黄山常绿阔叶林优势种甜槠的种群动态与点格局分析
2019-01-04李小东方炎明
李小东,丁 晖,吴 俊,方炎明*
(1 南京林业大学 生物与环境学院/南方现代林业协同创新中心/亚热带森林生物多样性保护国家林业局重点实验室, 南京 210037;2 环境保护部南京环境科学研究所自然保护与生物多样性研究中心/国家环境保护生物安全重点实验室, 南京210042;3 黄山风景区管理委员会园林局, 安徽黄山 245800)
甜槠(Castanopsiseyrei)是壳斗科锥属常绿乔木,主要分布于中国长江以南各地[1],是常绿阔叶林或针阔混交林中的主要树种[2]。甜槠具有特别的解剖学与导水功能特征,与多数锥属树种一样,木材为环孔材或半环孔材[3-4],而环孔材与非等水(anisohydric)气孔调节行为关联[5],在低水势情况下,环孔材具有比散孔材更高的气孔导度[6]。因此,甜槠富有森林水文功能,甜槠林具有增强地表枯枝落叶持水与土壤蓄水能力、减少地表径流量与地下渗流量等水文效应[7]。作为常绿阔叶林的优势树种,甜槠林为动物提供有利生存环境,与动物形成互惠关系。甜槠凋落物支撑了土壤节肢动物群落[8]。在锥属树种的种子扩散与啮齿类取食[9]、传粉服务与昆虫取食方面[10],植物与动物形成了互惠关系。锥属树种具有独特的光合生理特征,最大净光合速率、暗呼吸速率、表观量子效率、光补偿点和光饱和点相对较低[11],不同冠层的光合生理特性存在明显差异,最大净光合速率、暗呼吸速率、光补偿点和光饱和点等指标均以林冠上层较高,中下层较低[12]。因此,在不同环境下,植物可以通过个体发育的差异表达或者表型可塑性表达来实现对局部环境的适应。此外,锥属树种在增加森林碳汇[13]、应对全球气候变化[14]等方面均有重要意义。
黄山拥有保存较好的以甜槠为主的中亚热带常绿阔叶林,甜槠林在黄山地区具有很高的生态价值,具体体现在水源涵养、物种多样性保护、生物量维持、局域性气候调节等方面[15]。然而,黄山森林面临诸多挑战,如生物入侵[16]、生态环境承载力限制[17]、气候变暖[18]自然灾害频发[19]等。气候变暖会加剧病虫害的流行及森林脆弱性,极端天气及自然灾害发生频繁。近年来,黄山甜槠种群面临类似的挑战,甜槠大树枯梢现象时有发生。对于世界文化与自然遗产地常绿阔叶林建群种甜槠,其种群是否存在生存风险这一问题,值得我们特别关注。
