茂兰喀斯特森林梓叶槭种群结构与数量动态
2018-12-05吴邦利龙翠玲秦随涛
吴邦利,龙翠玲,秦随涛
(贵州师范大学 地理与环境科学学院,贵阳 550001)
植物群落的基本特征是植物与植物、植物与环境之间的相互关系[1],而种群数量结构是植物种群生态学研究的重要内容,它能反映种群时间上的变动规律,体现了种群动态及其群落的演替趋势[2-3],其中生命表及根据生命表编制的存活曲线是研究种群结构及动态变化的重要工具,它能直观地展现种群各龄级的生长状态及存活趋势,揭示种群结构现状和更新策略,探索种群历史[4-7]。因此,研究群落特征及其种群数量动态对于了解群落未来的发展趋势具有重要意义。
梓叶槭(AcercatalpifoliumRehd.)为槭树科槭属的落叶乔木,为中国特有的珍稀濒危树种,国家二级保护植物,具有很高的利用和科研价值。其树形优美,树干高大,材质坚硬、致密,是四川西部城市理想的行道树或城市绿化庭园树种[8]。该树种已陷于濒危状态,由于大量的砍伐及不合理的毁林耕种,目前各产区极为罕见,现在主要分布于西南等地,零星散生于海拔500~1 300 m的亚热带常绿阔叶林中。梓叶槭在中国西南喀斯特地区森林生态系统中亦占据重要地位,位于贵州茂兰的喀斯特森林也是其分布地之一[9]。茂兰喀斯特森林是一类特殊的森林生态系统, 其群落性质为非地带性常绿落叶阔叶混交林,蕴藏着丰富的生物多样性。同常态地貌上的森林植被相比,在生态环境、森林群落性质、群落外貌、区系组成、演替更新动态、种群发展以及生态系统对环境的影响等方面都有显著不同[10],为地球同纬度带上仅存的原生性较强的喀斯特森林,具有重要的研究价值。茂兰喀斯特森林独特的小气候条件和高度异质多样的小生境,为梓叶槭等树种的生长提供了适宜的生存空间。当前,一些学者对梓叶槭的种子生物学特性[11]和种子萌发特性[8]等方面进行了研究。但关于茂兰喀斯特森林梓叶槭种群结构和数量动态方面未见系统的研究报道。有关梓叶槭种群在茂兰喀斯特森林不同地形部位生长状况和数量动态,梓叶槭种群所在植物群落的特征,喀斯特异质小生境对梓叶槭种群数量分布的影响等问题有待进一步研究。因此,本文拟通过对茂兰喀斯特森林不同地形部位森林群落的梓叶槭种群高度结构、年龄结构与数量动态等方面研究,掌握梓叶槭种群的生存现状和数量变化,揭示梓叶槭种群分布的动态规律及其影响机制,对于梓叶槭种群的合理保护与濒危机制的探索具有重要的意义,同时为茂兰喀斯特森林退化生态系统的恢复重建提供理论依据。
1 研究区概况和研究方法
1.1 研究区概况
