大豆油脂的合成途径及关键酶GPAT基因的研究进展
2019-01-09鲁中爽刘思言李广隆刘明明么梦凡关淑艳
鲁中爽,刘思言,李广隆,王 蕊,刘明明,么梦凡,关淑艳,姚 丹
(吉林农业大学生命科学学院,吉林 长春 130118)
大豆〔Glycine max(Linn.)Merr.〕隶属于豆科大豆属,为一年生草本植物[1],在全球范围内都有广泛种植。大豆含有丰富的脂肪酸、蛋白质及多种营养物质[2-3],其中大豆油脂目前仍占据世界总食用植物油脂的首位。我国不仅是大豆原产国,也是世界上最大的大豆进口国,由于国产大豆的产量和产油率不高,随着石油资源的日益缺乏、人类对高质量油脂需求的增长、对可再生资源的依赖性和环保意识的增强,培育各种比例更健康的油料和提高大豆油脂含量有很大的市场需求。面对国内对豆油的需求量日渐提高,以及在未来较长一段时间内我国仍需继续进口大量大豆及豆油的局面将很难发生实质性改变,因此提高我国大豆种子中的油脂含量,获得高油转基因大豆用于大豆油的生产具有重要意义,科研工作者有义务研究如何更好地提高大豆植物油脂含量及品质[4-5]。
随着生命科学的迅速发展,大量植物功能基因被分离并克隆出来,合成植物油脂的代谢途径中某些调控基因的分子机制和表达调控方式的阐明,对利用基因工程手段从分子水平上有针对性地改良大豆油料作物,生产有用的脂肪酸、提高植物油脂的质量及增加机体的抗逆性等发挥重要作用,这种新发展的分子育种冲破了传统育种的阻碍,是一种很有研究价值的育种手段[6-7]。三酰甘油是大豆油脂的主要成分,而脂肪酸是三酰甘油的来源产物,因此调节终产物三酰甘油和脂肪酸的合成表达,是提高大豆油脂的主要途径之一。在三酰甘油的合成过程中,甘油三磷酸酰基转移酶(GPAT)是合成途径中第一步反应关键酶,生物合成的限速酶,最终将脂肪酸催化形成三酰甘油,改变GPAT活性影响大豆油脂的组成和积累。因此通过分子育种技术手段过表达GPAT基因来改良大豆油脂并对油料作物育种有重要意义。
1 大豆油脂
1.1 大豆油脂的组成
大豆油脂的化学成分为三酰甘油(triacylglycerol,TAG),由甘油和3分子长链脂肪酸在多种酶的催化下形成,因此脂肪酸是植物油脂的重要组成成分,植物油脂的质量及在各方面的用途受脂肪酸的组成影响。
大豆油脂主要在质体和内质网中合成,脂肪酸前体在质体中通过聚合反应生成饱和脂肪酸,在硬脂酰-ACP脱饱和酶(SAD)的催化下生成18∶1-ACP,作为游离脂肪酸的底物,在酰基辅酶A合成酶的作用下,合成酰基辅酶A并进入内质网中。后通过一系列酶的作用合成油脂前体 -TAG[8]。
1.2 大豆油脂的代谢合成途径
油脂的合成代谢是维系其生命活动的基本代谢之一,也为人类提供了重要的能量来源,主要分为:脂肪酸的从头合成、脂肪酸的脱饱和、三酰甘油的合成及油体形成,合成途径如图 1 所示[9]。
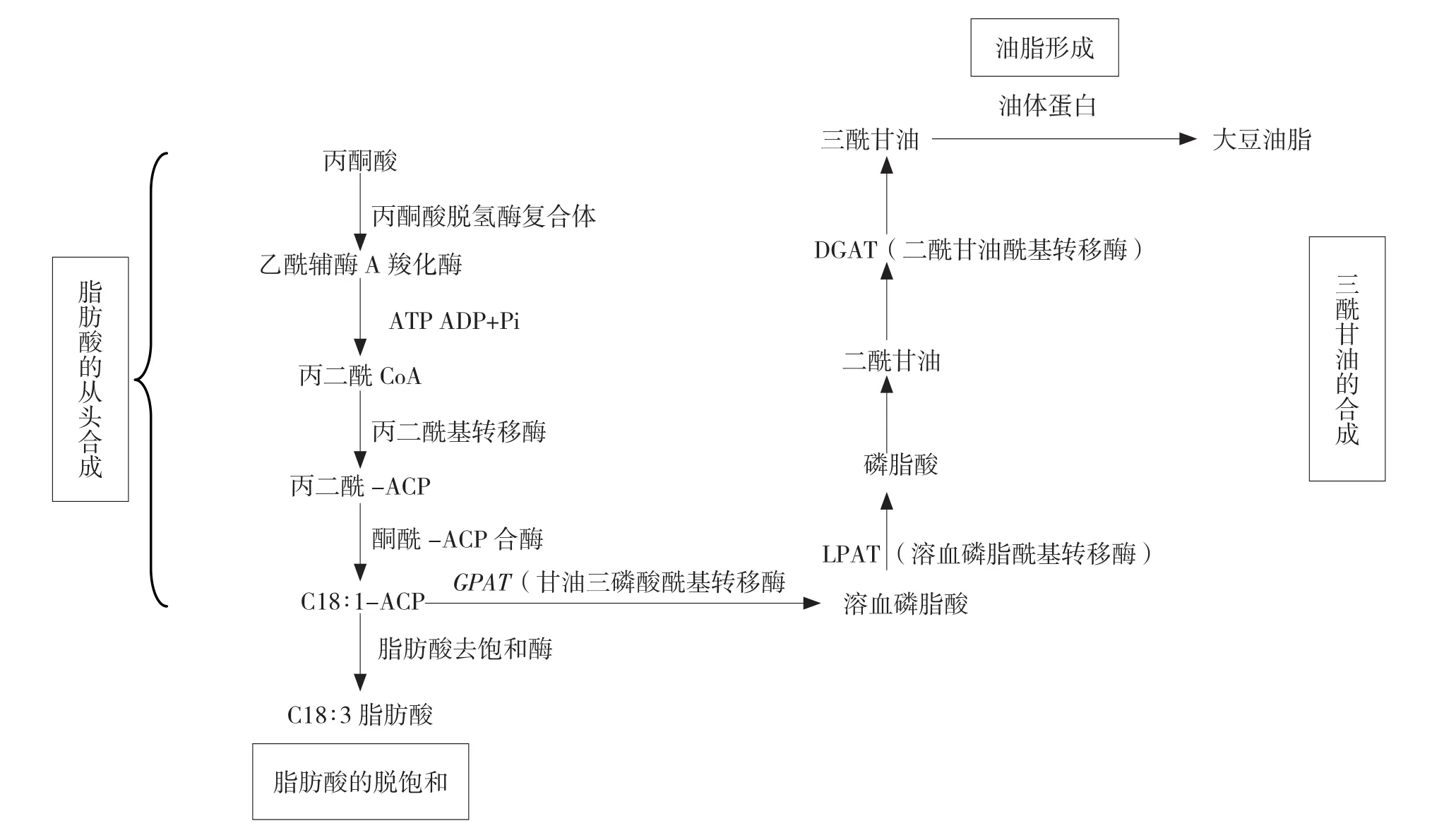
图1 大豆油脂代谢合成途径