本研究采用静态生命表、存活曲线、生存分析、点格局分析等种群生态学方法,借助本项目组前期获得的固定样地数据,探讨甜槠种群的大小结构与数量动态,分析甜槠种群的空间分布,为黄山的森林生物多样性的监测、管理和保护提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
黄山位于安徽省的南部(30°08′N,118°09′E),年平均气温为15.5 ℃,1月平均气温为2.9 ℃,极端最低气温为-10.5 ℃,7月平均气温为27.6 ℃,极端最高气温为37.5 ℃,≥10 ℃的活动积温为4 718.3~5 002.2 ℃,年平均降雨量为1 540 mm,无霜期为226 d。黄山是享誉全球的风景名胜区,常绿阔叶林是其地带性植被。黄山动植物区系丰富,植被发育和保护良好,先后列入联合国教科文组织“世界文化与自然遗产名录”、中国“35个生物多样性保护优先区域”和世界自然保护联盟 “最佳管理自然保护地绿色名录”[20]。
1.2 研究方法
1.2.1数据采集根据黄山中亚热带北缘植被特征,按典型性、稳定性、可操作性的原则,选择自然稳定的典型常绿阔叶林作为监测对象,在坡度相对较小、当前及潜在干扰小的林分中布设样地[21]。2014年,参照CTFS调查技术规范进行样地设置[20],确定设立3块投影形状为正方形的固定样地。个样地基本信息见表1。乔木群落样方设为20 m×20 m的样方,要求乔木群落样方覆盖整个样地。对样地进行每木调查,采集的甜槠数据包括:样方号、标牌编号、中文名、学名、胸径(cm)、坐标x和y(m)、枝下高(m)、高度(m)、冠幅x(m)和冠幅y(m)。
1.2.2种群年龄结构种群年龄结构可以预测种群的生长量,反映种群内大、小个体的相对数量关系,同时也能够反映出种群个体在空间上的配置、数量动态和发展趋势[2]。由于甜槠属保护区内保护植物,禁止砍伐,又因甜槠材质较为坚硬,木芯钻取困难,年轮不易读取,也没有可靠的外部特征可以确定其年龄,因此,本研究根据样地调查所得胸径数据资料,采用较为常用的径级代替龄级的方法来分析黄山甜槠的种群结构。结合甜槠的生物学特性和生长情况,将甜槠种群划分为多个径级,第Ⅰ径级为胸径(DBH)小于等于5 cm、第Ⅱ径级为5 cm < DBH ≤ 10 cm、第Ⅱ径级为10 cm < DBH ≤ 15 cm。之后每级间隔5 cm,以此类推(样地A划分为14个径级、样地B划分为6个径级、样地C划分为8个径级)。以径级代表龄级,统计各龄级株数。

表1 黄山常绿阔叶林优势种甜槠研究区样地基本信息
