有核-壳结构的热稳定疫苗。因此,利用基因工程的手段将成核多肽的基因片段插入到病毒的序列中,就能得到具有自矿化功能的疫苗[99]。表面嵌合了具有促进磷酸钙矿化的多肽的肠道病毒(EV71),可以在富含钙离子的溶液中自发地发生矿化,形成一层纳米磷酸钙层[99]。而且研究发现此磷酸钙自矿化的病毒疫苗具有良好的免疫原性和显著增强的热稳定性。重要的是,这种具有自矿化能力的疫苗是通过基因工程手段制得,因此被赋予的矿化能力是可遗传的。

图10 (a)磷酸钙矿化疫苗的TEM图片;(b)JEV和B-JEV在小鼠肾细胞BHK-21中的一步生长曲线;(c)原病毒 JEV(实心)与磷酸钙矿化病毒 B-JEV(空心)在 26 ℃下的热稳定性;(d)分别用 JEV、B-JEV或热处理以后的JEV、B-JEV免疫小鼠,免疫2周后小鼠脾细胞中γ干扰素表达水平(SFC是指斑点形成细胞)[92]Fig.10 (a)TEM image of B-JEV particles;insert is negatively stained B-JEV;(b)One-step growth curves of JEV and B-JEV on baby-hamster kidney (BHK-21)cells;(c)In vitro assessments of thermostability.Remaining infectivity of JEV (solid)and B-JEV (hollow)after the storage at 26 ℃;(d)Production of INF-γ by T cells of mice immunized with JEV or B-JEV(SFCstands for spot-forming cell)[92]
尽管磷酸钙矿化疫苗的热稳定性显著提高,但是这种疫苗策略仅能在室温环境中储存1周左右。自然进化中从温泉细菌到抗逆植物都选择了无定形硅矿物,而且有研究表明硅可以通过表面羟基与周围水分子形成氢键而禁锢水分子的流动性[123-124]。为了进一步探索生物矿化策略在提高疫苗的热稳定性方面的潜力,王广川等[110]研究了二氧化硅矿物在此方面的表现。实验证明,完整的二氧化硅矿物壳会让病毒疫苗丧失活性,但是不连续的硅矿物层则可实现在不影响疫苗活性的情况下大大增强其热稳定性。通过调节硅酸的pH值,实现了病毒疫苗的原位硅矿化,在疫苗表面引入大量的以锚定形式存在的无定形硅纳米簇。体外实验证明硅矿化的疫苗热稳定性增加了约10倍,可以在室温下保存35 d[110]。但是对于那些表面缺乏矿化活性位点的病毒来说,通常需要对其进行表面化学修饰。通过PEI对病毒疫苗表面进行修饰后[125],可以实现疫苗的表面硅矿化,并且大幅度提高疫苗的热稳定性,以JEV病毒疫苗为例,硅矿化后可以在室温下保存至少15 d。因此,硅矿化比磷酸钙矿化在改善疫苗热稳定性方面更具优势。此外,周航宇等[126]还发现纳米三氧化铝包裹疫苗可以同时提高热稳定性和免疫原性,为疫苗的纳米改造提供了另一种策略。
对生物体的热保护,其实质主要还是保护生物体中的蛋白质和酶,而且蛋白质和酶的保存在药物制品的保存中也具有非常重要的意义。杨宇玲等[127]通过原位矿化的方法将过氧化氢酶包裹于无定形磷酸钙纳米颗粒中(图11),使得过氧化氢酶在活性和结构方面均表现出更高的热稳定性。更重要的是,研究还发现这层磷酸钙矿化外壳除了以其刚性结构为蛋白质起到固定作用外,无定形相中存在的结构水才是给予蛋白质高温保护的关键因素。无定形矿物中稳定的结构水分子与蛋白质紧密结合,以更为稳定的水合物形式抑制了蛋白质与外界环境中水分子的氢键交换,减少高温条件下水分子剧烈运动对蛋白分子结构造成的破坏,从而提高蛋白分子的热稳定性。这种以生物矿化为基础的材料-蛋白相复合的方法不仅为今后蛋白或其他生物大分子的改性提供了一个新的思路,同时也为矿化提高生物体热稳定性建立了一个理论模型。

图11 无定型磷酸钙矿化包裹的过氧化氢酶结构示意图[127]Fig.11 Schematic view of the entrapped enzyme inside the calcium phosphate (ACP)-catalase (CAT)nanocomposite[127]
