龙眼ERF家族成员鉴定及其在体胚发生早期的表达
2019-01-04吕科良高玉莹林玉玲赖钟雄
陈 燕,吕科良,厉 雪,高玉莹,林玉玲,赖钟雄
(福建农林大学 园艺植物生物工程研究所,福州350002)
乙烯响应因子(ethylene response factors,ERF)家族是植物特异性转录因子APETALA2/ethylene responsive factor (AP2/ERF)超家族中的亚家族。在AP2/ERF转录因子超家族中,家族成员都具有AP2/ERF保守结构域。根据AP2/ ERF结构域的数量,亚家族可分为3类:仅包含1个AP2/ERF结构域的ERF家族;含有2个重复AP2/ERF结构域的AP2家族;含有1个AP2/ERF结构域和B3结构域的RAV家族[1-2]。从1994年在拟南芥中分离的第1个与花发育相关的AP2基因[3]开始,涌现了大量对AP2/ERF转录因子全基因组家族分析,其中包括:玉米[4]、粟[5]、甜橙[6]以及葡萄[7]等。ERF家族作为AP2/ERF转录因子超家族中的重要一员,已经有大量的深入研究对ERF家族基因功能进行验证。研究表明,ERF能够在各种生物胁迫与非生物胁迫下响应,包括病害[8]、干旱[9]、盐胁迫[10]以及通过乙烯和ABA信号通路介导各种生理过程[11]等发挥作用。除此之外,ERF在植物体胚发生过程中的调控作用也做了相关研究。Mantiri等[12]在蒺藜苜蓿体胚发生过程中,发现MtSERF1在球形胚以及心型胚分生组织中高表达;Piyatrakul等[13]首次揭示了AP2/ERF基因在体细胞胚胎发生过程中具有调节作用,为鉴定AP2/ERF功能开辟了道路。而关于龙眼ERF(DlERF)家族的系统分析未见报道。
龙眼(DimocarpuslonganLour.)原产于中国南部和越南南部的亚热带区域,是重要的经济作物。龙眼具有丰富的药理作用,具有抗氧化作用、降血糖作用以及神经系统调节作用等[14]功能。研究表明,龙眼胚胎发育状态与其果实的产量和品质有着密切关系,因此对龙眼胚胎发育机理开展深入研究对龙眼产业的发展意义重大[15]。赖钟雄等[16]建立的龙眼体胚发生系统被认为是木本植物优良的模式系统之一,是开展植物胚胎发育研究良好的替代材料。到目前为止,本实验室完成了龙眼全基因组测序[17],为龙眼ERF全基因组分析提供了基础。根据前人研究表明,在龙眼体胚发生过程中乙烯可能起着诱导体胚发生以及维持生长发育过程的作用[18]。李惠华等[19]鉴定了2个乙烯受体基因(Dl-ETR1和Dl-ERS1),发现Dl-ETR1在胚性愈伤组织阶段的表达量最高,心形胚阶段的表达量最低,而Dl-ERS1 基因在子叶形胚的表达量最高;陈秋金等[20]分离了龙眼ERF1基因cDNA全长序列,并发现DlERF1的表达量随果实的发育逐渐上升,且施加外源乙烯能够抑制处于果肉快速生长期ERF1的表达。此外,研究发现拟南芥lncRNA(DRIR)能够通过调节参与应激反应的一系列基因的表达来调节植物对非生物胁迫的反应[21];miRNVL5与ERF4共同参与拟南芥对盐胁迫的反应调节[22];以及在玉米中AP2/ERF转录因子被预测为miRNA的靶基因[23]。通过以上研究,推测lncRNA、miRNA能够与ERF基因共同参与植物体的调节作用。
因此,本研究对龙眼DlERF家族成员进行系统分析。首先,对龙眼基因组所有ERF家族成员进行系统命名,分析其基本理化性质、保守基序与系统进化树等基本结构与性质,以及所有ERF家族在龙眼体胚发生早期的RNA测序中的表达量。其次,通过龙眼体胚发生早期胚性愈伤组织、不完全胚性紧实结构与球形胚3个阶段的富集分析,筛选出5个表达差异显著的ERF基因,验证其在3个阶段的表达谱以及在外源乙烯处理的EC中表达模式。最后,结合本实验室构建的龙眼体胚发生早期过程3个阶段(EC、ICpEC与GE)的lncRNA库以及龙眼miRNA文库[24],预测DlERF家族基因与lncRNA、miRNA之间的调控网络,并利用qRT-PCR技术验证其在体胚发生早期过程中的表达模式。以期为后续DlERF不同成员在龙眼生长发育过程中的功能鉴定提供信息。