1.2.3静态生命表与存活曲线生命表结构分析是解释种群变化的前提[22]。黄山甜槠种群的静态生命表是根据各龄级的株数编制而成的,它不仅提供了种群出生率、死亡率等相关参数,而且其各项都是相互关联的,可以通过实测值ax求得,其中ax表示x龄级内出现的个体数,a0表示Ⅰ龄级个体数;lx表示标准化存活数,lx=ax/a0×1 000;dx表示x龄级到x+1龄级间隔期内标准化死亡数,dx=lx-lx+1;qx表示x龄级到x+1龄级间隔期内死亡率,qx=dx/lx;Lx表示x龄级到x+1龄级间隔期间平均存活的个体数,Lx=(lx+lx+1)/2;Tx表示x龄级及其以上各龄级存活的个体总数,Tx=∑Lx;ex表示进入x龄级个体的生命期望寿命,ex=Tx/lx;Sx表示存活率,即x+1期存活个体数与x期存活个体数之比,Sx=lx+1/lx;Kx表示致死力或消失率,即从x到x+1期受到的阻力,Kx=ln(lx)-ln(lx+1)。以龄级为横坐标,以死亡率、消失率、标准化存活数的自然对数(lnlx)为纵坐标,分别绘制死亡率消失率曲线、存活曲线,分析种群动态变化。
1.2.4生存分析为了更好地阐明黄山甜槠种群的生存规律,本研究应用生存率函数S(i)、累计死亡率函数F(i)、死亡密度函数f(ti)和危险率函数λ(ti)4个生存分析函数,来进行种群生存分析。其计算公式为:S(i)=S1×S2×S3,…,Si;F(i)=1-S(i);f(ti)=(Si-1-Si)/hi=Si-1qi/hi;λ(ti)=2(1-Si)/[hi(1+Si)]=f(ti)/S(i)。式中:n为龄级总数;Si为存活率;qi为死亡率;hi为龄级宽度(区间长度);ti为植物种群的生存时间[22]。
1.2.5点格局分析采用Programita软件分析甜槠种群的点格局。点格局分析方法[23]被广泛应用于不同尺度的种群空间分布格局及其种间关系的研究[24-25]。以研究尺度作为横坐标,上下包迹线作为纵坐标绘制函数图像。用种群实际分布数据(点图)计算得到不同尺度下的函数值,若函数值在包迹线以内,则种群呈随机分布;若函数值高于上包迹线,则种群呈集群分布;若函数值低于下包迹线,则种群呈均匀分布[23]。植物种群的空间分布格局常常是物种的种子扩散的局限性、环境的异质性以及种内种间竞争等生物因素与非生物因素综合作用的结果。种群的空间格局分析具有尺度依赖性,不同尺度下往往呈现出不同的分布格局。
2 结果与分析
2.1 年龄结构
3块样地甜槠种群年龄结构如图1所示,其中第Ⅰ龄级个体数占总数的比例最高,分别占到62.43%、61.53%、49.46%。第Ⅱ龄级个体数分别占到16.53%、19.23%、21.8%。第Ⅲ龄级个体数分别占到7.74%、5.76%、11.7%。从第Ⅳ龄级起,个体数占总数的比例均不超过10%。个体数随着龄级的增加逐渐减少。这表明3块样地甜槠种群幼苗众多,幼树储备丰富,具有很强的更新能力,属于稳定型种群。