3.1.3 光保护
随着人类对可再生能源需求的增长,生物质由于其潜在的作用引起了人们高度的关注。光合微生物,包括蓝藻,绿藻和硅藻等微藻,是地球上最重要的初级生产力供应系统[128-130]。以蓝藻为例,从全球范围来看,每年约有250亿吨的碳以二氧化碳的形式被蓝藻固定到高能量密度的生物质中[131-132]。蓝藻几乎存在于所有的陆地和水生环境中,为地球提供了20%~30%的光合生产力并且以大约450 TW的速率将太阳能转化为生物质储存的化学能[128]。所以蓝藻对于解决日益增加的能源需求和环境问题能够起到十分重要的作用。但是,自然界中微生物的光合作用通常是一个低效率的过程,因为它们的光能自养生长总是受到各种环境胁迫的影响[133-134],例如蓝藻的光合作用对强光非常敏感,过度的光照可以显著抑制蓝藻的光合作用甚至会导致光合器官的光氧化损伤[135-137]。因此,光抑制是造成生物质损失的主要原因之一。所以减缓强光胁迫造成的光抑制对增加光合作用产物起着非常重要的作用。
自然界中的硅藻可以通过生物硅矿化形成结构精致的纳米二氧化硅外壳为自身提供机械保护和光子响应[130]。这个纳米结构的外壳也被认为是硅藻具有更高光合作用效率的重要原因。受到硅藻的启发,熊威等[138]选取蓝藻集胞藻PCC 6803(Synechocystis sp.strain PCC 6803)为模式生物,通过仿生矿化的手段人工赋予蓝藻表面一层纳米级的二氧化硅外壳(图12a)。通过研究细胞表面二氧化硅外壳的光学性质,发现二氧化硅外壳可以帮助蓝藻细胞阻挡很大一部分的光照(图12b)[138]。因此,在强光照条件下,二氧化硅矿化的蓝藻比正常的蓝藻具有更高的光合活性(每毫克叶绿素a每小时的氧气释放量)(图12c)[138]。从某种程度上说,这种细胞外的硅矿物成为了生物体对抗强光照射的纳米“防护服”。这样一种基于材料技术对细胞的改性促进了蓝藻在强光下的光合作用,从而增加了光合生物质的产量。生物质合成的增加不仅可以用来生产更多的生物能源,还有助于缓解温室效应。

图12 (a)蓝藻细胞表面仿生硅矿化过程示意图;(b)普通蓝藻细胞(红线)与矿化蓝藻细胞(蓝线)的紫外可见光谱(投射模式);(c)普通蓝藻细胞(红线)与矿化蓝藻细胞(蓝线)在不同光照的以1,4-对苯醌(BQ)为电子受体时的光合放氧速率[138]Fig.12 (a)Procedure for silica encapsulation of individual cyanobacteria.LbL deposition of PDADMACand PSSonto the cell surface induced the formation of silica coating;(b)UV-Vis spectra of native cyanobacteria (black curve),PDADMAC/PSSmultilayer-coated cyanobacteria (red curve),cyanobacteria@SiO2 (blue curve)in the transmission mode;(c)Photosynthetic evolution of O2 with 1,4-benzoquinone (BQ)as the artificial electron acceptor in native cyanobacteria and cyanobacteria@SiO2 cells under different light intensities[138]

图13 (a)纳米二氧化铈一步法包裹小球藻抗紫外保护[111];(b)磷酸镧铈铽外壳保护斑马鱼卵细胞在紫外线辐射下发育[145]Fig.13 (a)Scheme of One-Step CeO2 Nanoshell Formation on the Chlorella Cell for UV Protection[111];(b)Scheme of La0.20Ce0.45Tb0.35PO4 shell for guarding embryo development of zebrafish against UV radiation[145]
