高黎贡山北段东、西坡捕食线虫真菌分布格局研究
2019-01-04王家亮刘硕然周汉宇房以好杨晓燕
王家亮 ,刘硕然 ,周汉宇 ,房以好 ,肖 文 ,杨晓燕 *
(1.大理大学公共卫生学院,云南大理 671000;2.大理大学东喜玛拉雅研究院,云南大理 671003;3.云南省高校洱海流域保护与可持续发展研究重点实验室,云南大理 671003)
微生物物种资源极为丰富,是地球生物多样性的重要组成部分〔1〕。微生物的空间分布格局受到众多驱动因子的影响,主要包括当代环境条件(温度、降水、光照、土壤养分、pH)和历史因素(地理阻隔、物理屏障、扩散限制等)〔2-3〕,而温度、降水等环境因子与山地微生物物种丰富度有着密不可分的联系。研究人员发现坡向对土壤的温度、含水量等理化性质均有影响〔4〕。不同坡向上的水分和光照强度存在差异,从而影响土壤微生物的生长和分布格局〔5〕。现阶段有关坡向对土壤微生物分布格局影响的研究还较少。
高黎贡山国家级自然保护区东坡垂直落差高达6 000 m,气候变化复杂,是全球生物多样性的重点区域,具有非常重要的生态意义〔6〕。高黎贡山西坡巨大的垂直落差造成气候、土壤和植被的垂直地带性差异明显〔7〕。高黎贡山坡向的差异使温度、降水等环境因子的变化显著,因此可作为研究坡向对土壤微生物分布格局影响的理想区域。
捕食线虫真菌是一种既可以营腐生生活,也可以以营养菌丝特化形成捕食器官捕食线虫的真菌类群,具备较强的环境适应能力〔8〕。有关捕食线虫真菌的分离、纯化技术和鉴定方法已很成熟。综合以上特点,捕食线虫真菌是作为微生物多样性分布格局研究的理想对象。
因此,本研究拟以滇西北高黎贡山国家自然保护区为研究区域,以捕食线虫真菌为研究对象,研究不同坡向上捕食线虫真菌的分布格局,探究坡向对捕食线虫真菌空间分布格局的影响。
1 材料与方法
1.1 样品采集土样采自贡山县至福贡县的独龙江公路,每1 km设置1个样点,以独龙江隧道为分界线,将高黎贡山北段分为东、西坡,每个采样点对角线5点取样(5~30 cm深土层),充分混匀,取约200 g装于采样袋,带回实验室低温密封保存,并于1周内进行撒样处理。共100个样点,其中东、西坡各50个样点,共500份土样,样点采集的海拔范围在1 400~3 400 m。
1.2 地理、环境指标在采集土样时,记录该区域样品的采集时间、经度、纬度、海拔和坡向等指标。
1.3 培养基的制备玉米琼脂培养基(CMA)和马铃薯葡萄糖琼脂培养基(PDA)参照文献〔8〕进行制备。
1.4 诱饵线虫的制备用贝尔曼氏法制备全齿复活线虫(Panagrellus redivivus)幼虫悬液,0.1 mL约含线虫500条,备用〔8〕。
1.5 分离纯化参照李天飞等〔9〕的诱饵平板法进行分离纯化。
1.6 捕食线虫真菌的鉴定通过菌株形态特征及捕食器官类型〔10〕进行形态学鉴定,同时通过ITS(internal transcribed spacer region of the ribosomal RNA gene,核糖体RNA上的非转录间隔区)和TUB(β-tubulin gene,微管蛋白编码基因)基因序列同源性分析进行分子生物学鉴定〔11-12〕,两者结合进行种属的鉴定。
1.7 数据处理 (1)检出率(occurrence frequency,OF)=(某个种出现的土样数∕总土样数)×100%〔13〕。
(2)总检出率=(检出菌的土样数∕总的土样数)×100%。
(3)物种丰富度:种群中物种数目的多少。物种丰富度往往作为生物多样性的代表性测度,具有简单直观等优点。
运用Excel 2016软件对原始数据进行整理分析。
2 结果
2.1 东、西坡捕食线虫真菌分布的整体差异本研究从500份土样中共分离和鉴定出2属15种120株捕食线虫真菌,其中东坡共检出2属12种46株,西坡共检出2属13种74株。见表1。西坡的物种丰富度和检出率均高于东坡。见图1。

表1 东、西坡捕食线虫真菌检出情况

图1 高黎贡山东、西坡各指数的比较
2.2 东、西坡不同坡位捕食线虫真菌的多样性差异将高黎贡山研究区域分为5个坡位,分别为下坡位(1 400~1 800 m)、中下坡位(1 800~2 200 m)、中坡位(2 200~2 600 m)、中上坡位(2 600~3 000 m)、上坡位(3 000~3 400 m)。对高黎贡山东、西两坡相同坡位的捕食线虫真菌物种丰富度和检出率进行比较。结果表明东、西坡捕食线虫真菌的物种丰富度和检出率随着坡位的升高均呈先上升后下降的趋势,其中东坡各指数均在中下坡位处呈现最大值,西坡物种丰富度在中下坡位处呈现最大值,检出率在中坡位处呈现最大值。见图2~3。