2.2 静态生命表及存活曲线
3块样地的生命表数据详见表2。样地A死亡率(qx)和消失率(Kx)曲线的变化趋势基本一致,出现3个死亡高峰,分别出现在第1、7和10龄级,其死亡率分别为73.3 %、46.9 %和54.9%。根据龄级分析,种群死亡高峰主要出现在以下3个时期,即幼年时期、近成熟前期和老龄阶段。第1个峰值产生的可能原因是甜槠幼苗个体较小,竞争力不够强,受到环境的强烈筛选,幼苗死亡率较高。第2个峰值是在近成熟前期和成年期,甜槠对水分和光照等资源的需求日益加大,种内竞争增大,出现了死亡高峰。第3个峰值伴随着甜槠种群进入生理死亡阶段而产生。样地B出现2个死亡高峰,分别为第2和第4龄级,其死亡率分别达到70%和33.4%。样地C中,出现3个死亡高峰,分别出现在第1、4和7龄级,其死亡率分别为55.9%、68.0%和50.0%(图2)。
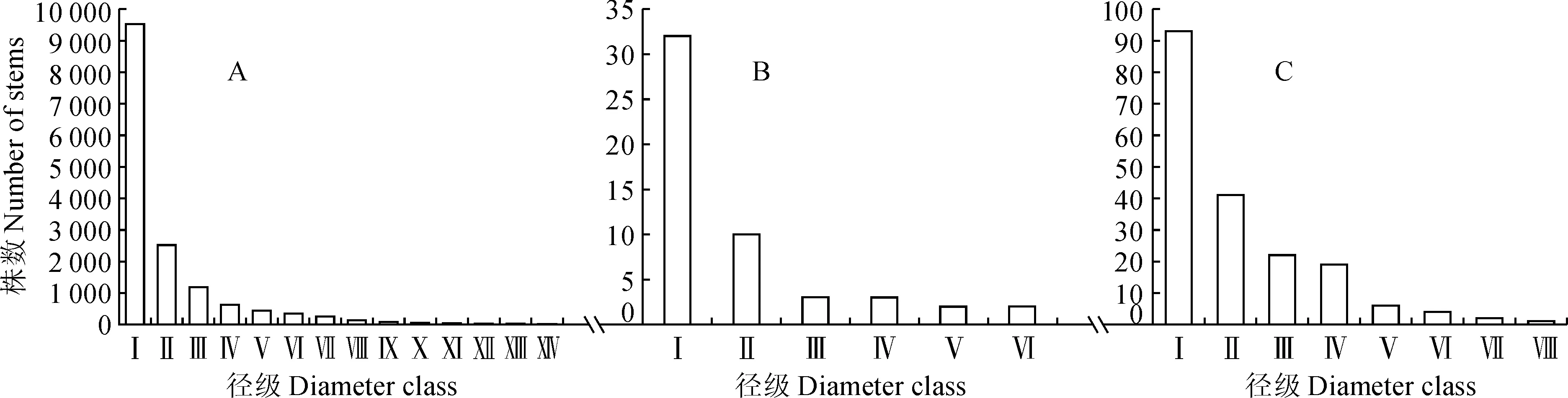
Ⅰ~.径级(代替龄级);DBH.胸径;Ⅰ. DBH≤5 cm;Ⅱ. 5 cm < DBH ≤ 10 cm;Ⅲ. 10 cm < DBH ≤ 15 cm;Ⅳ. 15 cm < DBH ≤ 20 cm;Ⅴ. 20 cm < DBH ≤ 25 cm;Ⅵ. 25 cm < DBH ≤ 30 cm;Ⅶ. 30 cm < DBH ≤ 35 cm;Ⅷ. 35 cm < DBH ≤ 40 cm;Ⅸ. 40 cm < DBH ≤ 45 cm;Ⅹ. 45 cm < DBH ≤ 50 cm;Ⅺ. 50 cm < DBH ≤ 55 cm;Ⅻ. 55 cm < DBH ≤ 60 cm;. 60 cm < DBH ≤ 65 cm;. DBH>65 cm。下同图1 黄山常绿阔叶林优势种甜槠3个样地种群年龄结构Ⅰ~. Diameter class (replacing age class);DBH. Diameter at breast height;Ⅰ. DBH≤5 cm;Ⅱ. 5 cm < DBH ≤ 10 cm;Ⅲ. 10 cm < DBH ≤ 15 cm;Ⅳ. 15 cm < DBH ≤ 20 cm;Ⅴ. 20 cm < DBH ≤ 25 cm;Ⅵ. 25 cm < DBH ≤ 30 cm;Ⅶ. 30 cm < DBH ≤ 35 cm;Ⅷ. 35 cm < DBH ≤ 40 cm;Ⅸ. 40 cm < DBH ≤ 45 cm;Ⅹ. 45 cm < DBH ≤ 50 cm;Ⅺ. 50 cm < DBH ≤ 55 cm;Ⅻ. 55 cm < DBH ≤ 60 cm;. 