2014-2017年山东省部分规模化猪场PRRSV ORF5基因的遗传演化分析
2018-12-29曲向阳孙英军张洪亮刘媛媛任夫波赖清华周明明杨汉春
曲向阳,孙英军,张洪亮,周 磊,刘媛媛,任夫波,赖清华,夏 天,周明明,杨汉春
(1.中国农业大学动物医学院,北京 海淀 100193;2.环山集团股份有限公司养猪事业部,山东 青岛 266000;3.中国农业科学院哈尔滨兽医研究所,黑龙江 哈尔滨 150069)
猪繁殖与呼吸综合征(PRRS)是由猪繁殖与呼吸综合征病毒(PRRSV)引起的以母猪繁殖障碍、保育育肥猪呼吸道症状与高死亡率为主要特征的病毒性传染病。1987年美国最早报道此病[1],1996年郭宝清等从流产胎儿中分离到PRRSV,证实我国存在PRRSV感染[2]。2006年,我国暴发以高热、高发病率与高死亡率为特征的“高热病”,童光志等从全国12个省市的猪场分离到以NSP2缺失90个核苷酸为特征的PRRSV变异株[3]。2012年以来国内陆续分离到与美国NADC30毒株同源性很高的毒株[4],统称为类NADC30毒株(NADC30-like),NADC30-like重组毒株和变异株的不断出现[5-6],给该病的防控带来了巨大的挑战。
PRRSV基因组全长约15 kb,至少包括10个开放阅读框,其中 ORF1a、ORF1b编码非结构蛋白,ORF2a-ORF7编码结构蛋白。ORF5基因编码的GP5蛋白是PRRSV中变异性最大的结构蛋白,常作为流行病学调查与遗传演化分析的研究对象。本研究对2014-2017年山东部分地区96个规模化猪场的236份PRRS疑似样品进行检测,并对阳性样品进行ORF5基因测序及遗传进化分析,以期阐述山东省规模化猪场PRRS的流行及病毒变异趋势,为PRRS防控提供参考依据。
1 材料与方法
1.1 病料采集与处理 2014-2017年间从山东部分规模化猪场(主要分布区域:烟台、威海、青岛、潍坊、临沂、淄博、日照、滨州等)采集236份疑似PRRSV感染病猪的肺脏、淋巴结、脾脏等病料,将组织研磨均匀,取一部分提取RNA,其余样品在-80℃保存。
1.2 主要试剂 RNeasy plus Mini Kit,购自Qiagen公司;AMV反转录酶、dNTPs、LA Taq DNA聚合酶和pMD18-T,载体等,购自TaKaRa公司;Gel Extraction Kit,购自OMEGA公司;TG1感受态由本实验室制备。