研究地位于贵州省南部黔、桂交界处的茂兰国家级喀斯特森林自然保护区(107°52′~108°05′E,25°09′~25°20′N)。区内为典型的喀斯特峰丛地貌,最高海拔1 078.6 m,最低海拔430 m,平均海拔800 m以上。成土母岩以中下石炭纪白云岩及石灰岩为主。年均温为15.3 ℃,7月均温26.4 ℃,1月均温8.3 ℃,≥10 ℃以上积温5 727.9 ℃,年均降雨量1 320.5 mm。全年平均相对湿度83%。土壤以黑色石灰土为主,土层浅薄,地面岩石裸露,pH 7.5~8.0,有机质和全氮含量丰富[12]。据统计,不同地形部位植物群落皆含有常绿和落叶成分,且垂直分布格局变化不明显,山脊地段生境严酷,却不乏常绿树种生存,亦有很多落叶树种在阴湿的洼地、漏斗底部等负地形中繁衍,落叶树种是植物对严酷的喀斯特生境相适应的结果。喀斯特生境由于地表水分缺乏,各种旱生植物发育良好,加上土壤中富钙及富盐基化作用,该群落的树种多为耐旱、喜钙的种类。如圆果化香(Platycaryalongipes)、黄连木(Pistaciachinensis)、圆叶乌桕(Sapiumrotundifolium)、云贵鹅耳枥(Carpinuspubescens)、南天竹(Nandinadomestica)等。
1.2 研究方法
1.2.1样方设置和数据采集在实地勘察的基础上,结合研究内容,在茂兰自然保护区选取坡地、槽谷、漏斗3种地形部位的典型森林群落,分别设置10 m×10 m不连续样地10块,共30块。采用GPS对各样地的海拔和地理坐标进行测量,并记录各样地的坡度、坡向等数据。对各样地的乔灌木进行每木检测,记录种名、胸径、高度、冠幅、郁闭度等因子。此外,在样地中随机设置5个1 m×1 m 的小样方,调查草本层的种名、数量、盖度等内容。以朱守谦等对茂兰喀斯特森林进行大量样地调查所确定的最小表现面积900 m2[9,13]作为依据,每个地形部位的样地总面积为1 000 m2。
1.2.2梓叶槭种群年龄结构的建立梓叶槭材质坚硬,不易钻取木芯,生长年龄也很难确定,故本研究用立木级结构代替年龄结构分析其种群结构和群落特征,大小结构按2种方式处理:胸径(DBH)在2.5 cm以下的个体分2级,胸径大于2.5 cm的个体分4级。根据以上划分标准,把30块样地中的梓叶槭种群个体分类如下[14-15]:Ⅰ级(幼苗),DBH < 2.5 cm,H < 33 cm;Ⅱ级(幼树),DBH < 2.5 cm,H ≥ 33 cm;Ⅲ级(小树),2.5 cm ≤ DBH < 7.5 cm;Ⅳ级(中树),7.5 cm ≤ DBH < 15 cm;Ⅴ级(大树),15 cm ≤ DBH <22.5 cm。根据种群的生长和高度特点,株高0~2 m为Ⅰ级,2~4 m为Ⅱ级,4~6 m为Ⅲ级,每个等级之间的间隔为2 m,共划分为7个等级。
1.2.3各群落类型重要值的计算重要值(importance value,IV)是判定群落各层次优势种和划分群落类型的常用指标。其计算公式如下[16]:重要值IV=(DR+PR+FR)/3,式中,DR为该种在群落中的相对密度,PR为该种在群落中的相对显著度,FR为该种在群落中的相对频度。