60 cm < DBH ≤ 65 cm;. DBH>65 cm. The same as belowFig.1 Age structure of dominant species of C. eyrei population in evergreen broad-leaved forests of Mount Huangshan of the three plots

表2 黄山常绿阔叶林优势种甜槠3个样地种群静态生命表
从3块样地的存活曲线(图3)可以看出,在幼苗成长的过程中第一个过渡期(从幼苗过渡到幼树)曲线斜率较大,因而死亡率也较高。这是甜槠生物学特性和环境相互作用造成的结果。甜槠幼苗比较耐荫,但幼树和大树均喜上方光照。随着林分的郁闭,林下光照显著减弱,以光照为主导的环境因子就成为了甜槠幼树存活的限制因子,只有林隙中的幼苗和少量生命力强的幼苗有机会进入下一个生长阶段。随着树高的增加,上方光照得到改善,树高生长加快,开始进入主林层,种间和种内竞争变得更加剧烈,再次出现了死亡的高峰。
Deevey把存活曲线分成了3种类型,Ⅰ型存活曲线呈凸型,表示种群的大多数个体均能生长达到其平均的生理寿命,在达到平均寿命时,几乎同时死亡;Ⅱ型存活曲线呈对角线型,表示各个龄级具有相同的死亡率;Ⅲ型存活曲线呈凹型,表示幼体死亡率较高,以后的死亡率较低而且比较稳定。本研究选用2种数学模型对DeeveyⅡ型和DeeveyⅢ型进行检验,即用指数方程Y=aebx来描述DeeveyⅡ型存活曲线,幂函数Y=axb用以描述DeeveyⅢ型存活曲线,运用SPSS统计软件进行拟合建立相应模型,采用指数模型的F检验及相关指数r值检验。测得3块样地的指数和幂函数种群增长模型分别为:
样地A:Nx=10.543e-0.21x(F=132.292,R2=0.917,P<0.05)
Nx=13.116e-0.997(F=28.981,R2=0.707,P<0.05)
样地B:Nx=7.003e-0.101x(F=22.421,R2=
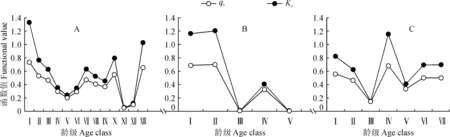
图2 黄山常绿阔叶林优势种甜槠3个样地死亡率与消失率曲线Fig.2 Mortality rate and vanish rate curves of dominant species of C. eyrei population in evergreen broad-leaved forests of Mount Huangshan of the three plots
