一个新的黄瓜叶色突变体的光合特性分析
2018-12-20胡亮亮赵子瑶张海强陈菲帆张朝文戎福喜李玉红
胡亮亮,赵子瑶,张海强,陈菲帆,张朝文,戎福喜,陈 鹏,李玉红
(1.西北农林科技大学 园艺学院,陕西杨凌 712100;2.西北农林科技大学 生命科学学院,陕西杨凌 712100)
叶片是植物进行光合作用的重要器官,叶色变化会影响植物的光合效率进而影响产量,因此叶色变异方面的研究受到不少科研工作者的高度重视[1-2]。叶色突变体是研究叶色变异形成机制的理想材料, 被广泛应用于植物光合作用、光形态建成、叶绿体结构功能与遗传发育调控机制等研究中[3-5]。此外,叶色突变体也可作为标记性状用于辅助育种及杂一代种子的生产中[6-9]。
叶色突变体来源较广,其产生方式主要有自然突变、插入突变和物理化学诱变等[10]。叶色突变体在水稻、玉米、小麦和大豆等作物中均有大量报道[5,11-14],如在水稻上有200多个叶色突变体,其中有50多个叶色突变的候选基因被克隆[15],玉米上已报道的叶色突变体超过210个[13]。在大豆上已定位了25个导致叶绿素缺陷的相关基因[12]。相比较以上植物,在黄瓜上有关叶色突变体的研究还相对滞后,只报道了10余个叶色突变体,如黄叶转绿(v)、黄叶转绿-1(v-1)、黄色子叶-1(yc-1)、黄色子叶-2(yc-2)、白化致死(pl)、白化转绿(albin)、光敏白化致死突变(ls)、叶绿素缺陷突变(cd)、黄转绿白斑叶(vvi)、金黄叶(yp)、金黄色子叶(gc)和黄绿叶(ygl)[16-20],仅克隆到黄瓜的金黄叶突变体C528和黄绿叶突变体9110Gt的候选基因[19-21],且对这些突变体的研究多集中在形态特征描述及遗传规律分析方面,对其光合特性的研究报道较少,与大田作物如水稻、玉米等相比,报道的黄瓜叶色突变仍然很少,远不能满足对其叶色突变机制的解析及利用。
西北农林科技大学园艺学院黄瓜课题组在前期研究中发现1个新的黄瓜黄化转绿突变体C777。经过连续3 a的种植观察,发现该突变体能够稳定遗传。本试验从叶绿素质量分数、光合速率及叶绿素荧光参数等方面对突变体C777及其野生型CCMC进行测定分析,旨在了解该突变性状对光合作用效率的影响,为进一步解析该突变体叶色变异的调控机制奠定基础,也为合理利用这一新种质提供理论依据。
1 材料与方法
1.1 试验材料
黄绿叶突变体C777由黄瓜自交系CCMC通过EMS化学诱变得到。该突变体子叶抽出时即为黄色,随着第1片真叶的长出,子叶慢慢转绿。第1片真叶长出时也为黄色,随着第2片真叶的抽出慢慢转绿,突变体整个生育期都表现出这种转色现象。野生型CCMC叶色为绿色;突变体C777和野生型CCMC均由西北农林科技大学园艺学院黄瓜课题组提供。2016-03-15将突变体和野生型种子各150粒播种于72孔穴盘中进行育苗,4月10日定植于西北农林科技大学园艺场,株距25 cm,行距60 cm,试验设野生型和突变体2个处理,在幼苗的第4真叶期进行相关指标测定。
1.2 试验方法