1 材料和方法
1.1 材 料
试验材料为福建农林大学园艺植物生物工程研究所提供的龙眼胚性愈伤组织[25]。参照赖钟雄等[16]培养方法,获得龙眼体胚发生早期胚性愈伤组织(embryonic callus, EC)、不完全胚性紧实结构(incomplete embryonic compact structure, ICpEC)、球形胚(globular embryo, GE)。参照王亚婷等[26]的处理方法,在MS液体培养基中加入不同浓度的乙烯,处理浓度分别为0、25、50、75、100和125 μg·L-1,再接入0.15 g生长状态良好的龙眼EC,3次生物学重复。培养条件为25 ℃、115 r/min摇床上培养24 h,过滤冻存于-80 ℃冰箱,用于RNA提取。
1.2 方 法
1.2.1龙眼ERF家族成员鉴定与基本理化性质分析从GIGADB(http://gigadb.org)中获取龙眼基因组氨基酸序列以及全长核苷酸序列,并通过Pfam软件分析其结构域,鉴定出115个DlERF家族候选成员,删除冗余序列,最终确定108个DlERF家族成员。根据拟南芥ERF家族成员在龙眼基因组中的查找注释,参考拟南芥ERF命名方法,对龙眼ERF家族成员进行命名鉴定。利用Expasy(https://web.expasy.org/compute_pi/)分析DlERF家族成员的基本理化性质。
1.2.2进化树分析利用MEGA5.2软件构建系统进化树,该软件基于龙眼中ERF基因编码的氨基酸序列比对,通过邻接法和泊松校正等,进行Bootstrap分析,设置参数为1 000次重复检验。使用Interproscan(http://www.ebi.ac.uk/interpro/)识别ERF的AP2结构域的精确位置,DNAMAN对龙眼ERF基因保守结构域核苷酸序列进行比对,区分ERF家族中的亚家族。
1.2.3实时荧光定量PCR采用Tripure试剂盒提取总RNA,参照SMARTTMRACE cDNA Amplification KitTransScript miRNA First-Strand cDNA Synthesis SuperMix使用说明书进行cDNA合成。以cDNA 10倍稀释液为模板进行扩增,于罗氏LightCycler 480仪器中进行qRT-PCR检测。
采用SYBR premix Ex TaqTMⅡ kit (TaKaRa)进行qRT-PCR,反应体系为20 μL,应用2×SYBR 10 μL,cDNA模版1 μL,上下引物各0.8 μL,ddH2O 7.4 μL。程序为:95 ℃预变性30 s,95 ℃变性10 s,60 ℃退火30 s,72 ℃延伸15 s,40次循环。miRNA qRT-PCR采用TransStart Tip Green qPCR SuperMix,反应体系为20 μL,应用Tip 10 μL,cDNA模版1 μL,特异与通用引物各0.8 μL,ddH2O 7.4 μL,程序为:95 ℃预变性30 s,95 ℃变性10 s,60 ℃退火30 s,72 ℃延伸10 s,40次循环。lncRNA与mRNA以ELF-1α为内参基因,miRNA以miR172a为体胚发生早期表达内参基因,U6为乙烯处理表达内参基因。利用2-ΔΔCT方法来计算基因的相对表达量。每个取样点设3个技术重复,试验共设3次生物学重复。
