柔毛淫羊藿及其近缘种叶片非腺毛形态及其分类学意义
2019-01-04植汉成郭宝林冯学锋郭佳琪温春梅
植汉成,刘 翔,郭宝林,冯学锋,葛 菲,郭佳琪,温春梅
(1 中国医学科学院 北京协和医学院 药用植物研究所,北京 100193;2 江西中医药大学,南昌 330006;3 重庆市中药研究院,重庆 400065;4 中国中医科学院 中药研究所,北京 100700)
柔毛淫羊藿(EpimediumpubescensMaxim.)为小檗科淫羊藿属植物,1887年发表,以叶背面密被柔毛为特征,是中国淫羊藿属植物中分布较为广泛的物种,集中分布于四川、重庆和陕西南部,也分布于甘肃东南角[1],柔毛淫羊藿也是常用中药淫羊藿的主要物种来源。其近缘种星花淫羊藿(E.stellulatumStearn)为1993年发表的物种,与柔毛淫羊藿形态近似,但区别在于花的内轮萼片比较大,小叶片为较宽的卵形[2],特别是叶中几乎不含有淫羊藿苷类成分[3],分布于湖北十堰、陕西商洛。本课题组还发现分布于陕西镇巴的一个类型,花形态极似柔毛淫羊藿,但小叶片小而卵圆,根状茎细长横走,暂定名为镇巴淫羊藿(E.zhenbaensesp.nov.),以及分布于四川万源的一个类型,花形态和小叶片形态似柔毛淫羊藿,但内萼片粉红色,叶背面为短伏毛,暂定名为万源淫羊藿(E.wanyuanensesp.nov.)。
非腺毛是由植物表皮细胞分化而成、不具分泌功能的突起物,它广泛存在于植物叶、茎、花及果实的表皮上,由单细胞或多细胞构成,无头部和柄部之分,顶端通常狭尖[4]。大部分淫羊藿属植物叶背被非腺毛,研究表明淫羊藿属植物非腺毛的特征对该属植物分类、鉴别很有价值[5]。梁海锐等最早对淫羊藿属植物的非腺毛研究发现,该属植物不同物种非腺毛形态具有专属性[6]。何顺志等也印证了该观点[7-8]。本课题组前期对淫羊藿属植物非腺毛进行研究,发现不同物种叶背非腺毛的形态、顶细胞的长度、直径、伸长细胞的有无、基部细胞数目、顶细胞与相邻细胞的衔接方式等具有特征性,总结了淫羊藿属非腺毛可以分为5种主要类型,并推测了非腺毛特征的演化趋势[9]。课题组又针对柔毛淫羊藿14个居群及其近缘种星花淫羊藿1个居群的非腺毛和ISSR分子标记开展了研究,依据每个居群具有的非腺毛主流类型将非腺毛分为5类——长直毛、长卷曲毛、伏曲毛、拟短伏毛和长伏毛,并初步推测了长直毛-长卷曲毛-伏曲毛的演化趋势[10]。
我们继续研究发现,柔毛淫羊藿叶背面的非腺毛是多种类型共存的,且类型间存在连续的变化关系,因此本研究基于原来的基础,增加了更多分布区域的居群样本,并将与柔毛淫羊藿相关的镇巴淫羊藿(1个居群)和万源淫羊藿(1个居群)纳入研究,通过观察每个居群所具有的各类非腺毛形态,以期进一步阐明柔毛淫羊藿及近缘种所具有的非腺毛的形态特征及演化关系,并据此提供柔毛淫羊藿和几个近缘种亲缘关系的证据。
1 材料和方法
1.1 实验材料
柔毛淫羊藿31个居群,星花淫羊藿3个居群,镇巴淫羊藿1个居群和万源淫羊藿1个居群,所有居群的分布点参见表1。每个居群采集10片以上的成年叶,硅胶快速干燥。
1.2 实验仪器和试剂