1.2.1 光合色素质量分数的测定 用80%丙酮提取光合色素,参照Hill等[22]的方法用分光光度法分别测定叶绿素a、叶绿素b和类胡萝卜素的质量分数。根据 C777 叶色由黄转绿的时间,取样和测定指标时分2个时期,即 C777黄化期和转绿后,分别用时期Ⅰ和时期Ⅱ来表示。突变体和野生型各重复3次,每次重复测定10株。
1.2.2 突变体C777光合相关参数的测定 利用Li-6800(美国LI-COR 公司)型便携式植物光合测定仪,在晴朗天气9:00-11:00时间段内,开始测定幼苗功能叶(从顶芽往下数第2和第3片叶)的净光合速率(Pn)、蒸腾速率(Tr)、叶片气孔导度(Gs)和胞间CO2摩尔分数(Ci)。光强设置为1 000 μmol·m-2·s-1,CO2注入系统设定值为400 μmol·mol-1,气体流量为500 μmol·s-1。叶片温度及其周围的水气压分别为(29±1)℃和(1.7±0.1) kPa。分别测定突变体Ⅰ期和 Ⅱ期的光合相关参数。重复3次,每次重复测定3株。

用Fc800 FluorCam大型版开放式多光谱荧光成像系统(捷克PSI公司)对转色前的突变体和野生型的离体叶片进行荧光成像分析。通过成像情况来直观地比较突变体和野生型叶片的最大光量子产量QYmax(即Fv/Fm)之间的差异。
1.3 数据分析
采用Execl 2007和SPSS 13.0软件进行数据的统计与显著性分析,处理间显著性分析采用Duncan’s法(α=0.05)。
2 结果与分析
2.1 突变体C777的光合色素质量分数
由表1可以看出,在Ⅰ期,C777的总叶绿素质量分数为0.50 mg·g-1,显著低于野生型。突变体叶片的各光合色素质量分数也均显著低于野生型。叶绿素a、叶绿素b、总叶绿素、类胡萝卜素分别为野生型叶片相应色素质量分数的27.27%、23.53%、26.46%和40.00%。在Ⅱ期,突变体C777叶片转绿之后,各光合色素的质量分数显著升高,C777总叶绿素质量分数比转色前叶绿素质量分数升高3.5倍,但与野生型相比,仍略低于野生型。叶绿素a、叶绿素b、总叶绿素以及类胡萝卜素为野生型相应色素质量分数的82.95%、75.00%、81.11%和83.33%,由于叶绿素质量分数变化幅度显著高于类胡萝卜素(表1),由此初步推断叶绿素质量分数的变化是引起突变体叶色变化的主导色素,该黄瓜黄绿叶突变体为总叶绿素缺陷型突变体。突变体C777在Ⅰ期和Ⅱ期叶绿素a/b均大于野生型,分别为5.00和4.87。
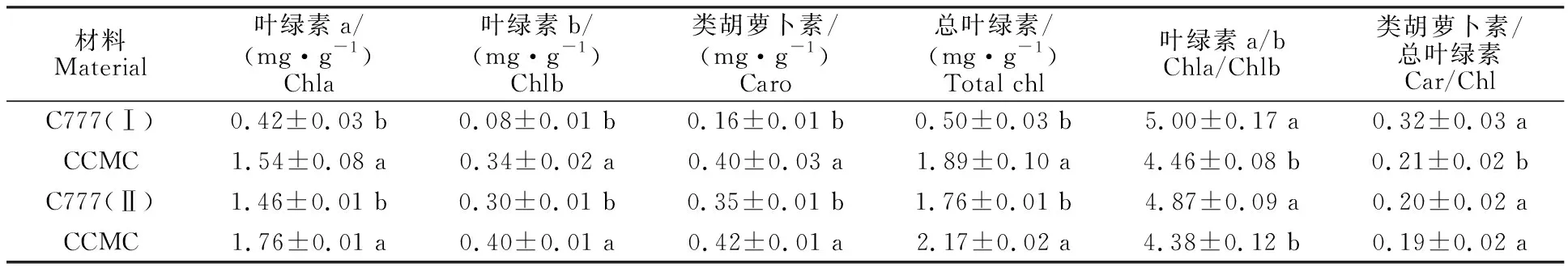
表1 黄瓜叶色突变体C777叶片光合色素质量分数及相对比值Table 1 Photosynthetic pigment mass fraction and relative ratio of leaf color mutant C777
注:数据为“平均数±标准差”。不同小写字母表示突变体与野生型差异显著(P<0.05)。下同。
Note:Data are “means±SD”.Different lowercase letters indicate significant difference between mutant and wild type(P<0.05). The same below.