1.2.1 脂肪酸的从头合成 蔗糖是油脂合成的代谢的主要碳源,蔗糖通过糖酵解、磷酸戊糖途径和卡尔文循环生成磷酸二羟丙酮和乙酰辅酶A作为三酰甘油的合成原料[10]。乙酰辅酶A在乙酰基辅酶A羧化酶作用下生成丙二酰辅酶A,再与酰基载体蛋白结合,在丙二酰辅酶A酰基载体蛋白转移酶生成丙二酰-ACP(MCAT)的催化下生成丙二酰ACP。随后,一分子乙酰辅酶A与多分子丙二酰辅酶A在脂肪酸合酶复合体的作用下,经缩合、羰基还原、脱水,再还原多次循环,每多一个循环β-酮脂酰基ACP比上一循环多两个碳原子的酰基ACP逐步生成含12~18个碳脂肪酸[11]。
1.2.2 脂肪酸的脱饱和 SAD催化18:0-ACP形成18∶1-ACP单不饱和脂肪酸;另外通过酰基-ACP硫酰酶(FAT)催化部分18:1-ACP单不饱和脂肪酸生成自由脂肪酸。自由脂肪酸在长链酰基辅酶A合成酶的作用下生成酰基辅酶A为三酰甘油的合成提供底物并运到内质网上。另外在一系列酶的作用下将剩下的18∶1-ACP单不饱和脂肪酸生成磷脂酰胆碱(PC),PC是FAD的唯一底物来源。在FAD 的作用下生成亚油酸、亚麻酸[12]。
1.2.3 三酰甘油的合成 Kennedy途径即脂酰辅酶A和甘油三磷酸在内质网中先由GPAT催化形成1-单酰甘油三磷酸;随后在溶血磷酸酰基转移酶(LPAAT)催化下形成PA;磷脂酸磷酸化酶使PA去磷酸基,形成DAG;最后在DGAT的作用下与另一分子脂酰辅酶A形成TAG。另外磷脂酰胆碱(PC)sn-2位上的酰基在磷脂二酰甘油酰基转移酶(PDAT)作用下转移到DAG上,形成TAG和LPAA。还有逆脂酰转移酶(TA)将二酰甘油上的一个脂酰基转移到另一个二酰甘油上,形成三酰甘油和一酰甘油[13-14]。三酰甘油所含有的3个脂肪酸可以是饱和脂肪酸也可以是不饱和脂肪酸,可以是相同的脂肪酸也可以是不相同的脂肪酸[12]。
1.2.4 油体形成 油体蛋白与三酰甘油形成油体。三酰甘油在油体中成液态,油体蛋白在外面包裹的半单位膜中,由半单位膜包裹液态基质三酰甘油脂形成油体。而高含量的三酰甘油可以在内质网膜脂质双层膜结构中形成“脂质凸起”进而从膜结构中分离出来,形成油体。烘干种子过程中植物油体蛋白能够保持油体稳态,并在油体之间相互作用和改变油体膜结构中起到重要作用[15]。
1.3 大豆油脂的功能
大豆油脂是国际市场上重要商品之一,是医药、化工、生物柴油等原料[16-18],也是居民餐饮中重要的食物能源,同时还可作为一种可再生的生物能源。随着国内油料消费增加,油料加工产品需求的快速增长,带动了国内油脂油料种植业的发展,因此研究如何提高大豆油脂的产量至关重要。
脂肪酸作为细胞组成的重要成分又是重要能源物质,还是某些信号分子的前体,与人类的生活紧密联系,很多研究者一直很重视对脂肪酸的研究[19]。大豆油脂的结构成分主要是不饱和脂肪酸,对维持植物膜系统液晶状态和延长植物细胞有重要作用,同时构成叶绿体膜骨架的主要成分也是不饱和脂肪酸,起着对类囊体稳定的作用[20]。大豆不饱和脂肪酸对人体具有降低人体中胆固醇含量、减少脑血栓的形成的作用,对动脉硬化和心脑血管疾病的预防和治疗起到保健作用,而过多摄取饱和脂肪酸可增加患有心血管疾病的风险。提高不饱和脂肪酸中的亚油酸、亚麻酸可增加生物燃料释放的能量、降低废渣温度而提高利用率,高油酸的生物燃料的氧化稳定性更高,油脂中亚麻酸含量高会导致大豆油保质期减短,大豆油易酸败,而人体中不能合成身体所需的亚油酸和亚麻酸的必须脂肪酸,必须从植物中摄取得到,因此根据人体和工业需求不同,培育不同配比脂肪酸的大豆品种具有可行性,提高大豆油脂具有一定的应用价值[21]。