OLYMPUS BX51型光学显微镜,DP70型CCD成像系统,无水乙醇、冰醋酸、甘油、H2O2、甲醛均为分析纯试剂。
1.3 实验方法
1.3.1制片与观察方法(1)叶表面片制备(刮去叶肉离析法):将干燥叶放清水中泡至软化后转入FAA固定液中保存。取FAA浸泡叶片,在中部近中脉处切取约3 mm×6 mm 的小块,刮去上表皮及叶肉组织,放入盛离析液(30%H2O2∶冰醋酸=1∶1)的烧杯中,60 ℃烘箱加热约1~2 h,小叶块透明后,醋酸甘油试液(50%冰醋酸∶甘油∶蒸馏水=1∶1∶1)封片。

表1 柔毛淫羊藿及近缘种样品信息
(2)非腺毛片制备(刮取茸毛法,适用于非腺毛密被,表面片不易观察的情况):取干燥叶,在叶背中部近中脉处刮取非腺毛,置于载玻片上,醋酸甘油试液封片。
(3)观察和记录:光学显微镜观察和拍照,并测量和记录非腺毛特征(每个居群均取5枚叶片观察)。
(4)图片处理:用Photoshop软件排版处理。
1.3.2数据测量统计与处理选取放大200倍非腺毛照片,先划分非腺毛形态类型,然后每一类型取20根非腺毛,记录顶细胞形状、顶细胞与相邻细胞衔接方式、基部细胞数和伸长细胞数,用Digimizer软件测量非腺毛全长、顶细胞长度、顶细胞(中间)直径和壁厚。选取放大100倍照片的1 μm2区域用Digimizer软件统计非腺毛数量,共10个重复(每个居群取5枚叶片,每片2个区域),计算得出单位面积非腺毛个数(个/mm2)。
2 观察结果
2.1 柔毛淫羊藿及近缘种的非腺毛类型
柔毛淫羊藿及近缘种的多细胞非腺毛形态和结构与以前研究结果一致[10],由基部细胞(指非腺毛与表皮细胞相连接的薄壁细胞,一般扁小)和顶细胞(指末端的一个细胞,壁厚,顶端尖或偶钝)构成,部分多细胞毛有伸长细胞(在基部细胞和顶细胞之间,壁厚,一般比基部细胞长);顶细胞是柔毛淫羊藿及近缘种非腺毛的主要部分,大部分居群顶细胞长度占非腺毛总长90%以上;顶细胞特征变化(顶细胞形状、顶细胞长、顶细胞占的比例、顶细胞直径)、基部细胞数、伸长细胞有无及数量,以及顶细胞与相邻细胞衔接方式存在多样性。本研究在原来发现的5类非腺毛(长直毛、长卷曲毛、伏曲毛、拟短伏毛和长伏毛)的基础上,新发现短伏毛和单细胞毛等两类新的非腺毛,短伏毛基部细胞扁小,无伸长细胞,顶细胞一般梭形,顶细胞与基部细胞成一夹角,与箭叶淫羊藿[Epimediumsagittatum(Sieb. et Zucc.) Maxim.]中报道的非腺毛形态相似[11],仅在万源淫羊藿中发现;单细胞毛仅由1个细胞构成,形状平直或略弯曲,呈剑形或梭形,在25个居群中发现,这类毛在淫羊藿属非腺毛中为首次发现。在详细观察和测量了各类非腺毛的形态和大小,又将直毛、曲毛、长伏毛区分出亚类,各类毛形态参见图版Ⅰ。