1.2.4龙眼体胚发生早期lncRNAs、miRNA与DlERF关系预测以及特异性表达为了预测DlERF基因与lncRNA、miRNA之间的关系,将龙眼的ERF基因序列和miRNAs文库提交到psRNATarget (http://plantgrn.noble.org/psRNATarget/),期望值≤5,预测出DlERF基因可能作为miRNA靶基因。再通过lncRNA邻近的mRNA以及计算结合能的方法,筛选作为lncRNA靶基因的DlERF家族成员。最后将预测结果通过Cytoscape软件绘制调控网络图。
为进一步了解龙眼ERF家族各成员在龙眼体胚发生早期可能发挥的功能特点,结合龙眼转录组数据库DlERF家族基因在不同体胚发生早期阶段特异表达的FPKM值,分析DlERF家族各成员的表达情况,并对表达差异显著的5个DlERF基因的FPKM值与qRT-PCR表达量进行对比分析,同时采用 SPSS 24软件进行不同表达水平之间的差异显著性分析。利用在线网站Omicshare(http://www.omicshare.com/tools/Home/Soft/ heatmap)绘制热图、Graphpad绘制折线图。
2 结果与分析
2.1 龙眼ERF家族成员鉴定与基本理化性质分析
为了鉴定龙眼中ERF家族基因,通过Pfam软件分析其结构域,筛选出只有1个AP2/ERF结构域的DlERF家族。在‘红核子’龙眼基因组[17]中分别获得了115个ERF的全长核苷酸序列以及氨基酸序列,除去CDS相同的冗余序列,最终确定108个ERF基因的全长核苷酸序列以及氨基酸序列。经TAIR在线网站Blast比对,与拟南芥ERF家族成员进化分析,对其进行命名(表1)。
为进一步揭示龙眼ERF家族的功能以及结构特点,对DlERF家族氨基酸序列进行基本理化性质分析。结果(表1)表明,该家族的蛋白分子量为13.15 kD~63.49 kD,等电点4.53~10.82之间,不稳定系数为31.44~96.1,亲水性为-1.120~-0.284之间。龙眼ERF家族大部分氨基酸个数在300 aa以下,其中Dlo_000585.2AIL6氨基酸个数最多(575 aa)。96条DlERF基因的氨基酸序列比对到拟南芥的ERF家族成员,其中包括AtDREB26、AtTINY与AtWIND1等。值得注意的是,DlERF家族中9个成员注释到AtERF38,8个成员注释到AtERF22,5个成员注释到AtERF13等。研究表明,AtERF38与胁迫相关[27],并且被认为是次生代谢壁的候选调节剂[28];AtERF13位于SA、JA、ET和ABA信号通路的交界处,并通过协调这些激素增强植物防御反应[29]。以上均可说明,DlERF成员对龙眼的抗胁迫能力以及对病菌的防御能力可能起着重要作用。


续表1 Continued Table 1
续表1 Continued Table 1
2.2 DlERF家族的系统进化树分析
对基因组鉴定出的108个DlERF基因,构建系统进化树以研究DlERF基因系统亲缘关系。如图1所示,根据Nakano等[30]对拟南芥的分组方式,将龙眼ERF家族分为2个亚家族:CBF/DREB(A1~A6)和ERF(B1~B6)。CBF/DREB和ERF亚家族主要区别在于AP2/ERF结构域中2个位置上氨基酸残基,CBF/DREB亚家族第14位和第19位氨基酸分别是缬氨酸和谷氨酸,而ERF亚家族是丙氨酸和天冬氨酸[31]。其中CBF/DREB亚家族包括55个成员,ERF亚家族包括53个成员。虽然对应于12组的节点的自展值都不高,但是这种分支聚类可靠性是通过内含子的位置以及除AP2/ERF结构域外的保守基序支撑的。