2 甘油三磷酸转移酶(GPAT)
2.1 GPAT基因分类
GPAT基因在拟南芥、菠菜、向日葵、豌豆、水稻等多种植物中被克隆出来,在拟南芥中发现GPAT基因家族有ATS1、AtGPAT1/2/3/4/5/6/7/8/9 等10个成员,其中ATS1定位于质体,AtGPAT1/2/3定位于线粒体,AtGPAT4/5/6/7/8/9定位于内质网[22-23]。AtGPAT1/4/5/6/7/8具有sn-2酰基转移活性,AtGPAT4/6/8还具有磷酸酶活性[24]。很多研究表明AtGPAT1与养分的分泌和绒毡层分化有关[25];AtGPAT4与植物茎、叶中角质的合成有关,AtGPAT5与木栓质脂肪族化合物的积累有关,影响木质栓的合成,AtGPAT6/8与花瓣中角质的合成有关,AtGPAT9与家族中其他几个成员的一致性在10%左右,与动物中GPAT4结构域有很高的同源性,参与脂肪细胞中脂肪酸的合成,并且不同植物的GPAT9氨基酸的保守性很高,在种胚发育期表达量显著提高,导致AtGPAT9对大豆油脂的积累起到重要的调控作用。但GPAT家族中哪个基因在种子发育中大量表达仍需研究[26]。
2.2 GPAT基因分子特征
GPAT的C端序列较为保守,在靠近C端的区域所有GPAT基因都含有脂酰基催化结构域,该结构域中有4个(模体I、Ⅱ、Ⅲ、Ⅳ)高度保守的模体。这些模体对GPAT的脂酰基转移酶活性及其结合G3P底物至关重要。不同的GPAT的N端序列差别较大,因此不同的GPAT之间的分子大小、生化特性及亚细胞定位也有所不同。同时不同的GPAT具有(脂酰基链长度、不饱和度及ω-氧化程度的选择性)不同的脂酰基种类, 从而参与不同的脂质代谢过程[27]。
在内质网上,GPAT是三酰甘油合成途径中第一关键酶,主要负责将饱和脂肪酸酰基转移到甘油三磷酸(G-3-P)的sn-1号位上,形成溶血磷脂酸(LPA)。随着研究发现,GPAT还可将酰基转移转移到G-3-P的sn-2号位上,可产生sn-2LPA或sn-2-单酰甘油(sn-2MAG),但具有酰基转移sn-2位特异性的GPAT只在陆生植株中被发现,因此生化分子生物学国际联合酶学委员会命名这一具有酰基转移到sn-2位上的特异性GPAT酶为EC2.3.1.198[24]。
大多数GPAT酶(GPAT1、GPAT2、GPAT3、GPAT4 和GPAT5)已被报道居住在内质网中,通过保守的跨膜基序锚定在那里。GPAT1受位于30 kb间的近端和远端启动子的调控,在内质网合成三酰甘油(TAG)的第一步限速步骤是由GPAT3 或GPAT4催化的,它将G-3-P和脂肪酰辅酶A(FA-CoA)转化至LPA。然后通过GPAT酶将LPA转化为磷脂酸(PA),PA随后通过脂蛋白PAP酶(lipin 1,2或3)去磷酸化为二酰甘油(DAG),TAG生物合成的最后一步是将DAG转化为由DGAT1和DGAT2催化的TAG,DGAT1和DGAT2的内质网膜定位可能是不同的[28-29]。G-3-P途径的酶除了调节TAG合成外,也影响磷脂合成,如:由GPATs产生的PA可作为合成酸性磷脂、磷脂酰乙醇或磷脂酰肌醇、心磷脂的底物;由脂蛋白PAP酶作用产生的DAG可以转化为TAG或磷脂、磷脂酰胆碱、磷脂酰乙醇胺和磷脂酰丝氨酸[30]。