随着人类活动的影响不断扩大,地球环境逐渐恶化,其中令人瞩目的一个问题就是平流层中臭氧层密度的下降[139]。在过去的近10亿年间,大气中的臭氧层保护着地球上的生物免受有生物伤害性的中波段紫外线(UVB)的伤害[140]。紫外光辐射不仅会对浮游植物产生胁迫[141],也会对水生动物产生伤害[142-143]。自然界中的颗石藻可以利用表面特殊的球霰石外壳提高它对紫外线的散射能力,从而缓解紫外光胁迫[144]。受到自然界矿化外壳保护生物体的启发,段鹏强等[111]通过构建纳米二氧化铈外壳单细胞包裹小球藻细胞,实现了对紫外光胁迫的缓解(图13a),而王本等[145]通过仿生矿化的手段为斑马鱼受精卵细胞构建了一层磷酸镧铈铽的外壳,这层外壳可以吸收特异性的紫外线辐射,从而为斑马鱼受精卵细胞的发育提供紫外保护(图13b)。
3.1.4 生物隐身
分子识别是生物体内许多界面相互作用的基础,可以帮助生物系统传递信号和抵抗外源性侵害,比如免疫反应就是生物体基于分子识别做出的防御反应[146-147]。但是,目前的生物医学治疗效率严重受到不必要的免疫反应制约。例如,病毒载体由于可以把基因导入各种人体细胞中,已经被用于基因治疗[148-149]。但是活体实验发现病毒纳米载体总是被人体当作外源性生物体识别,进而被抗体中和甚至快速清除[150]。这种不必要的识别和清除极大限制了病毒载体在生物医学中的应用。由于这种识别依赖于病毒表面和特殊生物大分子的特异性相互作用,因此屏蔽病毒表面与生物大分子结合的位点是消除这种识别的一个有效途径。
化学修饰是改变生物界面的常用方法。王晓雨等[151]首先用过阴阳离子聚电解质层层自组装的方法对人类黄热病疫苗17D(YF-17D)表面改性后,发现病毒的感染性被限制了,说明表面抗原被屏蔽了。但是这种方法将病毒纳米颗粒包裹后,当病毒载体进入细胞后,表面这层聚电解质层无法脱除,因此这种改性方式还是有较大的局限性。受到自然界仿生矿化的启发,王晓雨等[152]对腺病毒血清型5(Ad5)实现了原位的磷酸钙矿化,并首次提出了“基于生物矿化的病毒壳工程”这个概念。磷酸钙矿化后的Ad5,可以规避抗体的中和作用,进入到细胞中,并且释放出病毒载体,并且依然具有侵染性[152]。随后,王晓雨等[153]进一步将能表达猴免疫缺陷病毒(SIV)包膜蛋白的重组腺病毒血清型 5(rAd5-Env)进行磷酸钙矿化。矿化后的病毒能规避Ad5抗体的预免疫,最终增强包膜蛋白特异性的T细胞免疫反应(图14a)。这项研究为艾滋病(HIV)疫苗在内的疫苗优化与改造提供了一种全新思路。此外,王晓雨等[154-155]还将磷酸钙矿化修饰用于鼻接种疫苗和登革热病毒的改造,并且实现了对体内预存抗体的免疫,进一步将“基于生物矿化的病毒壳工程”这个概念在生物隐身中应用。最近周航宇等[156]发现磷酸钙矿化的禽流感病毒(图14b)可以同时增强热稳定性和感染性,并可能导致禽流感病毒传染给人类。
分子识别在人体中的另一个重要用途就是识别血型。人的血型是由红细胞表面的抗原蛋白决定的,例如最常见的ABO血型系统,A型血的红细胞表面带A型抗原,B型血的红细胞表面带B型抗原。进行输血时,受血者血浆中的抗体会识别供血者的红细胞表面抗原,如果血型不匹配,抗体就会把它们定义为“外来物种”,并向它们发起进攻,造成严重的后果,甚至还会致命[157]。王本等[85]利用多巴胺表面聚合对红细胞进行改造,通过聚多巴胺的包裹屏蔽了红细胞表面的抗原,使得红细胞在血型错配的情况下,依然不发生抗原免疫反应。这种红细胞改造的方法为“万能血”的制造提供了一种新的策略,但是离实际应用还有一段距离。

图14 (a)矿化病毒克服预存免疫的示意图[153];(b)禽流感病毒磷酸钙矿化示意图[156]Fig.14 (a)Scheme of vaccine engineering with mineral shell for overcoming preexisting immunity[153];(b)Scheme of CaPshell formation on avian influenza virus[156]