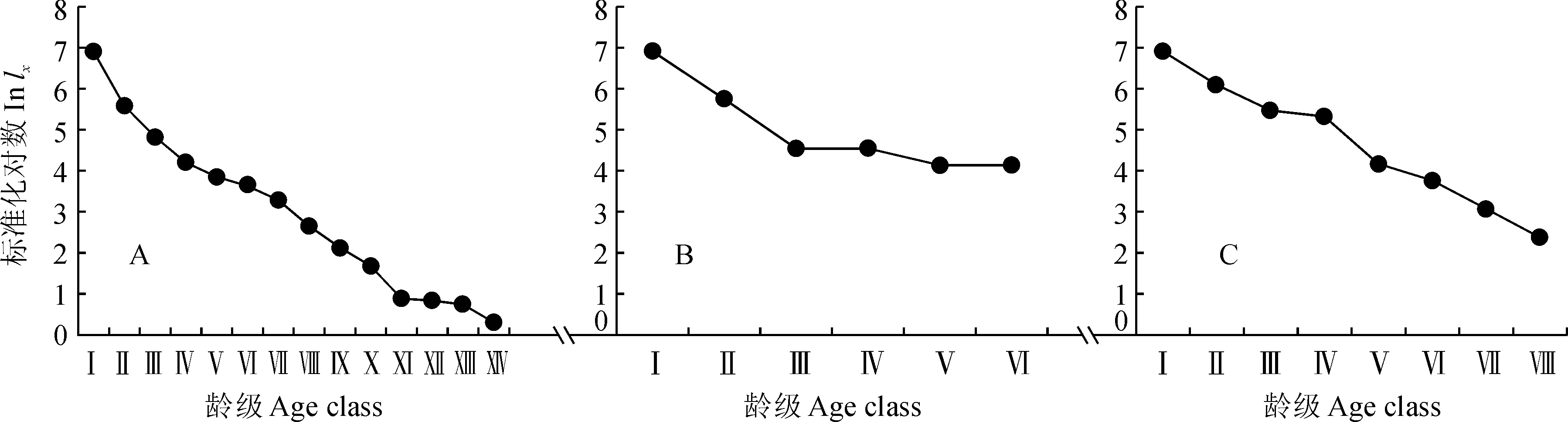
图3 黄山常绿阔叶林优势种甜槠3个样地存活曲线Fig.3 Survival curves of dominant species of C. eyrei population in evergreen broad-leaved forests of Mount Huangshan of the three plots
0.849,P<0.05)
Nx=6.853e-0.304(F=88.662,R2=0.957,P<0.05)
样地C:Nx=8.482e-0.146x(F=156.94,R2=0.963,P<0.05)
Nx=8.168e-0.467(F=26.099,R2=0.813,P<0.05)
A样地和C样地指数模型的相关系数r值大于幂函数模型的相关系数,因此认为,A样地和C样地甜槠种群的存活曲线更趋近于DeeveyⅡ型存活曲线,即表明这2块样地中的甜槠种群各龄级具有相近的死亡率,目前总体上正处于一种动态稳定状态。B样地中幂函数模型的相关系数r值大于指数模型的相关系数,因此认为,B样地中甜槠种群的存活曲线更趋近于Deevey Ⅲ型存活曲线,即表明B样地中的甜槠种群前期死亡率较高,中期和后期死亡率比较低而且稳定,种群目前总体上正处于稳定状态。
2.3 生存分析
由S(i)、F(i)、f(ti)和λ(ti)4个生存函数估算值(表3,图4)可知,3个大样地中甜槠种群均呈现随着龄级增大,种群生存率逐渐减小,种群累积死亡率逐渐增加的趋势。S(i)曲线F(i)曲线均在前面4个龄级变化率较大,从第4龄级之后,F(i)曲线和S(i)曲线的上升和下降幅度均比较平缓,变化率较小。f(ti)曲线在第1龄级和第2龄级变化较大,第2龄级以后,曲线趋于平稳。λ(ti)曲线与qx、Kx曲线呈现出相同的变化趋势。从f(ti)曲线和λ(ti)曲线可以看出:甜槠种群在第1龄级阶段,密度效应最大,种群危险率最高,第1龄级死亡率较高。第6龄级的种群死亡密度不大,但是由于种内和种间竞争逐步加强,所以导致种群危险率较高。A样地中的甜槠种群表现出前期锐减,中、后期比较稳定,末期衰退的特点,这与种群的死亡率函数qx曲线、消失率函数Kx曲线和Deevey 存活曲线的分析结果相似。
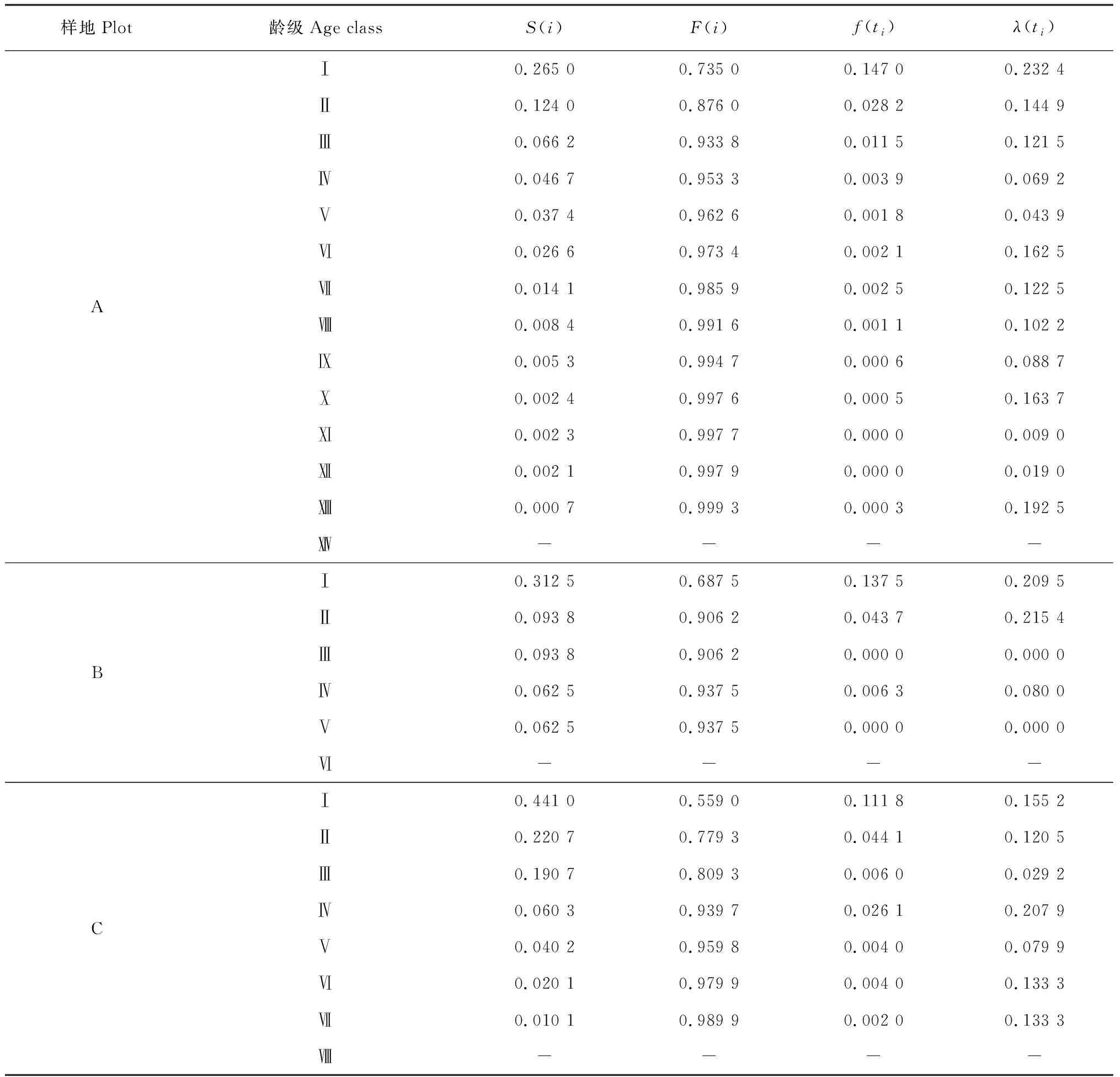