图2 高黎贡山东、西坡相同坡位物种丰富度的比较
2.3 东、西坡捕食线虫真菌优势种和特有种的差异对高黎贡山东、西坡的优势种和特有种进行分析,结果表明A.thaumasium为高黎贡山东、西两坡的优势种,东坡特有种为A.elegans和A.eudermatum,西坡特有种为A.rutgeriense、A.javanica和Dac.drechsleri。见图4。
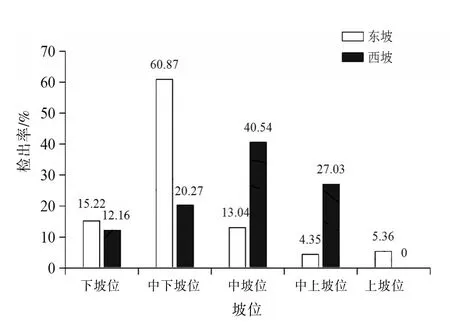
图3 高黎贡山东、西坡相同坡位检出率的比较

图4 高黎贡山东、西坡每个样点菌株检出数
3 讨论
3.1 东、西坡捕食线虫真菌的整体差异高黎贡山东坡共检出2属12种46株捕食线虫真菌,西坡共检出2属13种74株捕食线虫真菌,表明高黎贡山西坡的捕食线虫真菌资源较东坡更为丰富。由于西南和东南季风大气水分输送、流域高山河谷地貌的阻挡因素以及地面水分蒸散的共同作用,形成滇西北三江并流区域特殊的多元立体气候特征:三江并流区域降水量自西向东逐步减少,蒸发量逐步增大〔14〕。高黎贡山西坡属于中亚热带西南季风气候,由于高黎贡山、碧罗雪山山脉均呈南北向排列,当印度洋气流由西南向东北行进时,水气遇到高耸的山脉上升之后逐渐形成降雨,使该区域常年具有丰沛的降雨。高黎贡山西坡小气候十分明显,立体气候尤为突出,一山之内热、温、寒三带气候兼有,雨量特别充沛,年均降雨量达4 000 mm,年平均气温为16℃,为东南亚3个多雨中心之一〔15〕。东坡的气候为半湿润气候,据贡山县气象资料显示,该区域年平均气温为14.7℃,年均降雨量为1 667.4 mm。因此,与坡向相关的温度和降雨量的差异可能是导致西坡捕食线虫真菌物种丰富度、菌株检出数和检出率高于东坡的原因。
3.2 东、西坡相同坡位捕食线虫真菌差异东、西坡捕食线虫真菌的物种丰富度、菌株检出数和检出率随着坡位(海拔)的升高,呈先上升后下降的趋势。Mccain〔16〕发现土壤微生物生物量的周转及其数量随着土壤含水量的提高而有所增加。张萍等〔17〕对高黎贡山土壤微生物分布及生化特性的研究表明高黎贡山土壤的含水量随海拔高度的升高呈先上升后下降的趋势,土壤含水量在中海拔段呈现最大值。相关研究表明高黎贡山土壤微生物的分布格局与生态环境有着密不可分的联系,高黎贡山土壤微生物数量随海拔的下降而增加,中海拔段土壤中物质能量代谢较快,微生物种类较为丰富,高海拔段由于温度偏低的原因,物质能量代谢较为缓慢,微生物数量明显减少,而低海拔段由于人为干扰的影响,植被破坏严重及气候干燥、水分含量较低的原因,使得土壤微生物的数量有所下降〔17〕。因此,土壤含水量和温度作为坡向的间接影响因子可能是高黎贡山捕食线虫真菌物种丰富度和检出率随海拔的升高呈先上升后下降趋势的原因。
3.3 东、西坡捕食线虫真菌优势种和特有种分布高黎贡山北段捕食线虫真菌的优势种群为A.thaumasium,这与杨浩然等〔18〕对云南文山进行捕食线虫真菌多样性的调查结果相一致。其原因可能是Arthrobotrys属捕食线虫真菌的腐生能力较强,对营养要求不严格,在不同生境(水生、陆生、粪生及植物根际、温泉)中,该属捕食线虫真菌资源最为丰富,优势种也都是在该属中出现〔19-23〕。根据Gray〔24〕的调查发现,不同捕器类型的捕食线虫真菌对营养的要求不同,产黏性菌网的种类因生长速度快,对营养需求不严格且腐生能力较强而处于优势。而特有种的差异可能是不同的生态环境所导致,表明产不同捕食器官的捕食线虫真菌对环境的适应能力存在差异,表现为不同的地理区域、不同的生态环境下,该类型真菌的优势种和特有种存在差异。
综上所述,温度和降雨量作为坡向的间接影响因子可能是造成捕食线虫真菌分布存在异质性的主要因素。