1.3 引物设计 根据GenBank中的登录的PRRSV NADC30(JN654459)的基因序列,针对ORF5序列利用Oligo6.0设计一对引物用于PRRSV的检测和测序。引物序列由上海生工生物工程技术服务有限公司合成。
1.4 PRRSV的ORF5基因扩增与测序 取300 μL研磨后的组织上清液,提取总RNA,经下游引物PRRSV-ORF5-R反转录获得cDNA。PCR反应程序为:94℃ 2 min;94℃ 30 s、55℃ 30 s、72℃ 1 min,共35个循环,72℃ 10 min。对PCR产物用1%琼脂糖凝胶电泳,用Gel Extraction Kit对目的片段进行回收,纯化后的PCR产物克隆到pMD18-T载体中。重组克隆由上海生工生物工程技术服务有限公司进行测序。
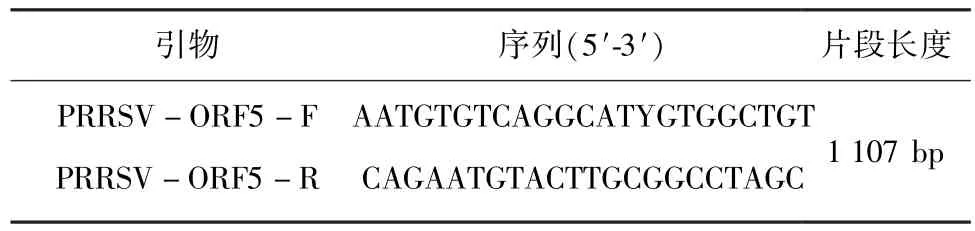
表1 ORF5引物序列
1.5 PRRSV遗传演化分析 利用MEGA5.1软件的Cluster W对GenBank中11株代表性PRRSV和检测到的122株PRRSV的ORF5核苷酸序列比对,并利用Neighbor-joining Method对其进行了遗传演化分析,bootstrap数为100。
1.6 PRRSV ORF5核苷酸序列及推导氨基酸序列分析 利用DNAStar 7.1软件包中的MegAlign分析ORF5基因核苷酸同源性及推导氨基酸序列的同源性。分析代表性毒株GP5推导氨基酸的遗传变异。
2 结果
2.1 PRRSV RT-PCR检测结果 对2014-2017年猪场疑似PRRSV的236份样品进行检测,共122份PRRSV阳性,RT-PCR结果见图1,总体阳性率51.7%。2014年共检测123份,58份阳性,阳性率47.2%;2015年共检测14份,3份阳性,阳性率21.4%;2016年共检测54份,23份阳性。阳性率42.6%;2017年共检测45份,38份阳性,阳性率84.4%。各年度PRRSV阳性率差异与样品的选择有关。
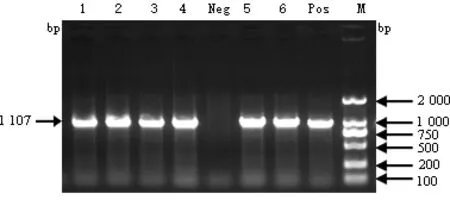
图1 部分病料样品ORF5片段的RT-PCR检测结果