1.2.4静态生命表和存活曲线编制静态生命表又称特定时间生命表,多用于对长寿命的木本植物种群的统计研究,种群的静态生命表对于种群动态研究具有重要意义,根据梓叶槭各龄级的统计株数,按照特定时间生命表的编制方法[17-18],计算生命表中各参数,完成梓叶槭种群特定时间生命表的编绘,其中,ax:x龄级内出现的个体数,a0:Ⅰ龄级个体数;lx:标准化存活数,lx=ax/a0×1 000;dx:x龄级到x+1龄级间隔期内标准化死亡数,dx=lx-lx+1;qx:x龄级到x+1龄级间隔期内死亡率,qx=dx/lx;Lx:x龄级到x+1龄级间隔期间平均存活的个体数,Lx=(lx+lx+1)/2;Tx:x龄级及其以上各龄级存活的个体总数,Tx=∑Lx;ex:进入x龄级个体的生命期望寿命,ex=Tx/lx;Sx:存活率,即x+1期存活个体数与x期存活个体数之比,Sx=lx+1/lx;Kx:致死力或消失率,即从x到x+1期受到的阻力,Kx=ln(lx)-ln(lx+1)。以龄级为横坐标,以标准化存活数为纵坐标,绘制梓叶槭种群存活曲线,以死亡率、消失率和期望值为纵坐标,以龄级为横坐标绘制死亡率曲线、消失率曲线和期望值曲线,分析种群动态变化。数据处理和相关图表制作在Excel 2010和SPSS 20.0中完成。
2 结果与分析
2.1 群落的分布特征
茂兰国家级自然保护区内梓叶槭各样地的基本概况见表1。由表1可知:在茂兰国家级自然保护区,梓叶槭主要分布在3种不同地形的群落类型,即漏斗森林、槽谷森林、坡地森林,其海拔主要集中分布在500 m~900 m、坡度为5°~60°之间的低山区域。不同地形部位森林群落的乔木层、灌木层都有优势种分布,林分郁闭度大且林下植物种类丰富。
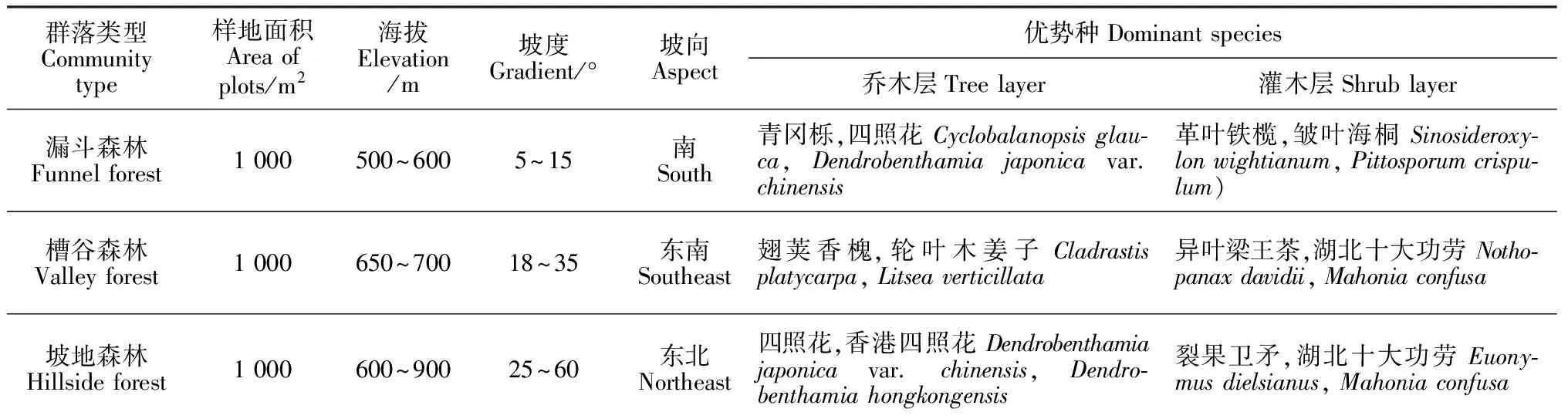
表1 茂兰喀斯特森林梓叶槭群落类型的样地基本信息及其优势种
2.2 群落特征分析
2.2.1漏斗森林漏斗森林林下岩石露头多,土壤为黑色石灰土,土被不连续,低凹处土层厚度为5~20 cm,水分条件好,且郁闭度较大的某些地段有积水现象,但光照条件差,林分垂直结构复杂。该类型样地乔木层优势种为壳斗科的青冈栎和山茱萸科的四照花,重要值分别为2.337和1.809(表2),构成青冈栎-四照花群落(Cyclobalanopsisglauca-Dendrobenthamiajaponicacommunity);共存乔木有桂皮(Cinnamomumtamala)、圆叶乌桕、狭叶润楠(Machilusrehderi)、朴树(Celtissinensis)、香港四照花(Dendrobenthamiahongkongensis)、齿叶黄皮(Clausenadunniana)、尾叶罗伞(Brassaiopsisproducta)和樟叶槭(Acercinnamomifolium)等种类。