对龙眼转录组数据库中的103个DlERF基因进行外显子分析,结果表明,103个DlERF基因中,81个基因仅包含1个外显子,13个DlERF基因包含2个外显子,包含3个外显子的有3个DlERF基因,其余包含4~9个外显子的DlERF基因均只有1个。由此可见,大部分(78.6%)龙眼ERF家族成员不含有内含子,仅有22个DlERF基因具有内含子,且这22个DlERF基因集中在其中4组中(A5、B2、B4与B6)。在A5组存在内含子的成员中,除Dlo_013069.1ERF48-1外其余的成员内含子数量均≥2,B6组存在内含子的成员中,则除Dlo_012177.1ERF15-3外其余的成员均只包含1个内含子。其余的8个组中DlERF基因都不含有内含子,这也验证了DlERF家族的保守性。

A1~A6属于CBF/DREB亚家族;B1~B6属于ERF亚家族。图1 龙眼ERF家族成员氨基酸序列系统进化分析A1-A6 belong to the CBF/DREB subfamily; B1-B6 belong to the ERF subfamilyFig.1 Phylogenetic analysis of amino acid sequences of longan ERF family members
2.3 DlERF家族基因RNA-Seq表达量分析
为进一步了解龙眼ERF家族在龙眼体胚发生早期过程中的表达情况,本研究结合龙眼体胚发生早期过程中3个阶段(EC、ICpEC与GE)的转录组数据库,对转录组数据库中所有注释为DlERF基因的FPKM值进行分析,并制作热图(图2)。除了在龙眼体胚早期3个阶段不表达的13个ERF成员外,其余的95个DlERF成员均检测到表达。
RNA-Seq分析结果显示,DlERF家族成员在龙眼体胚发生早期过程3个阶段表现出不同的表达模式。表达模式总体分为三大类:第一类,31个DlERF基因在EC阶段表达量最高,其中Dlo_013069.1ERF48-1、Dlo_025861.1TINY2-2、Dlo_017325.1ERF87、Dlo_004486.1ERF53-4、Dlo_015407.1WIN1、Dlo_022579.1ERF38-7、Dlo_014678.1ERF4-3、Dlo_027111.1ERF96-1、Dlo_030328.1ERF22-3、Dlo_002336.1ERF15-1与Dlo_009842.1ERF110-1在ICpEC至GE阶段表达量递减,Dlo_001352.1ERF95、Dlo_022634.1ERF22-1、Dlo_016788.1ERF22-6、Dlo_022353.1ERF9-1与Dlo_030665.1ERF22-8在ICpEC至GE阶段表达量递增,而其余的成员在ICpEC到GE阶段表达量变化不大,说明其余的15个DlERF基因在EC阶段高表达,可能对胚性愈伤组织的维持起作用。第二类,11个DlERF在ICpEC阶段表达量最高,除Dlo_019371.1ERF2-1、Dlo_018664.1WIND1-2与Dlo_004403.1ERF2-3外,其余的8个在EC与GE阶段的表达量无明显变化,说明了在ICpEC阶段的调控作用最为明显。第三类,53个DlERF基因在GE阶段表达量最高,Dlo_026157.1ERF73-2、Dlo_014299.1ERF13-3、Dlo_022613.1ERF5-1、Dlo_023524.1ERF71-2、Dlo_005616.1CBF3、Dlo_023666.1ERF8、Dlo_022612.1ERF1-2与Dlo_007317.1ERF38-2在EC至ICpEC阶段表达量递减,Dlo_013163.1DREB2、Dlo_010734.1ERF3-2、Dlo_007513.1ERF70-3、Dlo_025226.1ERF70-4、Dlo_017388.1ERF10-2、Dlo_000290.1ERF70-1与Dlo_000247.1ERF4-1在EC至ICpEC阶段表达量递增,其余的在EC与ICpEC阶段表达量变化不大。以上结果表明,不同的DlERF基因可能参与龙眼体胚发生早期不同阶段的维持。