3.2 癌症治疗
癌症是全世界导致大量人口死亡的主要原因之一,并且癌症发生率还在持续增加[158-159]。目前,癌症的治疗方法还是限于化疗、放疗和手术。不幸的是,化疗和放疗通常会对正常细胞也产生副作用,而手术虽然可能把原发性肿瘤和可见转移的肿瘤全部移除,但肿瘤总是倾向于侵犯相邻组织或者扩散到其他微小不可见的区域,这是限制手术清除效果的主要因素[160-163]。近年来,纳米技术在癌症治疗中的研究为癌症的治疗提供了很多新的思路和方法[164-168]。但是纳米材料的体内细胞毒性、生物安全性以及在人体内的富集都是富有争议的,因此限制了其在生物医学中的应用。
研究发现,叶酸受体在很多人体癌细胞中的表达会比正常细胞中上调,而叶酸受体可以导致叶酸分子的积累[169]。叶酸分子中的羧基可能特异性螯合钙离子进而诱导生物钙化的发生[169-170]。之前王本等[68]的研究表明,当细胞被矿物包裹后,细胞的生理行为将会改变,特别是细胞的生长和增殖将受到抑制。因此,赵瑞波等[79]提出通过癌细胞的靶向钙化(CCTC)来抑制肿瘤细胞的生长和转移进而达到治疗癌症的目的,并且通过实验证实癌细胞可以通过表面上调的叶酸受体结合更多的叶酸从而诱导磷酸钙在表面矿化沉积,而正常的细胞不会发生钙化(图15a)。在体内实验中,通过癌细胞靶向钙化,肿瘤最后转变为钙化的组织,而钙化会破坏肿瘤细胞的细胞膜最终导致肿瘤细胞死亡(图15b和c)[79]。这项研究通过人工诱导病理矿化,将不可逆的癌症转化为可以治疗的组织硬化,为癌症治疗提供了一种无需药物的新策略。

图15 (a)癌症细胞靶向钙矿化(CCTC)示意图;(b)体内CCTC处理后的肿瘤Micro CT检测照片;(c)体内CCTC处理后的肿瘤组织切片TEM图(内部为钙化组织元素分析图)[79]Fig.15 (a)Scheme of cancer cell-targeting calcification (CCTC);(b)Micro computed tomography (μCT)detection of tumours in vivo CCTCtreatment;(c)TEM images and (inset)energy-dispersive X-ray spectroscopy (EDX)of calcified matter in tumour cellular gaps[79]
3.3 生物催化
微生物能够合成很多不同的酶用于工业催化反应,但是一个重要的问题是这些酶离体后在有机溶剂中很容易被破坏,因此基于全细胞的水/有机两相生物催化是一个很好的实现低水溶性物质转化的方法,但是细胞长期置于有机溶剂中会产生致命的损伤从而降低催化效率[171-174]。于是细胞包埋的策略被用于改善微生物的催化活性和稳定性[175],而这种方式又会降低细胞内外的传质从而影响催化效率。因此需要设计理想的细胞包埋体系使得微生物细胞在保持活性的同时不降低催化效率。在皮克林界面反应体系中,胶体颗粒可以同时稳定乳液并催化两相界面的反应,所以很适合用于生物催化[176-179]。为了改善细菌在皮克林试剂中的稳定性,曲晓刚课题组[175]首先对粪产碱杆菌细胞进行仿生钙矿化包裹并同时负载磁性四氧化三铁纳米颗粒,然后在磷酸钙矿化层表面吸附磷酸单十二烷基酯钠盐(sodium monododecyl phosphate)以增强表面润湿性,最后放入皮克林催化反应体系中(图16)。改造后的细菌在皮克林催化反应体系中,可保持长期的催化活性和稳定性,并且这种生物催化剂可以重复使用30个周期[175]。这种基于细胞壳工程的改造方法,对于其他的两相全细胞催化体系也具有很高的参考价值。此外,曲晓刚课题组[180]还通过仿生矿化的方法为酵母细胞构建了一层二氧化锰纳米酶外壳,纳米二氧化锰外壳可以帮助酵母清除环境胁迫产生的自由基,从而对细胞起到响应保护作用。