灌木层优势种为海桐花科的皱叶海桐,重要值为1.322,伴生种有革叶铁榄、石岩枫(Mallotusrepandus)、南天竹、湖北十大功劳、六月雪(Serissajaponica)、粗丝木(Gomphandratetrandra)和鼠李(Rhamnusdavurica)等。
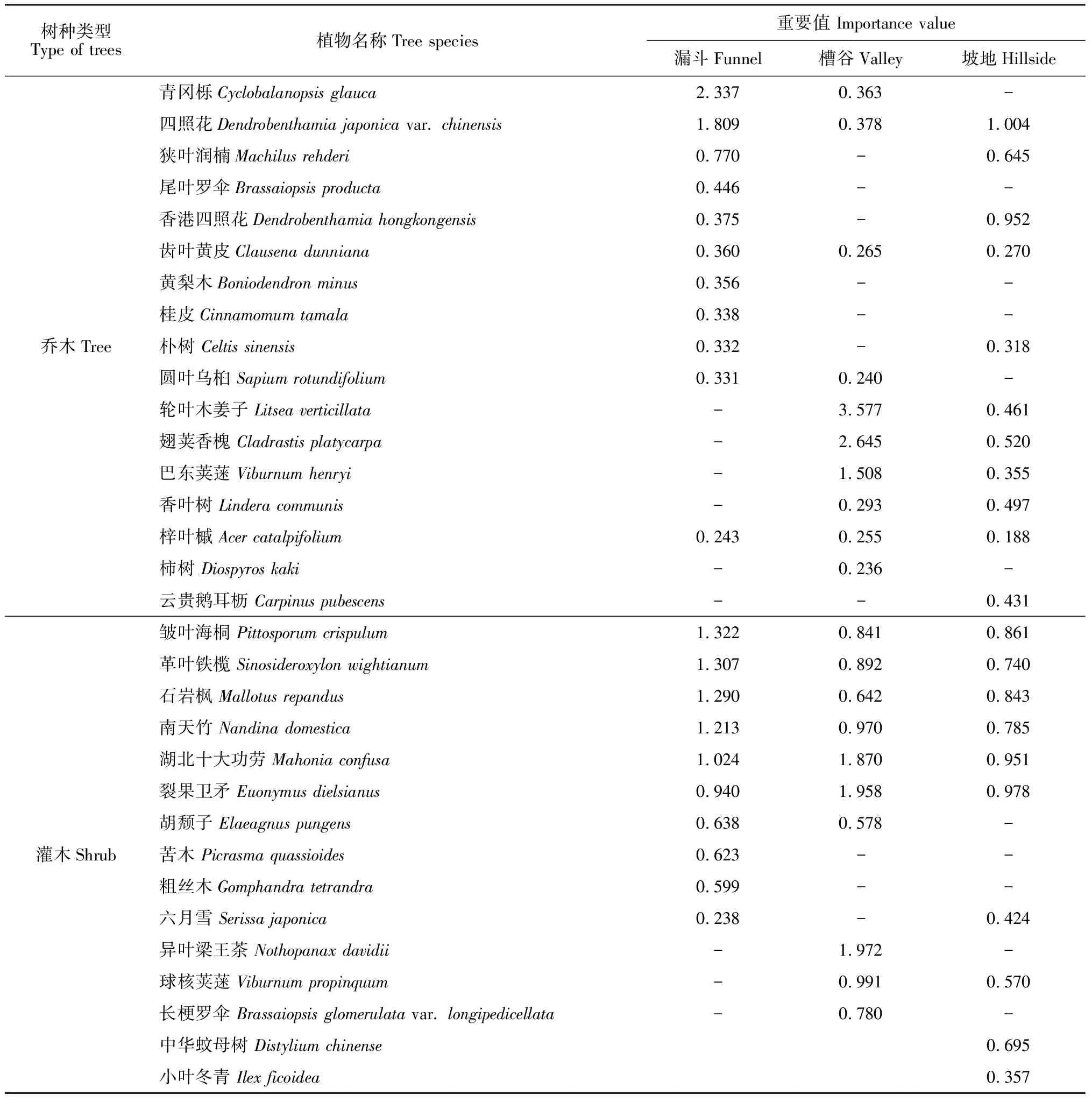
表2 茂兰喀斯特森林不同森林群落优势种
2.2.2槽谷森林槽谷森林位于下坡坡位,谷底地势平坦,林下岩石露头多,土壤为黑色石灰土,土被不连续,土层厚度为2~6 cm,水分、光照条件适中,乔木层分布不均匀。该样地乔木层优势种为樟科的轮叶木姜子和豆科翅荚香槐,重要值分别为3.577和2.645(表2),构成轮叶木姜子-翅荚香槐群落(Litseaverticillata-Cladrastisplatycarpacommunity),乔木共生种有巴东荚蒾(Viburnumhenryi)、香叶树(Linderacommunis)、柿树(Diospyroskaki)、四照花和青冈栎等种类。灌木层优势种为五加科的异叶梁王茶,重要值为1.972 ,伴生种有南天竹、长梗罗伞(Brassaiopsisglomerulatavar.longipedicellata)、胡颓子(Elaeagnuspungens)、吴茱萸(Evodiarutaecarpa)、皱叶海桐、球核荚蒾(Viburnumpropinquum)、湖北十大功劳和火棘(Pyracanthafortuneana)等。
2.2.3坡地森林坡地森林位于坡体中上部,地势较陡,林内岩石露头多,具大面积岩石崩塌碎块,土壤为黑色石灰土,土层浅薄且不连续,低凹处土层厚度为1~3 cm,水分条件差,光照条件好,林分垂直结构简单。该样地乔木层优势种为山茱萸科的四照花和香港四照花,重要值分别为1.004 和0.952 (表2),构成四照花-香港四照花群落(D.japonica-D.hongkongensiscommunity),乔木层共生种有狭叶润楠、轮叶木姜子、翅荚香槐、云贵鹅耳枥、多脉榆(Ulmuscastaneifolia)、黄连木、巴东荚蒾等种类。灌木层优势种为卫矛科的裂果卫矛,重要值为0.978,伴生种有革叶铁榄、中华蚊母树(Distyliumchinense)、扁核木(Prinsepiautilis)、水麻(Debregeasiaorientalis)、小叶冬青(Ilexficoidea)、紫珠(Callicarpabodinieri)等。
2.3 种群结构分析