EC. 胚性愈伤组织; ICpEC. 不完全胚性紧实结构; GE.球形胚图2 龙眼ERF家族在龙眼体胚早期3个阶段的特异表达分析EC. Embryogenic callus; ICpEC. Incomplete embryonic compact structure; GE. Globular embryoFig.2 Specific expression analysis of longan ERF family in three stages of early longan somatic embryo
总体而言,龙眼ERF家族成员在龙眼体胚发生早期的形态建成中发挥重要作用,大部分成员在GE阶段高表达,特别是在个别阶段表现出阶段特异性的DlERF基因,可能在该阶段对形态功能等的维持起到了一定作用。
2.4 龙眼体胚发生早期DlERF基因表达的qRT-PCR分析及乙烯处理下的表达
为了更好地了解ERF家族在龙眼体胚发生早期的可能调控作用,通过在龙眼体胚发生早期3个阶段(EC、ICpEC与GE)RNA-Seq表达两两对比,筛选出在龙眼体胚发生早期表达差异显著的5个ERF基因(Dlo_008317.1ERF15-2、Dlo_009070.1ERF106-3、Dlo_022634.1ERF22-1、Dlo_009939.1ERF98与Dlo_022310.1ERF1-1)。qRT-PCR分析5个DlERF的表达趋势(图3,A)与RNA-Seq的表达结果(图3,B)进行对比。由图3可以看出,除Dlo_022634.1ERF22-1外,其余4个DlERF从EC到GE阶段,RNA-Seq与qRT-PCR中的整体表达趋势一致。Dlo_008317.1ERF15-2、Dlo_022310.1ERF1-1与Dlo_009939.1ERF98在GE阶段qRT-PCR中的表达量显著高于EC与ICpEC阶段;Dlo_009070.1ERF106-3则在EC阶段qRT-PCR中的表达量最高,显著高于GE与ICpEC阶段;Dlo_022634.1ERF22-1在ICpEC阶段表达量显著高于EC与GE阶段。从不同表达趋势来看,DlERF在龙眼体胚发生早期过程不同阶段都发挥了作用。

A. 实时荧光定量PCR;B. RNA测序;EC. 胚性愈伤组织; ICpEC. 不完全胚性紧实结构; GE.球形胚;不同小写字母表示阶段间差异显著(P<0.05)图3 DlERF在龙眼体胚发生早期3个阶段的表达A. qRT-PCR; B. RNA Seq; EC. Embryogenic callus; ICpEC. Incomplete embryonic compact structure; GE. Globular embryo; The normal letters indicate significant differences among different stages (P <0.05)Fig.3 Expression of DlERF in the early three stages of longan somatic embryogenesis
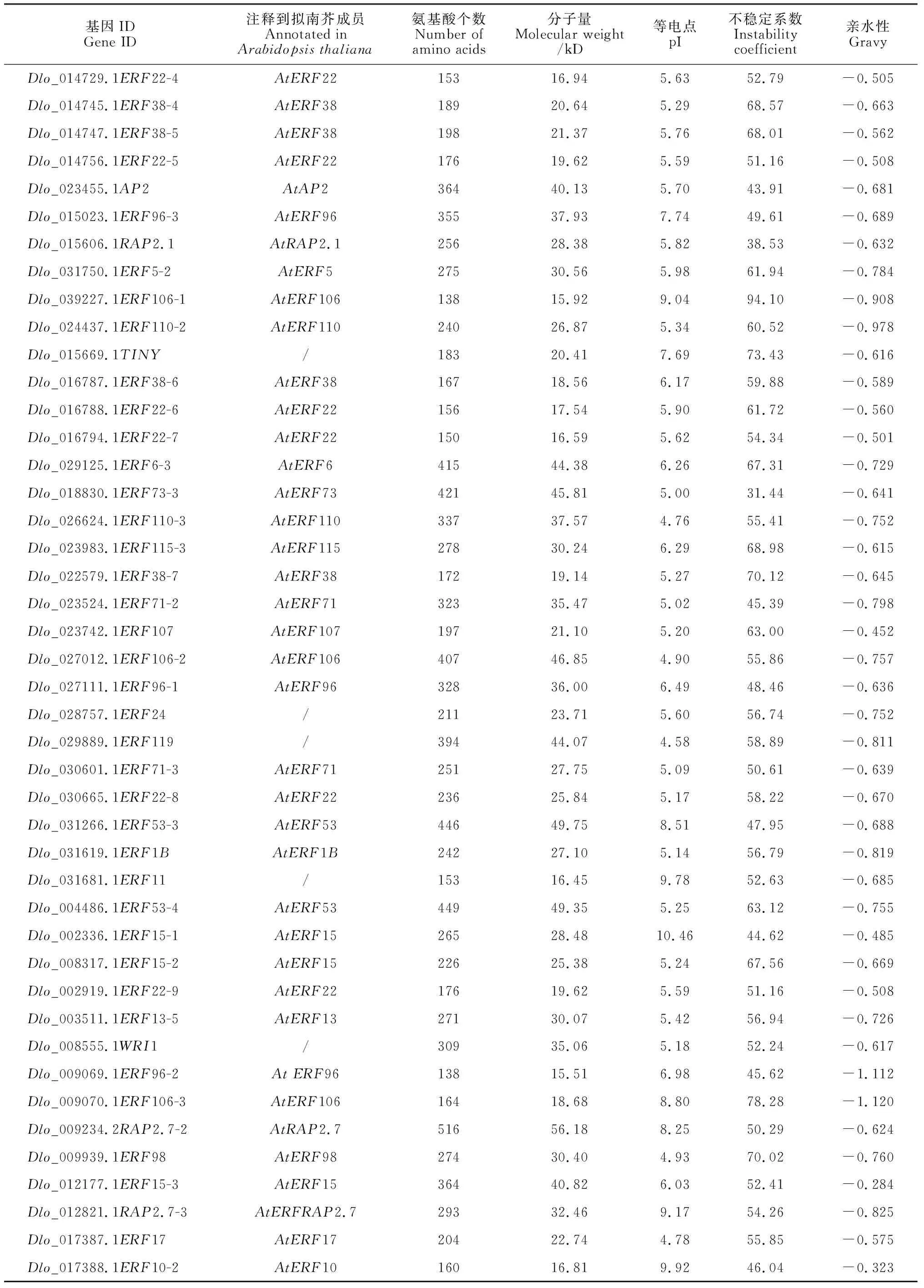
使用qRT-PCR研究了DlERF在不同浓度乙烯处理下的表达(图4)。在不同乙烯浓度24 h处理下,5个DlERF基因均表现出差异的表达模式。与对照(CK)相比,不同梯度浓度的乙烯处理使得DlERF基因显著下调,虽然Dlo_022310.1ERF1-1、Dlo_009939.1ERF98与Dlo_022634.1ERF22-1在乙烯处理浓度50 μg·L-1以及Dlo_008317.1ERF15-2乙烯处理浓度为125 μg·L-1时表达量显著上升,但总体仍为负调控趋势。