2.1.1直毛(图版Ⅰ,A1~A6) 长约50~900 μm,整个非腺毛平直,基部细胞一般短小(部分亚类向上有细胞较为伸长),偶有伸长细胞,顶细胞与其他细胞直接连接,根据非腺毛长度、基部细胞数和顶细胞形状,又细分为6个亚类(A1~A4为长直毛,长度常大于300 μm,顶细胞常呈剑形;A5~A6为短直毛,长度常小于300 μm,顶细胞常呈梭形),其中A1类有1~2个伸长细胞,其他亚类无伸长细胞。A4类为最常见类型直毛。具体特征为:A1、长约300~500 μm,基部细胞3~9个,短小或向上有细胞略伸长,伸长细胞1~2个;A2、长约300~500 μm,基部细胞3~9个;短小或向上有细胞略伸长;A3、长约300~600 μm,基部细胞4~7个,短小; A4、长约300~900 μm,基部细胞1~4个,短小或扁小;A5、长约200~300 μm,基部细胞1~3个,短小或扁小;A6、长约50~200 μm,基部细胞1~2个,扁小。
2.1.2曲毛(图版Ⅰ,B1~B5) 长约150~1 500 μm,顶细胞波状弯曲或卷曲状,基部细胞一般短小(部分亚类向上有细胞较为伸长),偶有伸长细胞;顶细胞与相邻细胞直接连接,偶钝角连接,根据非腺毛长度、基部细胞数、有无伸长细胞和顶细胞卷曲程度,又细分为5个亚类(B1~B4为弯曲毛,顶细胞波状弯曲;B5为长卷曲毛,顶细胞长卷曲状),其中B1类有1~3个伸长细胞,其他亚类无伸长细胞。另外,B5有的顶细胞与基部细胞钝角连接。B4、B5类为最常见类型曲毛。具体特征为:B1、长约250~500 μm,基部细胞4~10个,短小或向上有细胞略伸长,伸长细胞 1~3个,偶4~5个; B2、长约250~500 μm,基部细胞1~4个,短小或向上有细胞略伸长;B3、长约250~500 μm,基部细胞3~6个,短小;B4、长约150~700 μm,基部细胞1~3个,短小;B5、长约300~1500 μm,基部细胞1~3个,短小。
2.1.3伏曲毛(图版Ⅰ,C) 长约520~1 100 μm,基部细胞1~3个,顶细胞波状弯曲或卷曲状,顶细胞与相邻的基部细胞偏斜T型/T型(呈直角)连接。
2.1.4长伏毛(图版Ⅰ,D1a~D2b) 约230~630 μm,基部细胞1~15个,细胞由基部向上渐长,顶细胞平直或弯曲,顶细胞与相邻细胞直角或钝角连接,有伸长细胞的为D1类(又细分为顶细胞平直为D1a类,弯曲为D1b类),无伸长细胞为D2类(又细分为顶细胞平直为D2a类,弯曲为D2b类);D1a和D2a称为长伏直毛,D1b和D2b称为长伏曲毛。其中D2a为常见类型。
2.1.5拟短伏毛(图版Ⅰ,E) 约140~250 μm,基部细胞1~2个,扁小,顶细胞与基部细胞直接连接,但二者衔接处,顶细胞呈直角弯曲而形成假T型。
2.1.6短伏毛(图版Ⅰ,F) 约170~280 μm,基部细胞1~4个,扁小,顶细胞与基部细胞直角或钝角连接,顶细胞平直呈剑形或梭形。
2.1.7单细胞毛(图版Ⅰ,G) 由一个细胞组成,形状平直、弯曲或卷曲,呈剑形或梭形。
本研究还观察到有13个居群叶背存在乳突,乳突结构不同于非腺毛,是表皮细胞的细胞壁向外凸起形成,呈球形乳突,在淫羊藿属其他植物中也有发现[12-13]。
2.2 柔毛淫羊藿及近缘种非腺毛演化分析
基于在柔毛淫羊藿及近缘种中发现的非腺毛形态进行演化分析,依据以下原则:
①普遍存在性状为原始特征:长直毛和弯曲毛为柔毛淫羊藿及近缘种普遍存在,可以认为是原始的特征,因此演化关系为长直毛在先,短直毛在后,弯曲毛在先,长卷曲状毛在后。