2.3.1种群高度结构对不同群落类型中的梓叶槭种群进行高度级划分对于揭示种群个体在群落中的作用具有重要意义,由统计结果(图1)可知,梓叶槭数量主要集中在Ⅰ(0~2 m)、Ⅱ(2~4 m)、Ⅲ(4~6 m)3个级,占总个体数的93.26%左右,其中Ⅰ级比例高达49.65%,其次依次为Ⅱ、Ⅲ和Ⅳ,而达到Ⅶ级(>12 m)仅有1株,占0.03%。说明梓叶槭种群幼苗幼树较多,高大植株较少,种群整体偏矮小,呈现出个体数随高度的增长而逐渐递减的趋势。不同地形部位群落梓叶槭种群高度级的分布规律基本相同(图2)。在槽谷群落中,梓叶槭数量在前4位高度级结构中占绝对优势,但大于Ⅴ级(8~10 m)的个体仅有1株;漏斗群落的梓叶槭数量虽不如槽谷,但种群的高度级结构分布较为均匀;坡地是3种群落中梓叶槭分布最少的群落类型,且高度主要集中在Ⅰ~Ⅳ之间,无高大植株分布。3类森林群落梓叶槭高度结构虽各有特点,但从整体上看,其高度分布是连续的,呈现逆J字型的高度结构。
2.3.2种群年龄结构对漏斗、槽谷、坡地森林群落的梓叶槭种群数量进行统计,结果(图3)显示,不同生境Ⅰ级幼苗均严重缺乏,分别占其总个体数的15.6%(槽谷)、11.89%(漏斗)、15.09%(坡地);Ⅱ级幼树和Ⅲ级小树在槽谷、漏斗、坡地群落中均有分布,其中,Ⅱ级幼树数量最多,所占比例分别为44.9%、35.4%、19.7%,Ⅲ级小树所占比例分别为48.6%、27%、24.3%。漏斗生境梓叶槭种群径级结构最为完整,幼苗、幼树、小树、中树、大树数量分别占总数11.89%、62.38%、16.83%、6.93%、1.98%,槽谷和坡地生境径级结构与漏斗生境相比较不完整,据统计数据显示,槽谷梓叶槭数量最多,但主要集中在Ⅰ~Ⅲ级,无中、大树分布;坡地种群数量最少,且无大树分布。从整体上看,Ⅰ级幼苗向Ⅱ级幼树发育过程中死亡率较高,表明梓叶槭种群自然更新情况相对较差,致使种群结构不稳定;此外,梓叶槭种群Ⅲ径级数量急剧下降,分别较Ⅱ径级数量减少了65%、73%、74%,表明Ⅱ~Ⅲ级这一阶段梓叶槭生存率较低。综上所述,3种生境梓叶槭种群为中间大两头小的径级结构,具有一定的更新能力,属稳定型种群。如后续幼苗数量补充不足,种群逐渐向衰退型转变。

Ⅰ.树高0~2 m;Ⅱ.树高2~4 m;Ⅲ.树高4~6 m;Ⅳ.树高6~8 m;Ⅴ.树高8~10 m;Ⅵ. 树高10~12 m;Ⅶ.树高>12 m;下同图1 茂兰喀斯特森林梓叶槭种群高度级结构Ⅰ.0~2 m; Ⅱ.2~4 m; Ⅲ.4~6 m; Ⅳ.6~8 m; Ⅴ.8~10 m; Ⅵ. 10~12 m; Ⅶ.>12 m; The same as belowFig.1 The height structures of A. catalpifolium in Maolan Karst Forest
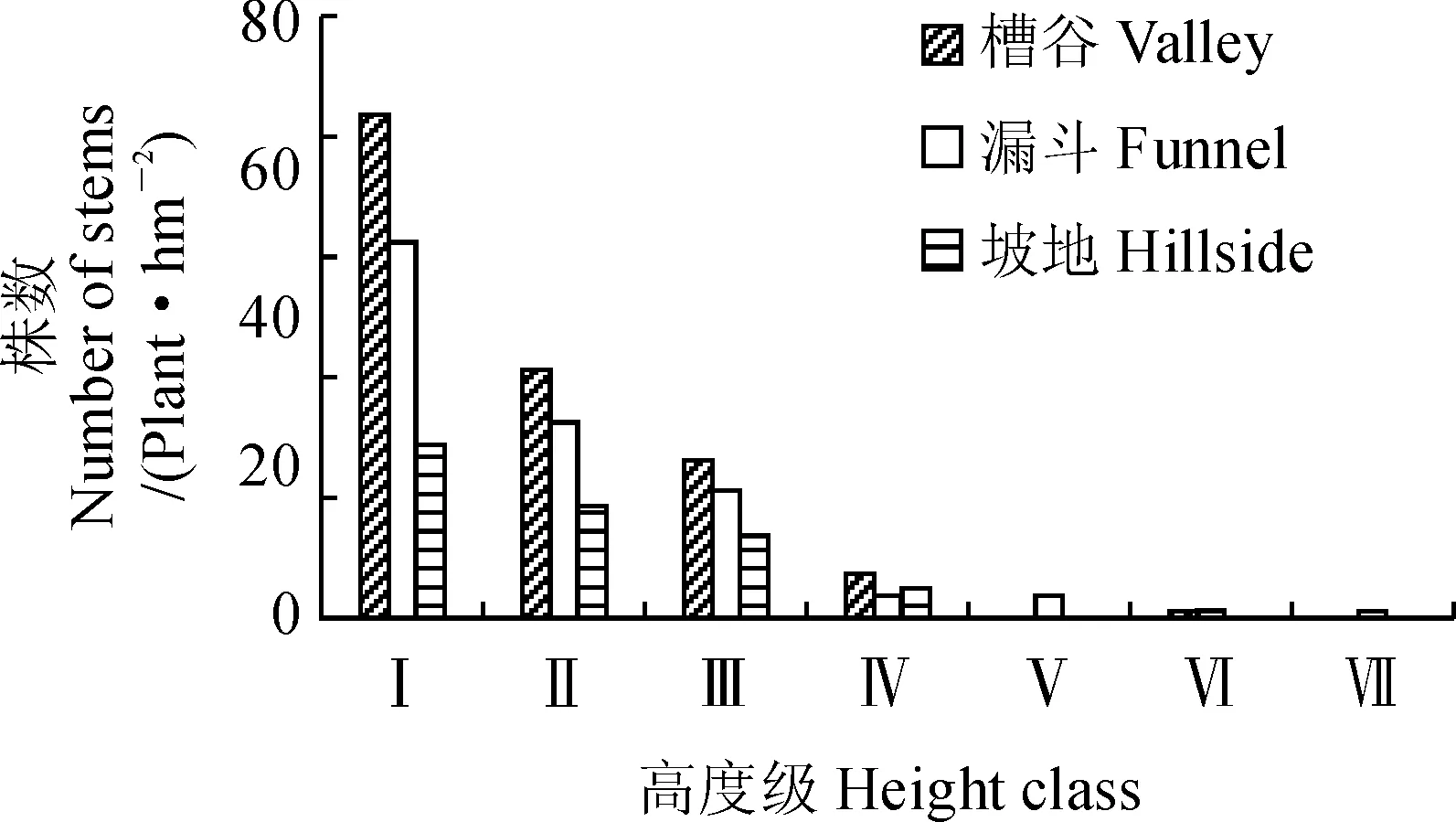