图16 磷酸钙包裹的细菌用于皮克林界面生物催化的示意图[175]Fig.16 Scheme of bioconversion by the robust Pickering interfacial biocatalyst of calcium phosphate (CaP)-encapsulated cells[175]
3.4 生物能源
生物能源是指通过生物的活动,将生物质、水或其他无机物转化为沼气、氢气等可燃气体或乙醇、油脂类可燃液体为载体的可再生能源[181]。随着化石能源大量使用带来的环境问题日益严重,生物能源在可再生能源中的重要性越来越引发各国的关注。在所用的生物能源中,以微藻生物能源为代表的微生物能源被认为是最有前景的生物能源形式[181-182]。但是目前微生物能源的开发和利用,还主要受到微生物本身功能的限制,因此,需要寻找理想的微生物或者对微生物进行人工改造。目前,通过纳米材料改造微生物在生物能源中的应用已有初步进展。
3.4.1 光合产氢
氢气被认为是一种有前景的化石燃料替代物,因为氢气燃烧转化效率非常高,对环境友好并且具有高能量容量。但是,当前氢气的生产主要依赖于烃类的裂解、煤的气化和水的电解,然而这些生产氢气的途径本身就是十分消耗能源的,并且是不可持续的[183-184]。因此,氢气的绿色生产一直是科学界和工业界的巨大挑战。光合生物制氢为氢气生产提供了一种可持续的并且是碳中性的途径,因为这个过程直接利用光能和水[185]。在自然界中,光合微生物,尤其是绿藻,能够在光合系统和氢化酶的共同作用下将水光解生成氢气[186-187]。但是,这通常是一个十分短暂的过程,并且只发生在黑暗和光照交替的几分钟之内[188]。这是因为氢化酶遇到氧气极其容易丧失其功能[189]。在黑暗中,细胞的呼吸可以为激活氢化酶创造厌氧的条件[190-191]。在从黑暗转变为光照的短暂时间内,在光合系统II(PSII)反应中心通过水的光氧化反应(H2O→2H++1/2O2+2e-)产生的光合电子能够被传递到氢化酶,在氢化酶的催化作用下与质子结合产生氢气(2H++2e-→H2)[192]。但是,光合作用产生的氧气会使氢化酶迅速失活,所以产氢时间极其短暂并且效率低下,绿藻在自然有氧条件下产氢的大规模应用还尚未现实。
当前,对生物光合产氢的研究主要集中在单细胞水平,但是,自然界还有很多生物代谢过程涉及到细胞形成细胞集群并表现出新的功能[193-194]。聚集能够影响细胞之间的溶质流动和通讯,并因此导致不同于单一细胞的生物学特性产生[195-196]。熊威等[103]通过仿生硅矿化诱导小球藻细胞的聚集,发现聚集的小球藻能在自然有氧的条件下进行光合产氢。在60 mL的玻璃管中加入30 mL的藻液,细胞密度为1.2×108mL-1,置于 100 μE·m-2·s-1的光照下,对于正常的小球藻细胞悬液,在每个检测时间点,顶空中都不能检测到氢气,而小球藻聚集体可以连续稳定的以大约 0.35 μmol·h-1·mg-1的速率(每毫克叶绿素每小时所释放的氢气含量,下同)释放氢气[103]。这个速率大约是自然界中瞬间生物光产氢速率(大约0.20 μmol·h-1·mg-1)的 1.75 倍[103]。更重要的是,小球藻聚集体的光合产氢过程可以持续至少48 h(图17a)[103],而不是短暂的几分钟。小球藻聚集体中的氢气和氧气分布在小球藻细胞聚集体中,存在着空间上的功能分化(图17b和c)。表层的细胞暴露在外界环境中,它们的功能与正常的细胞相似[103]。但是外部的细胞还能在一定程度上起氧气防护壳的作用,阻挡外界氧气渗透到聚集体内部中。聚集体内部的细胞通过呼吸作用消耗扩散进来的氧气和光合作用产生的氧气,从而创造一个缺氧的微区[103]。在这个微区中,氢化酶和PSⅡ的活性可以同时得到维持,为可持续的光合产氢提供了保证。