②与原始类群相关为原始特征:淫羊藿属的原始类群的毛为多细胞长毛,顶细胞和基部细胞为直接相接,因此基部细胞多而伸长,以及具有伸长细胞的非腺毛为原始特征;不具伸长细胞,基部细胞变短变少为演化特征。
③特化特征为次生特征:非腺毛顶细胞膨大、细胞壁格外加厚者,顶细胞异常弯曲、顶部细胞与相邻细胞出现夹角以及呈现T型/偏斜T型连接均为演化特征。
根据以上演化推断,以及同一个居群(同一片叶片)有多种类型而不同居群常有共同毛类型的现象。我们推断出柔毛淫羊藿及近缘种非腺毛的演化顺序见图1(其中实线推断可能性较大)。
2.3 各个居群非腺毛类型构成及形态特征
36个居群叶片背面非腺毛密度、类型构成,以及每个类型的非腺毛全长、顶细胞长、顶细胞直径、顶细胞壁厚、基部细胞数、伸长细胞数、顶细胞和下部细胞连接方式等非腺毛形态学数据汇总于表2。由表2可知,21个居群以直毛A类为主要类型,占所有居群的58%,以曲毛B类为主要类型的有7个居群,以长伏毛D类为主要类型有6个居群,以拟短伏毛E类为主要类型的只有1个居群,以短伏毛F类为主要类型的也只有1个居群,伏曲毛C类只存在于一个以B类为主要类型的居群中。因所有居群均有直毛A类型非腺毛,说明直毛A类为柔毛淫羊藿及近缘种的核心类型非腺毛。而E类和F类各自只存在于一个居群中,应属于特化类型。
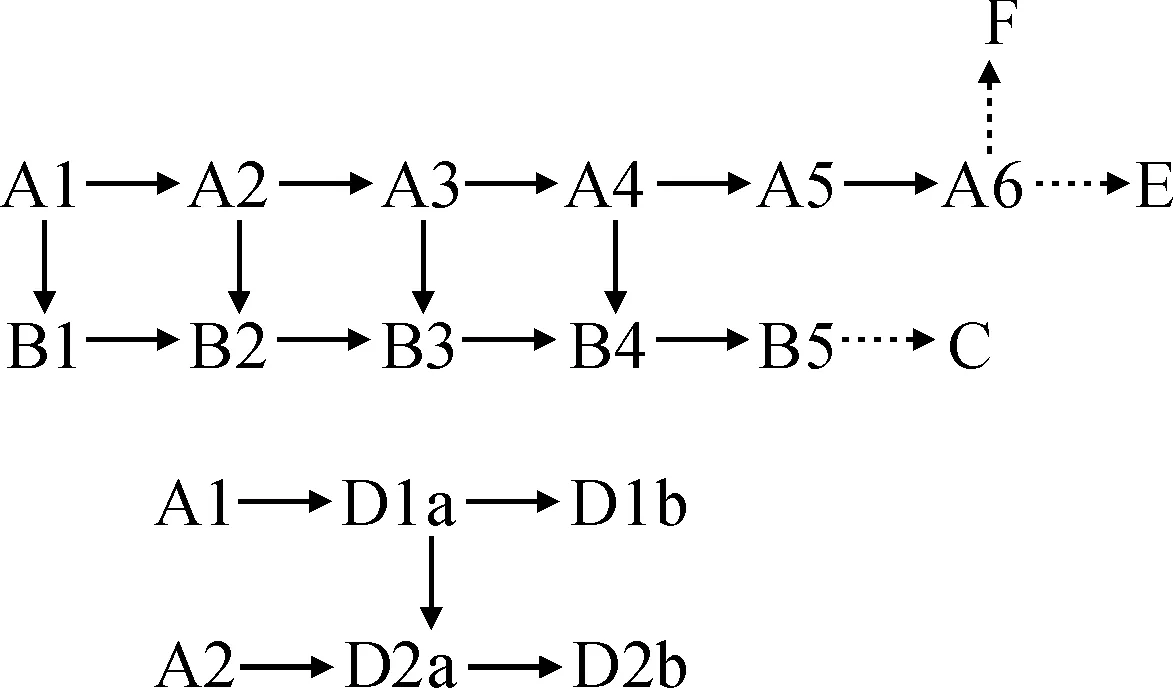
图1 柔毛淫羊藿及近缘种非腺毛演化关系Fig.1 The evolution relationship of non-glandular hairs in E. pubescens and related species
依据表2中各个居群的非腺毛类型构成,进行居群归类,归类原则为:以A类直毛为主的居群,编为直毛群A群,以B类曲毛为主的居群,编为曲毛群B群, 以D类长伏毛为主的类群编为D群,以E类拟短伏毛为主的居群编为E0群,以F类短伏毛为主的居群编为F0群。再依具体居群非腺毛类型构成,分为AⅠ、AⅡ、AⅢ群等,如AⅠ群以A4型毛为主要类型毛,同时存在A3、B4类型毛。柔毛淫羊藿及近缘种各居群按照非腺毛的分类见表3。其中以AⅡ群为最主要居群类型,包括11个居群;其次是AⅠ、BⅢ和DⅢ群,分别包括4个居群;再次是AⅢ群,包括3个居群;AⅣ群,包括2个居群;其余则均只有1个居群。
2.4 柔毛淫羊藿及近缘种各居群非腺毛关联与演化分析
依据表3的居群非腺毛类型划分,我们给出了不同类型居群的演化路线(图2):
1)直毛群:AⅠ→AⅡ→AⅢ→AⅣ→AⅤ。AⅡ群为主要类型,AⅠ与AⅡ群不同仅在于,AⅠ群存在有较为原始的A3类非腺毛。2)曲毛群:BⅠ→BⅡ→BⅢ→BⅣ。BⅢ群为主要类型,BⅠ和BⅡ群均是以较为原始的B4类毛为主,而BⅠ群则又存在更为原始的B3类(及A3类)曲毛。3)长伏毛群:DⅠ→DⅡ→DⅢ,DⅢ群为主要类型,DⅠ和DⅡ群以较为原始的D1a类毛为主,DⅠ更是不存在D2a类毛。4)直毛群-曲毛群:直毛群中具有少量曲毛中的弯曲毛,曲毛群中具有少量直毛类毛,从共有性上推测存在AⅠ→BⅠ,AⅡ→BⅡ或AⅡ→BⅢ或AⅡ→BⅣ的关系。5)短直毛-短伏毛和拟短伏毛特化群: AⅤ→E0,AⅤ→F0,E0群、F0群与AⅤ群发生联系,是他们均具有AⅤ的主流毛类型A5和A6。