图2 茂兰喀斯特森林不同地形梓叶槭群落的高度级结构Fig.2 The height structures of A. catalpifolium at different community types in Maolan Karst Forest
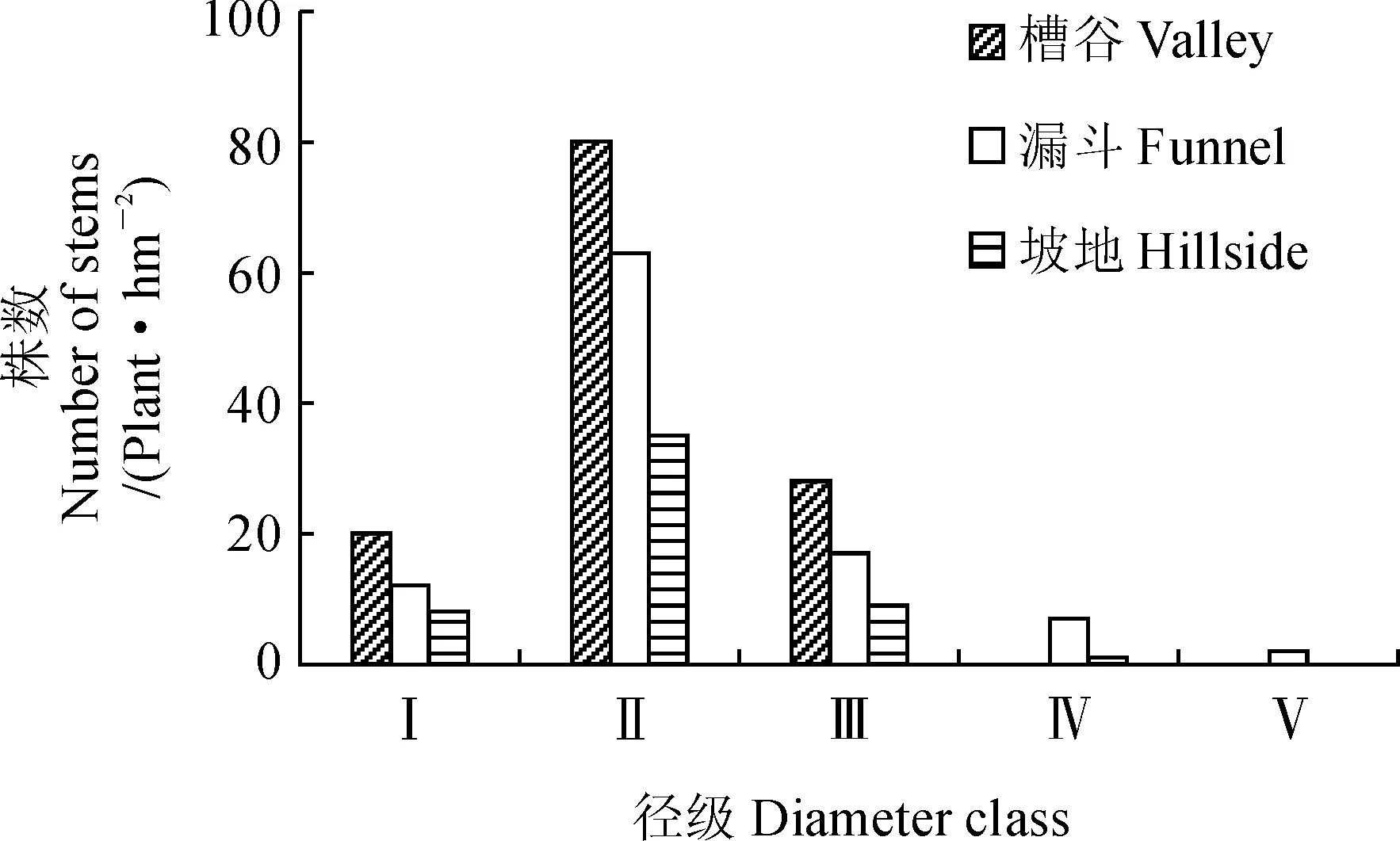
Ⅰ. 胸径< 2.5 cm,树高< 33 cm;Ⅱ. 胸径 < 2.5 cm,树高≥ 33 cm;Ⅲ. 2.5 cm ≤ 胸径< 7.5 cm;Ⅳ. 7.5 cm ≤ 胸径 < 15 cm;Ⅴ. 15 cm ≤ 胸径 <22.5 cm图3 茂兰喀斯特森林不同地形梓叶槭群落的种群大小结构Ⅰ.DBH < 2.5 cm,H < 33 cm;Ⅱ. DBH < 2.5 cm,H ≥ 33 cm;Ⅲ. 2.5 cm ≤ DBH < 7.5 cm;Ⅳ. 7.5 cm ≤ DBH < 15 cm;Ⅴ. 15 cm ≤ DBH <22.5 cmFig.3 Size structures of A. catalpifolium population at different topography sites in Maolan Karst Forest
2.4 种群静态生命表和存活曲线分析
不同生境梓叶槭种群存活数(除Ⅰ径级)随着径级增加而降低(表3),死亡率(qx)和致死力(Kx)曲线变化趋势基本一致(图4),随着径级增加大致呈上升趋势,主要是小径级个体的株高、冠幅较小,生存空间充足,随着径级增长和冠幅扩大,个体间对水分、光照及养分竞争激烈,导致qx和Kx不断上升;此外,Ⅰ径级在不同生境的死亡率均为负数,说明其幼苗严重不足。梓叶槭种群平均期望寿命(ex)总体上随径级增加而逐渐下降,这是因为径级越小,群落个体间竞争降低,能有效利用群落资源,从而ex值较大。除漏斗生境ex在Ⅲ径级略有回升外,其他径级均呈下降趋势,主要是梓叶槭种群达到一定年龄后生理机能开始退化,一直到种群活力最终消失,这种趋势符合种群生态学特征,反映梓叶槭种群整个生命过程中个体生命力的衰退。
存活曲线是对生命表的重要反映,其能反映种群存活率或死亡率随年龄的变化情况[17,19]。梓叶槭的存活曲线表明梓叶槭Ⅰ级幼苗存活数极低,从Ⅱ级幼树开始存活率逐渐降低而死亡率显著升高(图4)。总体上来说,3种生境条件下的梓叶槭种群的存活曲线介于DeeveyⅡ型和DeeveyⅢ型之间,根据Hett和loucks检验估算存活情况时采用2种数学模型,即指数方程式NX=N0e-bx用以描述DeeveyⅡ

表3 茂兰喀斯特森林不同地形梓叶槭种群静态生命表