2.5 DlERF家族成员与lncRNA、miRNA之间的关系预测与表达模式分析
通过对lncRNA邻近的mRNA以及计算结合能的方法对lncRNA靶基因进行预测,本次对108个DlERF进行lncRNA的靶基因预测。5个不同的DlERF基因分别被预测为LTCONS_00052840、LTCONS_00055212、LTCONS_00016255、LTCONS_00013739与LTCONS_00043464的靶基因。5个DlERF基因都属于lncRNA的顺式调控靶基因,其中除了LTCONS_00043464与对应DlERF基因位置关系为lncRNA位于mRNA上游10 k内外,其余的4个均为lncRNA位于mRNA下游20k内。
不同小写字母表示处理间差异显著(P<0.05)图4 不同乙烯浓度处理下DlERF的表达模式The normal letters indicate significant differences among treatments (P <0.05)Fig.4 Relative expression pattern of DlERF in different ethylene concentrations
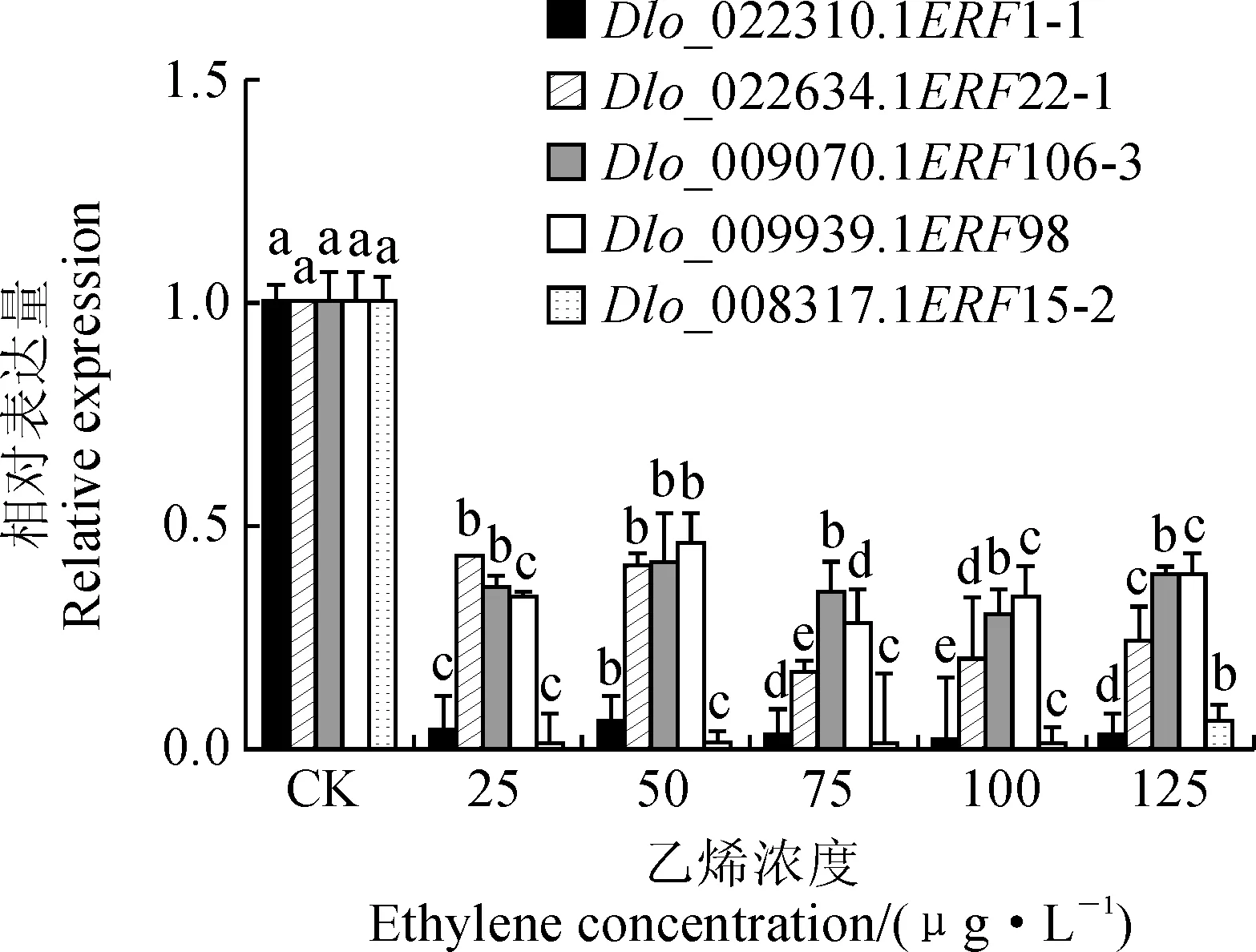
利用psRNATarget 在线预测(期望值≤5),预测龙眼 miRNA 数据库中靶向调控DlERF家族成员的miRNA。结合DlERF与相关lncRNA、miRNA的关系预测,构建了龙眼体胚发生早期差异显著的ERF-lncRNA-miRNA的调控网络(图5,A)。分析结果表明,Dlo_009070.1ERF106-3、Dlo_009939.1ERF98、Dlo_008317.1ERF15-2与Dlo_022310.1ERF1-1均同时被2个miRNA调控,且Dlo_008317.1ERF15-2能作为LTCONS_00013739的靶基因。值得注意的是,Dlo_miR413能够同时调控Dlo_009070.1ERF106-3与Dlo_008317.1ERF15-2。
使用qRT-PCR分析了5个DlERF以及相关lncRNA、miRNA在EC、ICpEC与GE中的表达模式(图5,B)。结果表明, LTCONS_00013739对靶基因Dlo_008317.1ERF15-2为正调控关系,从EC到GE阶段表达量显著升高;在EC至ICpEC阶段以及ICpEC至GE阶段,Dlo_miR164a通过靶向Dlo_022634.1ERF22-1负调控其表达;Dlo_miR413与Dlo_miR1510a共同靶向Dlo_009070.1ERF106-3,从EC到GE阶段均表现出显著负调控关系,而Dlo_miR413与Dlo_008317.1ERF15-2的表达量表现出正相关,推测相比于Dlo_008317.1ERF15-2,Dlo_miR413更倾向于调控Dlo_009070.1ERF106-3;调控Dlo_009939.1ERF98相关的Dlo_miR408与Dlo_miR774b,从表达趋势来看,Dlo_miR774b在龙眼体胚发生早期过程能负调控Dlo_009939.1ERF98;Dlo_miR399c与Dlo_022310.1ERF1-1在EC到GE阶段可能存在负调控关系。