表3 黄山常绿阔叶林优势种甜槠3个样地种群生存函数值
2.4 点格局分析
根据树木个体的坐标值用实测值,得到各样地甜槠种群空间分布点图(图5)。在A、B和C样地中,甜槠个体分别为15 246株、52株和188株。
在A样地中,在尺度小于150 m时,甜槠种群表现为显著的集群分布;大于150 m,接近于随机分布。在B样地中,小于20 m,为显著的集群分布;20~30 m,为显著的随机分布,30~46 m,为显著的均匀分布;大于46 m,显著的随机分布。在C样地中,小于13 m,为显著的集群分布;13~25 m,为显著的随机分布;25~37 m,为显著的均匀分布;大于37 m,为显著的随机分布(图6)。
综合以上结果,甜槠种群在小尺度上呈显著的聚集性,随着尺度的增加则表现为随机分布。由于样地内生境条件比较均一,外界因子对种群分布格局的影响不是很显著,所以,甜槠种群的分布主要与甜槠生物学特征有关。甜槠种群果实比较重,含有淀粉,可以靠动物传播种子也可以借助重力传播。但是传播距离是有限的,基本散落在母树周围。从种群在不同发育阶段的分布特征,可以看出种群的适应机制和生存策略。幼年期种群集聚度大,具有明显的群体效应,小龄级个体生长需要的资源较少,种内竞争较弱,聚集型分布也有利于幼苗、幼树的存活。成年植株个体增大,集聚程度有所降低,有利于充分的获取环境资源,维持个体的正常生长和群体的稳定性。长成大树后不考虑生境的异质性,则呈现随机分布状态。

图4 黄山常绿阔叶林优势种甜槠3个样地生存分析曲线Fig.4 Survival function curve of dominant species of C. eyrei population in evergreen broad-leaved forests of Mount Huangshan of the three plots
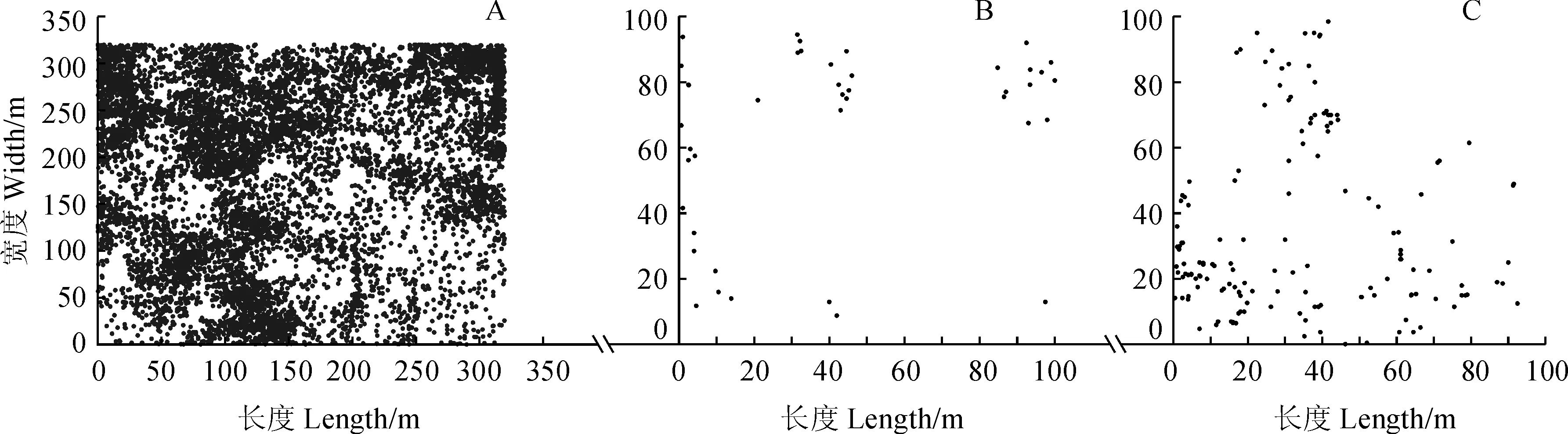
图5 黄山常绿阔叶林优势种甜槠3个样地的空间分布点图Fig.5 Graph of spatial distribution of dominant species of C. eyrei population in evergreen broad-leaved forests of Mount Huangshan of the three plots