2.2 光合速率及相关参数分析
由表2可知,在Ⅰ期和Ⅱ期,突变体的Pn均显著低于野生型,分别为野生型的58.68%和81.50%。推测在Ⅰ期由于突变体叶绿素质量分数低,使得其捕光能力降低,最终导致净光合速率低于野生型。Ⅱ期突变体叶片呈现黄绿色,叶绿素质量分数虽然有所上升,但净光合速率仍低于野生型。
突变体C777在 Ⅰ期和Ⅱ期,Ci显著高于野生型,分别高出81.41和47.13 μmol·mol-1。表明突变体对CO2的利用率较野生型低,这与两者之间Pn的表现一致。突变体Gs在Ⅰ期和Ⅱ期均比野生高,分别高出0.13和0.18 mol·m-2·s-1,而蒸腾速率在两者之间差异不显著。

表2 黄瓜叶色突变体C777叶片的光合特性Table 2 Photosynthetic characteristics of leaf color mutant C777 in cucumber
2.3 突变体叶绿素荧光参数及成像分析

由表3可知,突变体光化学猝灭系数(qP)显著低于野生型,为野生型的78.31%,说明突变体的PSⅡ系统将捕获的光量子转化为化学能的效率较野生型低。非光化学猝灭系数(NPQ)反映了植株以热能的形式耗散过剩光能的能力,突变体显著高于野生型,比野生型高出0.24,这说明突变体耗散过剩光能的能力较野生型强,通过耗散过剩的光能,突变体从而提高对自身光合系统保护的能力。突变体表观光合电子传递速率(ETR)显著低于野生型,为野生型的45.44%,说明突变体在电子传递效率上低于野生型,对光能的利用低于野生型,这与光合测定结果相符。
利用Fc800 FluorCam大型版开放式多光谱荧光成像系统检测突变体叶绿素荧光成像表型,发现突变体QYmax(即Fv/Fm)低于野生型(图1)。表明突变体由于叶绿素质量分数较野生型低,使得其PSⅡ的光能转换效率降低,成像分析结果与用叶绿素荧光仪所测结果相吻合。

表3 黄瓜黄绿叶突变体叶绿素荧光动力学参数Table 3 Chlorophyll fluorescence kinetic parameters of virescent leaf mutant in cucumber
3 讨 论
目前,黄瓜上报道的黄绿叶突变体相对较少,主要有苗晗等[16]报道的突变体9110Gt和王家训等[25]报道的一个黄绿叶突变株系ygl。9110Gt只在第4真叶以前转色明显,以后黄化和转色不突出[16];突变株系ygl真叶先呈黄绿色然后转为绿色[25],而本试验的黄绿叶突变体C777在整个苗期都表现出转色现象,且叶片先呈黄色然后转绿,最终为黄绿色,其表型不同于9110Gt和ygl,因此认为C777应是一个新的黄瓜黄绿叶突变体。
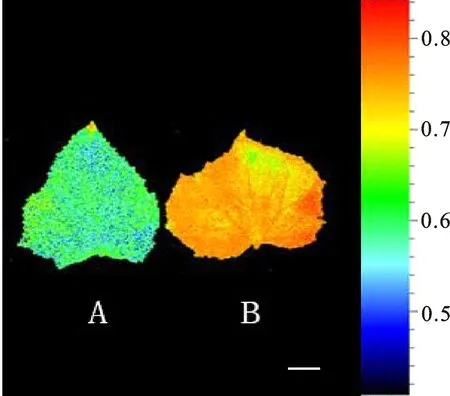
A.突变体 Mutant;B. 野生型 Wild type
在光合色素方面,苗晗等[16]研究表明,黄瓜黄绿叶突变体9110Gt转色前叶绿素及类胡萝卜素质量分数显著低于野生型,但转色后差异不显著,而本试验的黄绿叶突变体C777 转色前后的叶绿素及类胡萝卜素质量分数均显著低于野生型。此结果表明,C777的转色机制可能不同于同为转色突变体的9110Gt。本研究的黄绿叶突变体C777叶绿素a/b比值转色前后均显著高于野生型,说明突变体C777叶绿素a和叶绿素b之间的转换受到阻碍,使得叶绿素b的质量分数降低,此结果与其他学者在水稻、小麦等作物的黄化转绿突变体上的研究结果相同[11,26-27],而不同于黄瓜黄绿叶突变体9110Gt[19]。突变体C777为黄色时(转色前)类胡萝卜素/总叶绿素比值较野生型显著降低,而转为黄绿色后,其比值与野生型无显著差异,表明类胡萝卜素/与总叶绿素比值变化不会导致C777叶片黄化,而总叶绿素质量分数的降低和升高是造成C777黄化和转绿的主要原因。
光合色素的质量分数直接或间接地影响植物的光合能力,具体表现在净光合速率、蒸腾速率、胞间CO2摩尔分数等指标发生变化。在此方面,有关黄瓜黄绿叶突变体的研究还未见相关报道,本试验中突变体C777在 Ⅰ和Ⅱ期的净光合速率均显著低于野生型,胞间CO2摩尔分数显著高于野生型(表2),与李万青等[21]在黄瓜叶色黄化突变体C528上的研究结果一致。不同的是突变体C777的气孔导度在Ⅰ期和Ⅱ期均显著高于野生型,蒸腾速率之间无显著差异,说明突变体光合能力较低并非受气孔和蒸腾速率影响,而是由于对CO2的利用率较低所致。
叶片是植物进行光合作用的重要器官,光合作用效率的高低与叶片叶绿素质量分数以及叶绿体结构的完整性有密切的关系。黄瓜全基因组序列的发表和多种分子标记的应用,使得叶色突变体分子调控机制的研究更加深入,同时叶色突变体的研究价值得到了越来越多的关注。本试验的研究结果将在一定程度上为今后开展的该突变体的叶色突变机制、叶色基因的定位及克隆等相关研究工作奠定基础。