图4 茂兰喀斯特森林不同地形梓叶槭种群的存活数(lx)、寿命期望(ex)、死亡率(qx)和致死力(Kx)曲线Fig.4 Survival(lx),life expectancy(ex), mortality(qx)and killing power(Kx ) curves of A. catalpifolium population at different topography sites in Maolan Karst Forest
型存活曲线;幂函数式NX=N0x-b描述DeeveyⅢ型存活曲线[20-21],经拟合建立其相关模型得到:
槽谷:NX=5 317.2e-0.579x(F=0.057,R2=0.054,P<0.05)
NX=2 707.1x-1.109(F=0.164,R2=0.141,P<0.05)
漏斗:NX=1 268.4e0.1682x(F=3.337,R2=0.527,P<0.05)
NX=1 326.3x0.4887(F=1.361,R2=0.312,P<0.05)
坡地:NX=5 916.1e-0.76x(F=1.616,R2=0.447,P<0.05)
NX=2 372x-1.24(F=0.696,R2=0.258,P<0.05)
根据检验结果可以判断梓叶槭种群除在槽谷部位指数模型的F值和决定系数R2值稍低幂函数模型外,漏斗和坡地的指数模型F值和决定系数R2值均高于幂函数模型的F值和决定系数R2值,且P值均小于0.05,因此,梓叶槭存活曲线更趋于DeeveyⅡ型,属稳定型种群。
3 讨 论
植物种群数量动态不仅反映种群内不同个体的组配情况,也反映了种群的结构、发展趋势与环境之间的相互关系,是揭示种群生存现状和更新策略的重要途径之一[22-23]。本研究以大小级结构代替龄级结构分析茂兰喀斯特森林梓叶槭种群结构及其动态。从研究结果可知,不同地形部位森林群落均严重缺乏Ⅰ径级个体,Ⅱ径级个体占较大的比例,中、大径个体较少,表明梓叶槭种群幼苗在发育过程中死亡率较高,如果没有幼苗个体对幼树的补充,梓叶槭种群的稳定性将难于维持,随着时间的推移,种群逐渐向衰退型转变。这种径级结构与其他濒危保护植物,如天宝岩长苞铁杉种群、准噶尔盆地梭梭种群、星斗山水杉种群等的径级结构相似[20,24-25]。但与贵州其他槭树科槭属不同,覃弦等[26]在茂兰喀斯特森林不同演替阶段优势乔木种群结构及数量动态一文中提到樟叶槭演替早期、中期和晚期的存活曲线均接近DeeveyⅢ型,基本处于增长型向稳定型过渡的阶段。刘海燕等[27]研究了贵州特有植物贵州槭的种群结构与动态,发现贵州槭种群中小径级个体较多,大径级个体数量少,种群存活曲线为DeeveyⅢ型,属增长型结构。本研究中,梓叶槭在群落中并非优势种,其受到的种间竞争较为激烈,加之茂兰喀斯特生境的严酷性和土壤营养空间缺乏,造成梓叶槭生长速度缓慢。梓叶槭在3种不同森林群落生长差异明显,其中,槽谷梓叶槭数量最丰富,坡地数量最少,这与梓叶槭的生长特性有关。