图17 (a)正常的小球藻和聚集的小球藻在100μE·m-2·s-1的持续光照和有氧条件下的光合产氢积累量和密封管顶空中的氧气体积分数(重复实验次数n=6);(b)小球藻聚集体的空间功能分化;(c)不同尺寸大小的小球藻聚集体中氢气(蓝色)和氧气(红色)浓度分布(n=5)[103]Fig.17 (a)Amount of H2 and the content of O2 in the headspace of sealed tubes under a light intensity of 100 μE·m-2·s-1 at different time periods (n=6);Red lines:O2,Blue lines:H2;Open squares:native Chlorella;Filled circles:aggregated Chlorella;(b)Spatial-functional differentiation in aggregated Chlorella cells;(c)H2 (blue)and O2 (red)microprofiles of aggregated Chlorella with different sizes (n=5)[103]
这是人类首次通过材料技术实现有氧条件下长时间可持续的微藻光合产氢,该突破对于实现大规模的微藻光合产氢和促进绿色能源的发展都具有非常重要的理论指导意义。这种基于化学和材料的细胞改性手段不仅具有可行性高、成本低和效果好等特点,而且可以用于光合作用微生物细胞的改性,也能拓展到诱导其他类型微生物的功能转变。
3.4.2 生物人工光合作用
随着人类发展对可再生能源需求的增长,对太阳能的利用越来越引起大家广泛的关注和重视[197-199]。光合作用是自然界利用太阳能最主要的形式,人工模拟光合作用一直以来都是科学家们的梦想。放氧的光合作用是地球上最主要的光合作用,也是对人类最为重要的,因为它不仅为人类的生存提供氧气,还提供初级生产力。放氧的光合作用主要分成2个部分,在光反应阶段光合系统分解水产生氧气和电子;在暗反应阶段电子参与二氧化碳还原固定过程最终生成碳水化合物。目前,光反应阶段已能通过光催化和光电催化水分解实现,这个过程可以在放出氧气的同时产生氢气[198-201]。受到自然界光合反应过程的启发,利用生物体系中的光合系统或者氢化酶与光催化纳米材料复合互补,能够实现更高效的光催化水分解。在暗反应阶段,通过人工光合体系目前能够实现二氧化碳的还原,但是只能得到低碳的产物,并且缺乏可控性和选择性[202-204]。而在自然的光合作用中,暗反应的产物是具有选择性 的[205-206]。因此,生物与人工光合体系的有机结合是一个可行的方案。Sakimoto等[90]通过硫化镉纳米颗粒对热醋穆尔氏菌(Moorella thermoacetica)进行改造,使得这种非光合作用的细菌,能够在可见光下,利用硫化镉纳米颗粒产生的光生电子在细胞内通过Wood-Ljungdahl途径还原二氧化碳得到醋酸(图18)。这项研究不仅对于生物人工光合体系的构建具有重要的指导意义,也拓展了纳米材料在生物体改造中的应用。
最近,魏炜等[105]通过合成物学的手段构建了一种能在膜上表达重金属螯合蛋白PbrR和在细胞内部表达[NiFe]氢酶的大肠杆菌(图19)。硫离子先在大肠杆菌表面利用PbrR结合镉离子,形成硫化镉纳米颗粒。硫化镉纳米颗粒作为一种常用的可见光响应的光催化材料,可以充当类似光合系统II(PSII)的角色,为细胞提供光生电子源。他们受到熊威等工作的启发,通过仿生硅矿包裹,实现了大肠杆菌的聚集,并诱导了大肠杆菌聚集体在有氧条件下的光合产氢。这项工作利用合成生物学方法与纳米技术,实现了对生物体的定向改造,为生物改造提供了新的思路。
3.5 环境保护
作为地球上主要的初级生产者,蓝藻在地球上分布广泛,并且对地球的大气层和生态圈产生了深远的影响[207-208]。但是蓝藻的大量繁殖也会带来严重的环境问题和健康威胁[209-210]。藻类的大量繁殖现象发生在淡水中称之为水华,发生在海洋中则称之为赤潮[211-212]。其中,蓝藻水华是目前困扰全球最主要的水环境问题之一,并且发生率呈逐年上升的趋势。蓝藻水华爆发后,不仅会向水体中释放藻毒素,严重危害动物和人类的健康,而且会使水体溶解氧迅速下降,破坏水生环境的生态圈,并由此引发一系列其他环境问题[213-214]。为了避免和减轻蓝藻水华带来的危害,人们发明了很多种策略。目前最常用的处理方法,一是使用化学杀藻剂和絮凝剂[215-216],二是人工捕捞[217-218]。这两种方法,前者缺乏生物选择性,并会产生二次污染,给水环境带来其他不利影响,而后者则需要消耗大量的人力和物力,成本太高。更重要的是,目前人们采用的处理蓝藻水华的方法都主要集中在水华爆发后的处理,而并不能抑制蓝藻水华的爆发。由于水体的富营养化是水华爆发的最重要因素之一,因此减少水域和沉积物中的磷负载量是目前最广泛使用的方法,但是这种方法成本太高,也不适合大范围应用。所以,开发更加安全有效的抑制蓝藻水华爆发的方法对人类来说是一个巨大的挑战。