使用qRT-PCR研究了DlERF及其相关的miRNA、lncRNA在不同浓度乙烯处理下的表达(图6)。结果表明,Dlo_miR413在不同浓度乙烯处理均没有差异。乙烯处理对LTCONS_00013739与其靶基因Dlo_008317.1ERF15-2表现出显著抑制作用;Dlo_022634.1ERF22-1 及Dlo_miR164a在从75、100至125 μg·L-1乙烯处理下表现为负调控关系;与对照(CK)相比,Dlo_miR1510a在75 μg·L-1乙烯处理下表达量显著上升,与其靶基因Dlo_009070.1ERF106-3则表现相反趋势;除CK外,Dlo_009939.1ERF98在50 μg·L-1乙烯处理下表达量最高,与其相关的Dlo_miR774b则在50 μg·L-1乙烯处理下表达量最低,表现出显著的抑制作用;与CK相比,Dlo_022310.1ERF1-1在125 μg·L-1乙烯处理下表达量显著下降,与其相关的Dlo_miR399c则显著上升。根据前面对DlERF及其相关的miRNA、lncRNA在体胚发生早期的表达趋势分析对比可知,大部分的DlERF及其相关的miRNA、lncRNA在不同浓度乙烯处理下仍然表现出相同的调控趋势。
龙眼体胚发生早期过程中,ERF基因及可能对其起到调控作用的miRNA、lncRNA的不同作用模式,以及在不同阶段、不用浓度乙烯处理中的差异表达,表明了DlERF在龙眼体胚发生早期过程中可能存在复杂的调控机制。
EC. 胚性愈伤组织; ICpEC. 不完全胚性紧实结构; GE.球形胚图5 龙眼体胚发生早期差异表达的ERF基因与其相关的miRNA、lncRNA的表达趋势EC. Embryogenic callus; ICpEC. Incomplete embryonic compact structure; GE. Globular embryoFig.5 Trends of expression of ERF genes and their associated miRNAs and lncRNAs in the early stage of longan somatic embryogenesis

图6 不同乙烯浓度处理下DlERF基因与其相关的miRNA、lncRNA的表达趋势Fig.6 Trends of expression of DlERF genes and their associated miRNAs and lncRNAs in different ethylene concentrations
3 讨 论
3.1 龙眼体胚发生早期过程中大量的ERF基因在GE阶段高表达
AP2/ERF是一个庞大的转录因子家族,能够对植物生长发育[32-33]、基因表达[34]以及生物[35]与非生物胁迫[36]等方面发挥重要作用,并且在植物体胚发生过程中也起着至关重要的调控作用[13]。然而在龙眼ERF全基因组家族还未进行系统鉴定,大多数DlERF基因在龙眼中的功能和作用仍然未知。
Nakano等[30]对拟南芥与水稻ERF家族进行全基因组分析,包括基因结构,系统发育,染色体位置和保守基序等。因此本研究对龙眼ERF家族进行系统地分析,从龙眼数据库中鉴定了108个ERF家族基因,通过系统进化树的构建以及保守基序分析表明,108个ERF家族基因具有相同的保守基序,AP2/ERF结构域在龙眼中相对最为保守,系统进化树节点的自展值相对较低,基于DlERF保守基序分析,推测除AP2/ERF结构域之外,其余部分成员间差距较大。对5个在龙眼体胚发生过程中富集的ERF基因进一步分析发现,由于AP2/ERF结构域中2个位置上氨基酸残基的差距,5个龙眼体胚发生早期差异表达DlERF基因属于ERF家族中的2个亚家族。研究表明在拟南芥ERF家族成员中,AtERF1能够在非生物胁迫下扮演正调控的角色[37];AtERF15不仅是ABA响应的正调节因子,并且在根与胚胎中高度表达[38];以及AtERF98能够调节植物重要抗氧化剂生物合成,有助于提高拟南芥的耐盐性[39]。可见,在龙眼体胚发生早期过程中差异表达的ERF基因可能对提高龙眼抗逆性起到十分重要的作用,并且Dlo_022310.1ERF1-1、Dlo_008317.1ERF15-2与Dlo_009939.1ERF98在龙眼体胚发生早期过程中表达量均呈现上升趋势。这也进一步说明, 随着龙眼体胚发生早期进行,內源激素[40]等方面发生显著变化,为了促进胚胎的正常发育,需要更多的DlERF基因参与。