A1、A2、B1、B2类非腺毛并未在任何一个居群中作为主要类别的非腺毛存在,也未存在于直毛群和曲毛群中,而在长伏毛群D群中为次要类型毛。此外,在AⅠ群中,非腺毛A3与A2较相似,无伸长细胞,基部细胞较多,只是A3基部细胞略短小;在BⅠ群中,非腺毛B3与B2也较相似。我们认为这4种非腺毛可能与直毛群、曲毛群和伏毛群是有联系的。





表3 基于非腺毛类型对柔毛淫羊藿及近缘种36个居群分类表
注:具下划线的非腺毛类型为该居群的主要类型
Not: The underlined types represent the main type of non-glandular hair in this population

图2 柔毛淫羊藿及近缘种基于非腺毛形态的居群演化关系图Fig.2 Population-relation graph of E. pubescens and related species based on non-glandular types
2.5 柔毛淫羊藿及近缘种非腺毛形态与地理分布相关性分析
将各个居群按照非腺毛构成定义的名称填入地理分布图中(图3),由图3可看出,直毛群A分布在柔毛淫羊藿的核心区域,其中较为演化的AⅣ和 AⅤ分布在较外区域;曲毛群B则在核心区外围的北部、西部和南部,也可证实直毛群-曲毛群的演化关系;长伏毛群D分布为离核心区和曲毛群分布点较远的另一个区域,与直毛群可能存在一定的联系,在长伏毛群内,由于样本比较少,DⅠ~DⅢ 的演化关系不甚明朗,有待进一步研究。拟短伏毛群E0分布于直毛群中,可能为AⅤ直接演化而来;而短伏毛群F0形态离直毛群AⅤ的关系较近,但地理上与直毛群核心区域较远,因此还需要进一步研究。
顶细胞和相邻细胞的连接方式导致直接连接的直毛、曲毛和拟短伏毛,与直角/钝角连接的长伏毛和短伏毛两个大的类型群之间,在地理上是能区分的,其中陕西南郑居群SXNZ(DⅢ)和四川南江居群SCNJ(AⅡ)为两个类型的分界居群(均属于柔毛淫羊藿),值得进一步研究两类非腺毛毛分化的产生。具有BⅣ型非腺毛构成的只有一个居群SCHY,与距离很近的SCYA居群的非腺毛构成,多了C类伏曲毛(区别于B类毛特征在于其顶细胞和相邻细胞的连接方式为T型/偏斜T型连接),而连接方式在柔毛淫羊藿及近缘种的各居群中是不连续分布的,因此SCHY居群也有待研究。




图3 36个居群非腺毛形态与地理分布相关性Fig.3 Correlation between non-glandular hair characteristics and geographical distribution of 36 populations
3 讨 论
3.1 柔毛淫羊藿及其近缘种分类学关系