图18 (a)热醋穆尔氏菌-硫化镉复合体及其自光敏化太阳能转化;(b)热醋穆尔氏菌-硫化镉复合体中太阳能-化学能转化的可能途径[90]Fig.18 (a)Depiction of the M.thermoacetica-CdShybrid system and self-photosensitization of it for solar-to-chemical conversion;(b)Possible pathway for solar-to-chemical conversion of the M.thermoacetica-CdShybrid system[90]

图19 表面展示的生物复合技术诱导空气环境中的光驱动产氢[105]Fig.19 Proposed surface-display biohybrid approach to light-driven hydrogen production in air[105]
自然界中的许多蓝藻可以在细胞内部形成气囊,这些气囊可以帮助它们在水体中上浮以捕获更多的太阳光[219-221],正是这些漂浮的蓝藻在水面大规模集聚形成了水华。相比于蓝藻,自然界中的硅藻的细胞壁由具有特殊结构的二氧化硅构成[222]。在自然界中,硅藻利用硅化的细胞壁作压舱物[17,223],使细胞保持沉在水底的状态,在水底光照不足和温度较低的条件中,硅藻细胞的光合生长也受到抑制,所以硅藻相比于其他微藻而言,生长较为缓慢,更不易爆发水华。受到这个现象的启发,熊威等[104]以水华微囊藻(Microcystis spp.)为研究对象,模仿生物硅矿化的策略,在聚二甲基二烯丙基氯化铵(PDADMAC)的诱导下,实现了蓝藻与二氧化硅纳米颗粒的直接复合(图20a),与二氧化硅复合后的蓝藻迅速沉降到水底(图20b),并延缓光合生长,从而抑制蓝藻的大量繁殖和藻毒素的释放(图20c),并且水体中的氧浓度得以维持(图20d)。此外,这个方法不仅对实验室培养的微囊藻有效果,对太湖水以及野外现场试验都有良好效果[104]。这项研究不仅为抑制水华爆发提供了一种全新的策略,对全球水环境和水资源的保护有重要意义,而且此项研究所采用的处理方法可简便快速地诱导蓝藻细胞的聚集沉降,亦有望应用于微藻细胞的富集,降低微藻生物能源的利用成本。

表1 纳米材料改造生物体的方法及应用Table 1 Approaches and applications of modification of organisms by nanomaterials
综上所述,表1列出了近年来纳米材料改造生物体的一些成功案例。

续表1
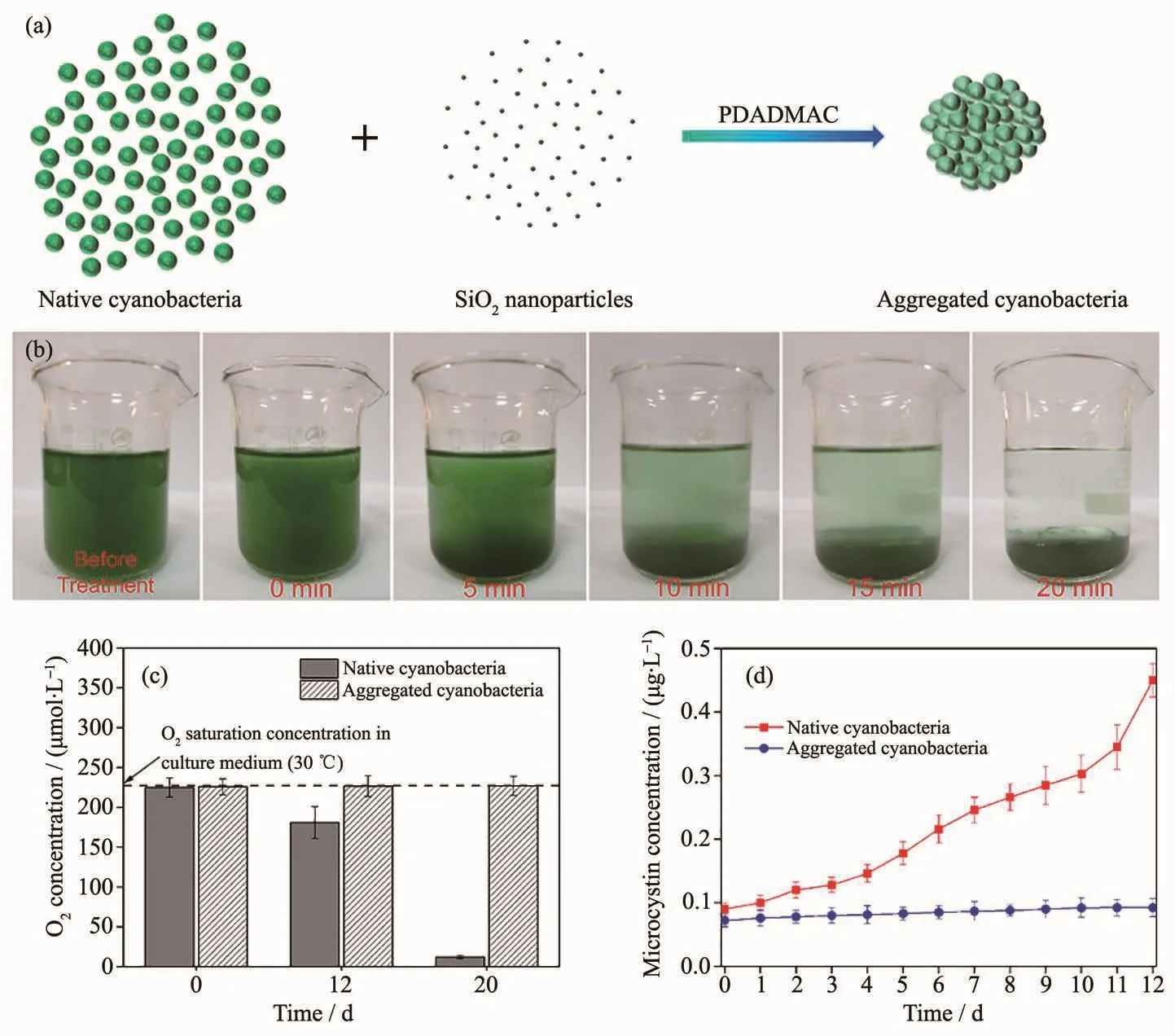
图20 (a)利用PDADMAC直接诱导二氧化硅纳米颗粒与蓝藻结合示意图;(b)正常的蓝藻处理后聚集的蓝藻在烧杯中的分散状态随时间的变化;(c)正常的蓝藻和聚集沉降的蓝藻的细胞悬液中氧气浓度的变化(n=5);(d)正常的蓝藻和聚集沉降的蓝藻的细胞外微囊藻毒素浓度(n=5)[104]Fig.20 (a)Scheme of the direct incorporation of silica nanoparticles on the cyanobacteria using PDADMAC;(b)Native cells (before treatment)and aggregated cells in the beaker (100 mL)at different times,which