2.3 GPAT功能
GPAT催化酰基供体向酰基转移[31],磷脂酰甘油(PG)在酰基脂化酶GPAT的催化下,GPAT具有ACP的结合位点,它能催化酰基辅酶A或酰基ACP到脂肪酰基的酯交换[32],以为酰基供体的GPAT酶定位于质体,属胞质溶解型GPAT;以acyl-CoA为酰基供体的GPAT酶定位于内质网和线粒体属膜结合型GPAT。将acyl-ACP的酰基转移到甘油三磷酸的sn-1位上合成溶血磷脂酸(LPA)[33],该反应构成储存-脂质组装的第一步,并且在极性和细胞外脂质生物合成中也是重要的[34]。将acyl-CoA的酰基转移到LPA的sn-2位上,溶血磷脂酸酰基转移酶(LPAT)催化形成磷脂酸(PA),后续反应之后形成 TAG[23]。
由于GPAT酶酰基转移位点特异性和底物选择性为脂质合成提供了底物,GPAT在植物生长发育过程中发挥重要作用,不同GPAT基因发挥着不同的表达模式,表现出多种生理功能:(1)与植物的育性有关:在拟南芥花粉发育过程中,AtGPAT1/6共同影响拟南芥的结实率,AtGPAT1/6突变体会阻碍雄蕊长纤丝的伸长和导致小孢子的释放,减少授粉概率导致结实率降低[35];(2)与植物的耐高温、抗低温有关:在酶学的研究中发现两类GPAT,一是可以优先选择188∶1脂肪酸为底物的选择型GPAT,二是以16:0脂肪酸为靶向底物优先选择18∶1脂肪酸的非选择型GPAT,一些研究表明选择型酶是耐冷植物,非选择型酶是冷敏感植物,在较低温度下,顺式不饱和磷脂酰甘油(PG)比饱和磷脂酰甘油能更好地防止膜相变,在低温下的流动性及生物膜的饱和度影响耐冷性[36];(3)与植物的耐干旱和耐高浓度盐有关:AtGPAT突变体导致拟南芥幼根和种皮中软木脂含量降低,由于软木脂阻止病原体入侵植物的物质,并能控制溶质和水分的运输,因此在高盐下容易失水,使得渗透性急剧增加和导致存活率降低[37];(4)与种子含油量有关:GPAT基因是脂肪酸合成途径中第一关键酶,拟南芥种子总脂肪酸的含量高达30.5%~40%,目前在拟南芥中已鉴定出9种GPAT基因,可见GPAT与油分组成相关,可提高种子重量和含油量[38];(5)其他功能:具有磷酸酶活性,可能因GPAT基因编码的酶其-N-端存在HAD-like 结构域有关,GPAT可对受伤植物进行修复伤口的功能,GPAT基因催化酰基辅酶A或酰基ACP到脂肪酰基的酯交换具有ACP的结合位点[37]。
3 GPAT基因在大豆油脂合成途径中的应用