刘翔等基于花、花序、主要非腺毛类型和ISSR分析结果建议将星花淫羊藿处理为柔毛淫羊藿种下等级。本文基于居群多细胞非腺毛构成以及地理分布,柔毛淫羊藿及近缘种各居群的关系得到了进一步的阐明,其中星花淫羊藿的3个居群均为长伏毛群的DⅢ群,地理上位于东北侧,但是与柔毛淫羊藿的陕西山阳居群形态地理上均具有联系性,同意刘翔等观点[10]。镇巴淫羊藿和万源淫羊藿地理上相距很近,但居群毛的构成上,前者为DⅠ群,后者为F0群(二者虽然属于顶细胞和相邻细胞的连接方式上为T型/偏斜T型连接,但基部细胞组成和形态差异很大),周围无A群和其他D群(F0和AⅤ虽然形态上有联系,但地理上也没有连接点)相关的联系居群样本,因此需要增加邻近地区的其他居群(特别是属于柔毛淫羊藿的居群)的研究才能进一步阐明他们和柔毛淫羊藿的关系。
3.2 单细胞毛和乳突的分类学价值探讨
本研究观察到柔毛淫羊藿及其近缘种共27个居群中具有单细胞毛,占本研究中居群总数的75%,直毛群(A群)21个居群中除SCCZ、SCJY1、SCQL2、SCDY 4个居群外,81.0%具有单细胞毛;曲毛群(B群)7个居群中仅SCPW3居群未看到单细胞毛,85.7%居群具有单细胞毛;而长伏毛群(D群)6个居群中有4个居群(SXZB、SXSY、HBSY、SXSN)无单细胞毛,仅33.3%居群具有单细胞毛,拟短伏毛群(E0群)和短伏毛群(F0群)各1个居群均有单细胞毛,说明单细胞毛主要存在直毛群、曲毛群、拟短伏毛群和短伏毛群中。以不同种类淫羊藿单细胞毛居群分析发现,柔毛淫羊藿80.6%居群具有单细胞毛,星花淫羊藿仅33.3%的居群具有单细胞毛,万源淫羊藿具有单细胞毛,镇巴淫羊藿无单细胞毛。因此,我们认为单细胞毛可能与主流非腺毛类型和地理分布有关,与种类是否有关还需进一步研究。
自梁海锐等[14]1992年首次发现淫羊藿叶背表皮细胞壁突起以来,王悦云等[13]、张晓东[15]和康帅等[12]相继发现20个物种,分别是湖南淫羊藿(Epimediumhunanense)、茂汶淫羊藿(E.platypetalum)、川鄂淫羊藿(E.fargesii)、粗毛淫羊藿(E.acuminatum)、箭叶淫羊藿(E.sagittatum)、黔岭淫羊藿(E.leptorrhizum)、宝兴淫羊藿(E.davidii)、星花淫羊藿(E.stellulatum)、长蕊淫羊藿(E.dolichostemon)、天平山淫羊藿(E.myrianthum)、水城淫羊藿(E.shuichengense)、小叶淫羊藿(E.parvifolium)、贵州淫羊藿(E.sagittatumvar.guizhouense)、偏斜淫羊藿(E.truncatum)、朝鲜淫羊藿(E.koreanum)、单叶淫羊藿(E.simplicifolium)、竹山淫羊藿(E.zhushanense)、绿药淫羊藿(E.chlorandrum)、四川淫羊藿(E.sutchuenense)、巫山淫羊藿(E.wushanense)存在乳突。柔毛淫羊藿(E.pubescens)、淫羊藿(E.brevicornu)、黔北淫羊藿(E.borealiguizhouense)、毡毛淫羊藿(E.coactum)、德务淫羊藿(E.dewuense)、无距淫羊藿(E.ecalcaratum)、短茎淫羊藿(E.brachyrrhizum)、膜叶淫羊藿(E.membranaceum)、腺毛淫羊藿(E.glandulosopilosum)、少花淫羊藿(E.pauciflorum)、木鱼坪淫羊藿(E.franchetii)、恩施淫羊藿(E.enshiense)12个种叶背无乳突,但本研究通过对36个居群柔毛淫羊藿及近缘种叶背特征研究发现13个居群(SCBZ、SCPW1、SCPW2、SCPW3、SCNJ、SXSY、SCLZ2、SCQC、HBYX、SXSN、SXZB、SCWY、SCJY1)具有乳突。