在本次研究中,为了鉴定龙眼体胚发生过程中ERF基因的表达模式,对101个在龙眼体胚发生早期过程检测到表达的ERF基因进行RNA-Seq表达分析。大部分DlERF基因在GE阶段表达量最高,其次是EC阶段。Boutilier等[41]与Tsuwamoto等[42]鉴定一些AP2/ERF基因能够促进胚胎发育,并推测他们在SE的诱导过程中具有特定的功能。通过对5个在龙眼体胚发生早期差异显著表达的DlERF基因进行qRT-PCR与RNA-Seq表达结果对比发现,大部分在龙眼体胚发生早期3个阶段的表达趋势一致,且其中3个DlERF基因在龙眼GE阶段表达量最高。与蒺藜苜蓿体胚发生过程中发现的MtSERF1相同趋势,在球形胚分生组织中高表达[12]。由此可见,从EC到GE阶段需要更多的ERF基因来参与GE阶段的维持,也进一步说明在体胚发生早期过程中,细胞分裂分化、抗性等方面也逐渐增强。
3.2 DlERF基因与乙烯之间存在负调控关系并且可能与miRNA、lncRNA形成调控网络
乙烯在体胚发生过程中起着重要的作用[43-44]。前人研究发现,在龙眼体胚发生发育的过程中,乙烯含量在不完全胚性紧实结构与心形胚分别出现了峰值,松散型的胚性愈伤组织、球形胚以及子叶形胚的表达量相对较低[18]。同样也有研究表明,对于依赖 GCC-box 表达的ERF基因来说,既可以是转录激活因子,也可以是转录抑制子,并且只有一小部分受乙烯调节[45]。因此对龙眼EC使用不同浓度乙烯处理时发现,与CK相比,在不同浓度处理下5个DlERF基因均表现为显著下调。结果表明,乙烯与龙眼体胚发生早期5个差异表达的ERF基因存在负调控关系。另一方面,5个DlERF基因在外源乙烯处理下的表达趋势是相似的,这意味着这5个DlERF基因可能在应对外源不同浓度乙烯处理时产生类似的响应,也进一步表明其功能保守性。
除了与基因相互作用外,ERF基因也可能与miRNA、lncRNA相互作用。目前,lncRNA在植物体胚发生过程调控作用的研究还未见报道,但是关于植物中lncRNAs的研究已有部分研究,多数以模式植物[46-47]与水稻[48]等作物为研究对象。但对于miRNA来说,已经在龙眼中做了大量的研究,例如:Dlo_miR156家族成员与早期胚胎培养发育阶段相关[49],Dlo_miR162与多个miRNA共同调节体细胞胚胎发育的特定阶段[50]等。分析结果显示,一个DlERF可能被多个miRNAs靶向,一个miRNA也可能靶向多个DlERF基因。通过对龙眼体胚发生早期过程中以及不同浓度乙烯处理下差异表达5个ERF基因的调控网络进行qRT-PCR验证,不仅初步验证了DlERF基因与其相关的miRNA、lncRNA可能存在的调控关系,同时推测对于2个miRNA同时调控同一个DlERF基因,并且miRNA对DlERF的调控作用具有偏好性。因此认为可能由于存在多个调控网络,导致其难以从表达量中发现其调控趋势。从复杂的关系网络中可以看出,作为生物与非生物胁迫相关的基因,在龙眼体胚发生过程中发挥着至关重要的作用,以上研究仅仅是DlERF基因与lncRNAs、miRNA调控网络中的冰山一角,在龙眼体胚发生早期过程中,DlERF所发挥的功能以及调控作用需要在今后的工作中进行研究验证。