2.2 ORF5基因的遗传演化分析 利用MEGA5.1软件对GenBank中的11株PRRSV代表性毒株和所检测的122株PRRSV进行遗传演化分析,结果显示,本研究所有PRRSV毒株均为美洲型,全部属于Lineage 1、3、5 和8 (图2)。 本研究中,以VR-2332为代表的 Lineage 5毒株 44株(占 36.1%),以HuN4为代表的Lineage 8毒株19株(占15.6%),以QYYZ为代表的Lineage 3毒株2株(占1.6%),以NADC30为代表的Lineage 1毒株57株(46.7%)。表明山东省 PRRSV存在4个 Lineage的毒株,其中以 Lineage 1、Lineage 5和 Lineage8的毒株为主要流行毒株(表2)。此外,2014年检测到的Lineage 1的一个分支与美国Minnesota 14同源性最高,且在进化树中处于同一分支,预示这类毒株可能由美国引入,这16份样品均来自于同一个养猪体系,在小区域内流行,但并没有在我国广泛流行,本研究是首次报道我国存在这类PRRSV毒株。
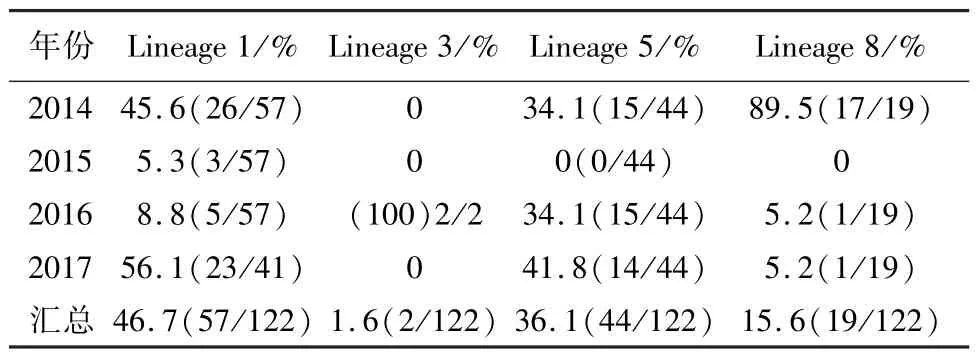
表2 2014-2017年山东地区不同PRRSV毒株的检出率
2.3 ORF5基因核苷酸序列及其推导氨基酸序列分析 将所测序的ORF5基因序列与VR-2332、HuN4、QYYZ、Minnesota 14 及 NADC30 进行序列比对,所检测PRRSV ORF5核苷酸序列同源性与氨基酸同源性分别是:78.9% ~99.7%,73.7% ~99.5%(表3)。Lineage 1中的分离株与NADC30和Minnesota 14的核苷酸和氨基酸同源性最高,分别为84%~96.2%,84.3% ~94.3%和85.2% ~97.4%,78.9% ~96.9%。Lineage 3中的分离株与QYYZ的核苷酸和氨基酸同源性最高,分别为97.2% ~97.7%,98.0%~98.5%。Lineage 5中的分离株与VR-2332的核苷酸和氨基酸同源性最高,分别为97.6%~99.0%,95.9% ~99.4%。Lineage 8中的分离株与HuN4的核苷酸和氨基酸同源性最高,分别为96.8% ~99.7%和96.0% ~99.5%。