demonstrated rapid sedimentation of the cells in the presence of silica and PDADMAC;(c)Oxygen concentration in the culture medium of native and aggregated cyanobacteria (n=5);(d)Extracellular microcystin concentration of native and aggregated cyanobacteria (n=5)[104]
4 总结与展望
在漫长的生物进化过程中,生物体在自然选择的作用下,渐渐学会了利用材料来改变自身的功能,去适应环境,进而迈向更高等的生命形式。进入21世纪以来,随着生命科学的发展和纳米技术的进步,受到自然进化过程的启发,科学家们尝试通过材料来改造生物的功能。经过近十年的研究和探索,仿生无机纳米材料改造生物体在方法和应用上都取得了全面的进展,从研究者的数量和分布,文章的档次和数量来看,这个领域已初具规模。目前,通过仿生矿化的手段实现生物体的纳米材料改造是生物相容性最好的途径之一,不过仿生矿化可选择的材料种类仅限于磷酸钙、碳酸钙、二氧化硅等,而这些材料能够赋予的功能有限。实现改造的生物体则涵盖了病毒、原核微生物、真核微生物细胞、动物细胞等几乎所有的生物体基本组成单元。对细胞的改造主要以酵母和大肠杆菌为模式生物居多,这些研究主要是探索改造方法和材料体系为主,应用主要是实现细胞保护。对病毒的改造,主要是集中在提高疫苗热稳定性和病毒载体生物隐身性能方面,也属于利用材料赋予生物保护的功能。对其他特定细胞的改造,比如微藻,红细胞,癌细胞等,由于细胞自身的功能和特性,通过特定的纳米材料可以赋予其相应的功能和应用。
尽管过去的10年,仿生无机纳米材料改造生物体已经取得了重大的发展,并且这个领域的整体框架已初步形成,但是目前这个领域已经报道的研究还是处于初级探索阶段,研究的思路主要是选择材料与生物体结合并寻找新功能。接下来这个领域还需要在材料与生物体的结合方法,材料与生物体的作用机制以及新的功能与应用方面进行更广泛的探索和更深入的探究。
(1)材料与生物体的结合方法:目前的仿生无机纳米材料改造生物体的过程以及材料本身都会对生物体本身有损伤,因此纳米材料的生物相容性和生物安全性是需要在今后的研究中改进和提高的。新的材料体系和结合方法是探索新功能和新应用的基础,纳米材料技术的进步是这个领域发展的重要驱动力。如果能实现纳米材料在生物界面的可控形成与结合,将有助于实现对生物体的精准改造。
(2)材料与生物体的作用机制:虽然目前已经实现了对多种类型的不同生物体的纳米材料改造,但是对于材料与生物的界面相互作用是否影响生物体,还缺乏深入的认识和理解。材料与生物体的作用机制是连接纳米材料和生物功能的“桥梁”,是这个领域的核心科学问题。对于材料与生物体作用机制的进一步认知不仅可以指导材料与生物体结合方法的改进,也能预示新的功能和应用。
(3)新的功能与应用:纵观这10年来仿生无机纳米材料改造生物体的应用探索,我们可以发现,起初的应用都是基于纳米材料对生物体的保护功能,而保护的功能有很多种,这也和自然界中大多数矿化外壳的功能类似。随着改造方法和材料-生物体系的改变,逐渐发现了新的功能和应用,比如生物能源、环境保护、生物催化和癌症治疗等都是最近几年才发现的应用前景。新的改造方法和新的生物体系是新功能探索的物质基础,材料与生物体的作用机制是理论基础,同时探索新的功能和应用也将促进对纳米材料与生物体相互作用机制的研究。
总之,仿生无机纳米材料改造生物体是一个涉及化学、材料、生物、医学、环境等多学科相互交叉的研究领域,正引起越来越多的研究者关注和参与。过去10年的研究为这领域未来的发展打下了坚实的基础。未来这个领域不仅在原有的基础上还有进一步完善的空间,在新的方法、机制和应用方面更加具有广阔的前景。
致谢:美国耶鲁大学医学院王广川博士,浙江大学求是高等研究院王晓雨博士,中国医学科学院苏州系统医学研究所周航宇博士,浙江理工大学材料与纺织学院赵瑞波博士提供相关引文图片和宝贵建议。