图6 黄山常绿阔叶林优势种甜槠3个样地的空间分布格局Fig.6 Distribution pattern of dominant species of Castanopsis eyrei population in evergreen broad-leaved forests of Mount Huangshan of the three plots
3 讨 论
本研究以黄山森林植物多样性监测大样地内的典型甜槠种群为研究对象,采用多种手段对样地内的甜槠种群进行调查与分析。经调查黄山样地内的甜槠种群密度为1 280株/hm2,这均多于[26-29]浙江古田山514株/hm2、浙江天台山421株/hm2、福建武夷山144株/hm2和大安源685株/hm2。经查文献可知其样地分布均在海拔500 m以上,而黄山样地分布下限是400 m。由此可见,黄山的甜槠林非常典型。
通过对年龄结构,静态生命表,存活曲线和种群增长模型的分析,表明A、B、C样地的甜槠种群均处于增长阶段,总共出现了3个死亡高峰,分别对应幼年期、近成熟前期和老龄期。幼苗和幼树数量占总体比例最高,随着龄级的增加,个体数逐渐减少。A样地和C样地甜槠种群趋近于DeeveyⅡ型存活曲线,各龄级具有相近的死亡率。B样地甜槠种群趋近于Deevey Ⅲ型存活曲线,前期死亡率较高,中期和后期死亡率比较低而且稳定,属于增长型种群。研究结果尽管与颜玉娟等[2]的研究结果不同,但与多数研究结果相吻合[26,30-31],总体而言,种群呈现增长趋势。
点格局分析表明,甜槠种群呈现小尺度的聚集性与大尺度的随机性。本研究结果与徐学红等[31]的研究结果相吻合,但是与李立、胡砚秋、杨云方、巩合德、Kamei等的研究结果不同[26,28,30,32-33]。这可能与研究所在样地的地形分布影响有关,生境的异质性对种群分布影响比较大,相比而言本研究结果所在样地较平坦,生境基本相同,所以才导致大尺度下呈现随机分布。
A、B、C3块样地中的甜槠种群在年龄结构、死亡率、消失率、存活曲线、生存分析和种群增长方面的趋势基本一致,说明黄山的甜槠种群生存状况良好。而在点格局分析中存在差异,这主要因为3块样地处于不同的海拔高度,且样地面积大小不同,导致3块样地中甜槠种群个体数目存在差异,并且导致个体分布格局在不同的尺度下呈现不同的分布特点。
甜槠作为黄山常绿阔叶林的优势树种,正处于稳定增长的状态,不存在生存风险。这对黄山整个生态系统有重要意义,能有效地增强地表枯枝落叶持水与土壤蓄水能力、减少地表径流量与地下渗流量,又为土壤节肢动物、啮齿类、鸟类及昆虫提供必要的生存环境,同时也能调节气候变化和增加森林碳汇。这也为黄山的旅游业注入新的活力,创造出更多的发展机会。