将部分分离株ORF5基因的推导氨基酸序列比对显示:信号肽区 Lineage 1 10Y;Lineage 3 6S,14F,24S,25I;Lineage 5 16S;Lineage 8 9C,13R,23Y是各群独有的特征(图3)。胞外域Lineage 1 47I;Lineage 3 30S,32N,33G,38Y,39S,59N;Lineage 5 29A,92A;Lineage 8 35N,39I,94A为各群独有的氨基酸(图3)。跨膜域Lineage 1 124A;Lineage 3 66C,98A,101F,102H,117F,128V;Lineage 5 66S,101F,102V,121T,127F为各群独有的氨基酸(图 3)。胞内域Lineage 3 151K,152I,170G,191K,192I,196R,199H,200P;Lineage 5 137A,164R,185V,189I,200P;Lineage 8 161V,189L,200L为各群独有的氨基酸(图3)。
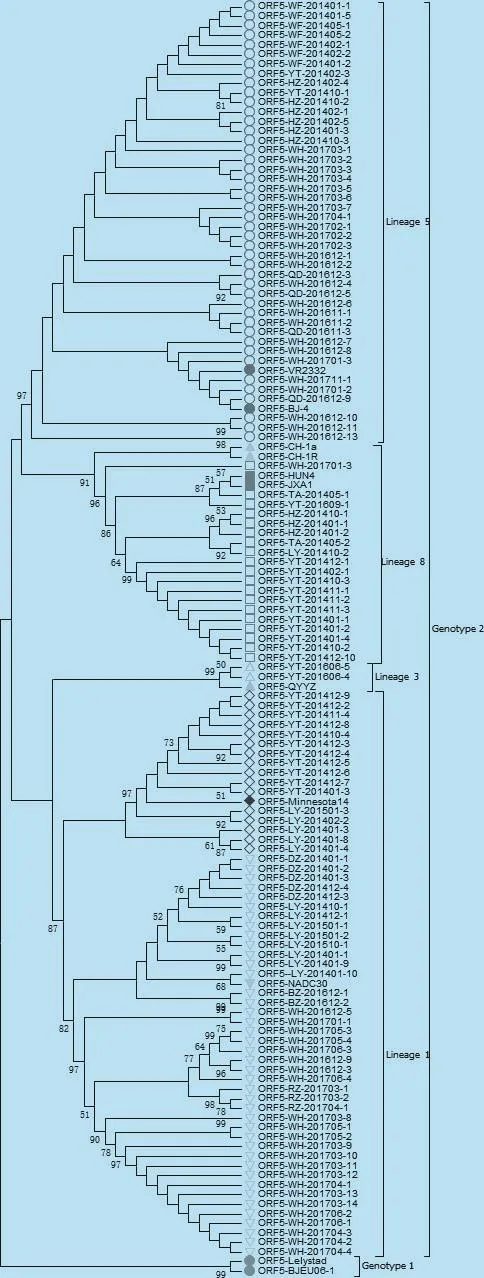
图2 山东地区PRRSV ORF5遗传进化树分析
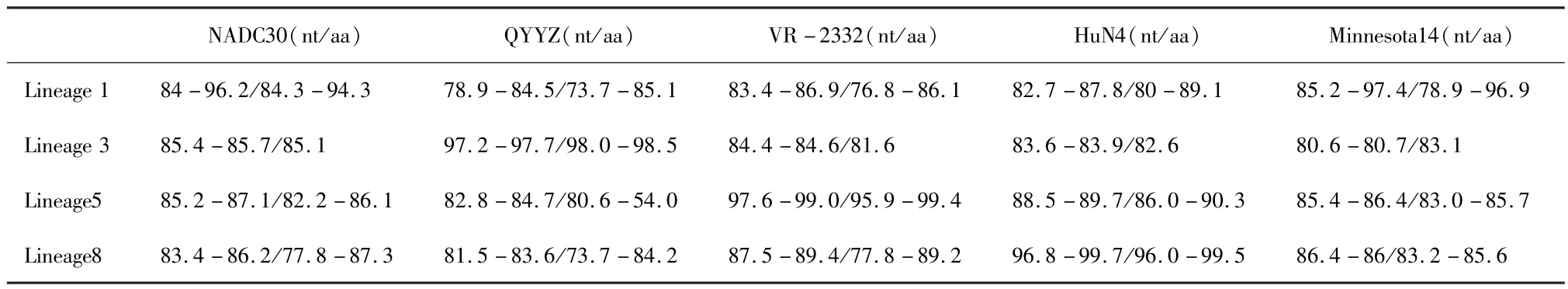
表3 与参考毒株的核苷酸及氨基酸相似性 (%)
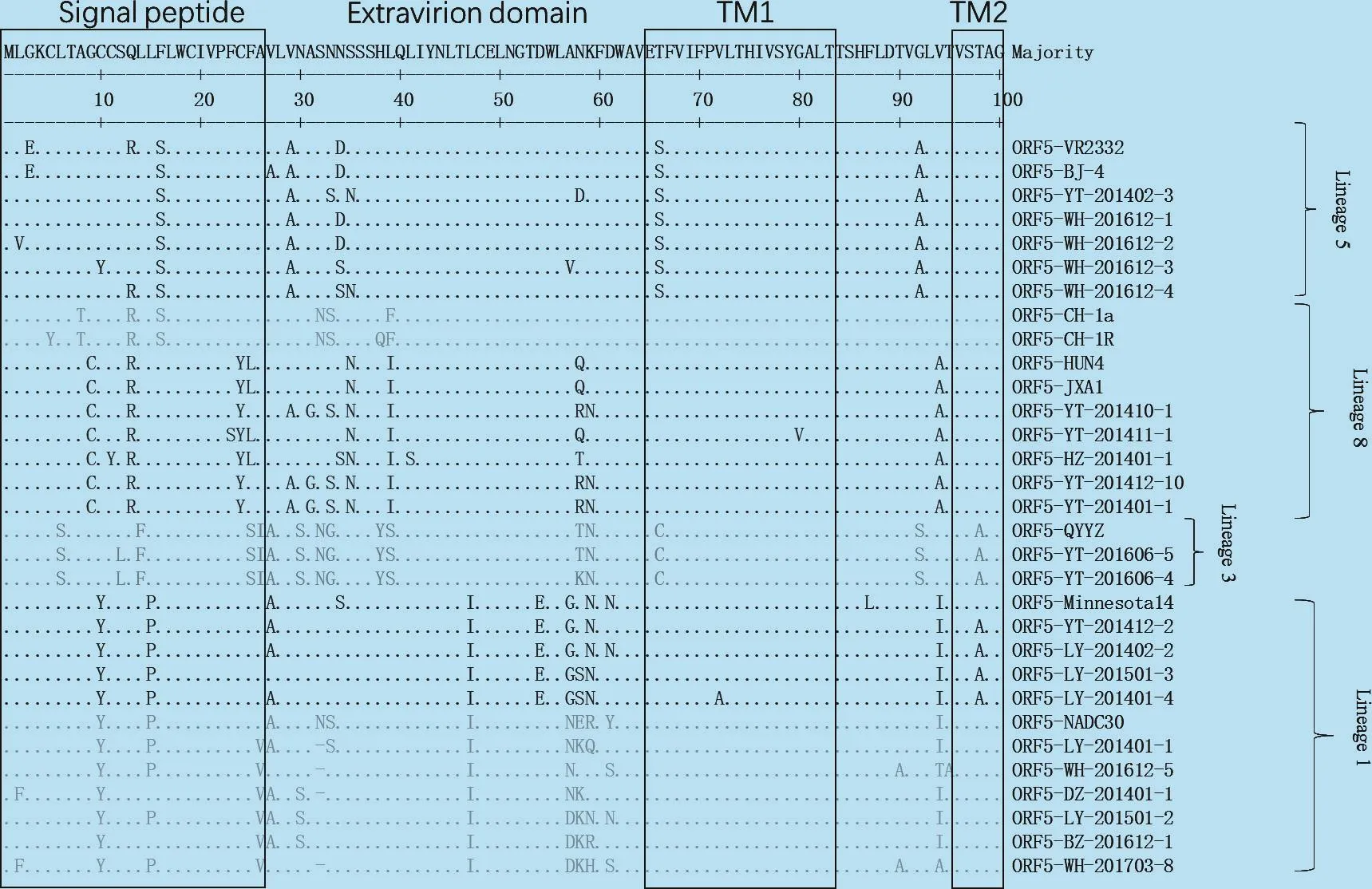
图3 ORF5基因推导的氨基酸序列比对分析
3 讨论
1996年,我国首次分离到 PRSSV,并命名为CH-1a株。2006年,我国首次出现HP-PRRSV,随后蔓延至全国各地,给我国养猪业带来了巨大的经济损失。在随后的几年里,我国先后报道了QYYZ-Like毒株,NADC30-Like毒株等各种新出现的PRRSV[7]。为研究山东省PRRSV的流行状况,在2014-2017年我们对山东省部分规模化猪场的PRRSV进行了跟踪检测,对阳性样品进行ORF5的测序与遗传演化分析,分析了山东省存在的PRRSV ORF5基因遗传变异。
PRRSV分为2个基因型,基因1型(欧洲型)和基因2型(美洲型)。我国主要流行PRRSV为美洲型。ORF5遗传演化分析表明:山东省流行的PRRSVs主要为4个Lineages,分别是以类NADC30为代表的Lineage 1,以QYYZ为代表的Lineage 3,以VR-2332为代表的Lineage 5和以HuN4为代表的Lineage 8。