最近几年GPAT基因作为调节脂肪酸的关键基因,其表达涉及油脂的合成在某些植物中的应用相继开展。2013年,郝静芳[39]认为GPAT7 和GPAT9参与拟南芥种子中TAG的合成及一些不饱和脂肪酸和调节。2014年,刘聪[37]在研究甘油型油菜的GPAT6基因的克隆与分析中发现GPAT6基因改变了拟南芥种子脂肪酸成分的比例;Sui等[40]在拟南芥中的通过过表达SsGPAT来提高磷脂酰甘油的不饱和脂肪酸含量,可以增强拟南芥耐盐性并缓解盐胁迫下PSII和PSI的光抑制。2016年,Niu等[41]在三角褐指藻中克隆了GPAT基因并在三角褐指藻中过表达,通过GC-MS分析脂肪酸组成,显示与野生型相比有更高的不饱和脂肪酸比例,鉴定出的GPAT可以上调三角褐指藻中TAG的生物合成。2017年,Srinivasan等[42]发现AGPAT1过表达显著改变了初级代谢,总脂质含量增加,但总碳水化合物和可溶性蛋白质含量降低,结果表明AGPAT1过表达可以提高TAG的生物合成;皮广静[43]在研究花生油脂合成酰基转移酶基因表达与籽仁含油量相关性分析时发现,GPAT3个时期平均表达量与含油量显著正相关;Aparna等[44]克隆了麻风树的两种GPAT基因并导入拟南芥中过表达,发现总油脂含量增加等。以上研究都证明了GPAT基因参与到调解脂肪酸的过程,对油脂的合成起到关键性作用。上述事例虽然都证明GPAT基因改变了油脂的含量,却是不同GPAT成员对不同物种的油脂合成起到调控作用,其他物种中的GPAT基因的功能是否对油脂的合成起到调节作用还有待研究。
由于GPATs在不同时期和不同组织的表达模式存在显著差异,预示着其参与不同的脂质代谢和生物学过程中GPAT的不同。通过这些实例可以表明GPAT基因的表达影响脂肪酸的合成,对油脂含量起到调控作用。GPAT基因作为油脂合成途径中关键的酰基转移酶,大豆中也含有GPAT基因,但至今还未验证大豆GPAT基因是否对油脂的合成起到调控作用,同时大豆GPAT基因的活性和某些生物学功能还有待进一步研究。因此研究GPAT基因表达及促进大豆油脂合成的功能性分析可能对获得高油转基因大豆用于大豆油的生产具有重要意义。
4 展望
大豆油脂合成涉及大量的基因及其编码的酶类,是一个极其复杂的过程,改变任何一个因素都可能造成植物含油量或者油酸成分的改变,因此弄清大豆油脂合成的每一个环节都具有重要意义。虽然GPAT基因作为油脂合成的关键酶,但目前对植物GPAT基因家族的认知却主要集中于拟南芥中。植物GPAT基因家族虽然只是一个小家族,却具有多种功能,GPAT基因的多样性和重要性很容易激起科研工作者对其深入研究的热情。GPAT酰基转移酶位点特异性和对底物的选择性为什么会对植物的生长发育起到调控作用、单个还是多个GPAT 成员对油脂代谢合成途径起作用以及大豆中GPAT基因的生物学功能等问题还有待探索。
随着人们对生活质量要求的提高,现在对油料作物的开发与利用越来越重视,油料植物不仅可供人体需求还可作为环保清洁能源,在生活中很多方面都有应用,科研工作者也越来越重视油料作物研究趋向人类需求的方向。大豆作为世界较大的产油作物,GPAT基因也是高等植物中油脂合成途径中的关键酶,虽然GPAT基因在很多植物中都克隆出来并进行了功能验证,但在大豆中的相关报道很少,利用基因工程等新科技手段,对大豆合成三酰甘油途径中第一关键酶GPAT基因的克隆及验证功能还需科研工作者进一步研究。