对13个居群进一步分析发现,非腺毛较少(非腺毛密度少于50个/mm2)的居群69.2%具有乳突,如SXSY(非腺毛密度为20个/mm2)、SCNJ(42)、SCQC(49)、SCPW1(40)、SCLZ2(29)、SXSN(36)、SXZB(3)、SCWY(9)、SCJY1(21)居群,另外4个居群SCBZ(60)、SCPW2(55)、SCPW3(77)、HBYX(65)非腺毛较多,密度大于50个/mm2,具有乳突,占非腺毛多居群的17.4%,也说明了非腺毛密度与乳突间可能存在负相关性。特别是以长卷曲毛(BⅢ群、BⅣ群)为主的SCYA(113)、SCHY(125)、SCLC(149)、SCMC(115)、GSCX(107)5居群非腺毛最多,密度达100个/mm2以上,叶背均不具乳突;以不同种类分析柔毛淫羊藿29.3%居群、星花淫羊藿66.7%居群、镇巴淫羊藿和万源淫羊藿具有乳突。因此,我们认为乳突与非腺毛密度可能具有一定的关系,具体仍需对淫羊藿属的其他类群进行研究。
通过对柔毛淫羊藿及近缘种36个居群非腺毛研究发现,柔毛淫羊藿及近缘种非腺毛形态结构特征、有无单细胞、有无乳突,并不是完全一致的,但分析表明居群间整体呈现连续变异,所有居群均有直毛A类型非腺毛,说明直毛A类为柔毛淫羊藿及近缘种的核心类型非腺毛。由此可见, 非腺毛性状至少在某些种内变异十分复杂, 不同产地/居群间存在不同程度的共性和个性类别。所以在以后利用非腺毛进行分类和演化分析时,应注意需要有足够的样本数量,特别是不同居群的样本量,除了进行形态的观察和主要类型的归类外,还应该总结出共性和个性类别。
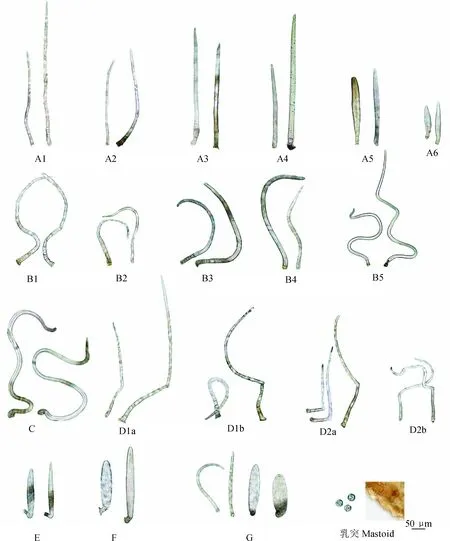
图版Ⅰ 柔毛淫羊藿及其近缘种非腺毛类型及乳突
A1~A4.长直毛;A5~A6.短直毛;B1~B4.弯曲毛;B5.长卷曲毛;C.伏曲毛;D1a、D2a.长伏直毛;D1b、D2b.长伏曲毛;E.拟短伏毛;F.短伏毛;G.单细胞毛
Plate Ⅰ Different types of non-glandular hairs and mastoid inE.pubescensand its related species
A1-A4. Long straight hair; A5-A6. Short straight hair; B1-B4. Curved hair; B5. Long curly hair; C. Appressed curly hair; D1a and D2a. Long appressed straight hair; D1b and D2b. Long curly appressed hair; E. Pseudo short appressed hair; F. Short appressed hair; G. Single cell hair