其中Lineage 5与Lineage 1分别占比36.1%与46.7%,为主要流行毒株。2014-2017年间,Lineage 8(HP-PRRSV)的检出率在逐年下降,而Lineage 1(类NADC30)检出率在显著升高,Lineage 5的检出率较稳定。2017年所检测到的38份阳性样品中,有23份为Lineage 1(类NADC30毒株),占比60.5%,14份为 Lineage 5(类 VR-2332毒株),占比36.8%,说明类NADC30毒株流行越来越广泛。目前商品化的疫苗对NADC30毒株的交叉保护性不佳[8],故对该毒株需要给予特别关注。类VR-2332 PRRSV高检出率可能与经典株PRRSV MLV疫苗的广泛应用有关。Lineage 3是2012年新出现的毒株,主要流行于我国南方各猪场,且广泛存在(21.3%,32/150)[9],该类毒株(GM2 和QYYZ)为弱毒疫苗与野毒重组的新型重组毒株[10]。近年来,随着南方的集团化养猪企业在北方各省建场及后备母猪引种,该类毒株开始在北方出现,但尚未成为主流毒株,其致病性与HP-PRRSV类似,但在未免疫猪场该毒株会造成较大损失。此外,本研究在2014年检测到Lineage 1的Minnesota 14分支毒株可能由美国引入,该类毒株可能不适应本地环境,尚没有在我国广泛流行,但是并不代表这类毒株会在中国消失殆尽,一旦这类毒株发生变异,有可能再次在中国出现和流行,引起新的疫情,因此有必要加大对这类毒株的监测。
ORF5基因是研究PRRSV遗传演化的重要靶向目标之一。本研究中测得的122条ORF5基因与参考毒株 VR-2332、HuN4、QYYZ、Minnesota 14 和NADC30的序列比对发现,ORF5基因遗传变异较大,尤其是Lineage 1的毒株。各亚群ORF5基因推导氨基酸比对发现,各亚群PRRSV毒株在ORF5基因的进化上具有保守性,在ORF5的信号肽区、胞外域、跨膜域和胞内域各亚群毒株具有各自独特的氨基酸特征[11]。
本研究对2014-2017年山东省部分规模化PRRSV ORF5基因进行了详细的分析,阐释了山东省近年来流行的4个Lineage的PRRSVs,并分析了各亚群PRRSV ORF5基因的分子特征,丰富了山东省PRRSV的流行病学数据,为区域化PRRS的防控与净化提供了科学依据。