余道平等[11]研究表明,温度对梓叶槭种子的萌发率有显著的影响,温度过高或过低均能影响种子活力,造成发芽和出苗不良。槽谷森林地势平坦、水热条件均衡,适宜梓叶槭幼苗幼树的生长,而坡地森林坡度大、地势陡峭、光照强、缺乏水分等条件造成种子生产与更新困难。同时,槽谷和坡地森林与漏斗森林相比径级结构不完整,出现径级缺失的现象(主要缺失中大径个体),这主要是因为槽谷和坡地生境土层较薄,再则由于岩石的崩塌,造成树干折断、倒伏等机械损伤,使病原菌入侵造成林木心腐,不利于梓叶槭中大树的生长。此外,严重的人为干扰及动物对种子的采食也是影响种群更新的原因。因此,梓叶槭种群的衰退与严酷的喀斯特生境、人为干扰有关。
静态生命表反映了多个世代的年龄动态历程中的一个特定时间,对深刻分析种群的现状、动态并预测未来具有重要意义[17,28-29]。梓叶槭为天然种群,并且静态生命表是以时间推空间,故在编制过程中会出现死亡率为负的现象,这虽与静态生命表的数学假设不符,但仍能作为重要的生态学依据反映种群在迅速的发展或是衰落之中[2,30]。本研究表明,梓叶槭种群Ⅰ径级在漏斗、槽谷、坡地的死亡率(qx)和致死力(Kx)等均为负值,说明梓叶槭种群缺乏幼苗,由于幼苗发育过程中易受病虫害侵扰,加之喀斯特生境的破碎化,导致梓叶槭幼苗幼树死亡率上升,这与种群结构方面的分析是一致的。梓叶槭种群存活数随着径级增加而降低,且生命期望单调下降,存活曲线接近于DeeveyⅡ型,种群死亡率和致死力曲线变化趋势基本一致,致死力在槽谷、漏斗第Ⅱ级、漏斗第Ⅳ级、坡地第Ⅲ级出现峰值。该种群在第Ⅱ级向第Ⅲ级转化过程中死亡率较高,是由于幼年个体以高死亡率为代价向成年阶段转换。在喀斯特地区,地表岩石裸露、土层浅薄,蓄水能力差,水分是植物幼苗生长的必要条件,而喀斯特森林的土壤水分不能满足所有植株的生长需要。土壤水分条件相对较差导致种内和种间竞争激烈,空间生态位受到限制[24,30]。第Ⅳ级出现峰值是由于梓叶槭个体为争夺空间和光照资源而导致激烈的种内竞争产生自疏现象,是导致中大径个体数量减少的原因之一。总之,梓叶槭种群生存动态与喀斯特自然环境及自身生态学特性有关。这是前期薄弱、后期衰退的濒危植物种群的一种现象[19]。
梓叶槭是中国特有的珍稀树种,颇具科研和用材价值,同时也是较好的园艺观赏树种,具有广阔的开发前景,因此在梓叶槭的保护上,可以采取以下措施:一是立足于现有的喀斯特生境,对梓叶槭进行就地保护,确保种群有足够的生存空间;二是在梓叶槭生长的区域,减少人为活动的干扰破坏,并适当采取人工干预的措施,如疏伐群落内过于密集的藤本、灌木,适度的干扰将有利于梓叶槭种群的散布并改善林内小环境,降低外部因素对种群幼苗的影响,从而增加梓叶槭的数量,促进种群的天然更新;三是补充低龄级个体,如补植幼苗、幼树等;四是加强科学研究,提高种子向幼苗的转化率。因此,对茂兰喀斯特森林梓叶槭群落特征及种群数量动态的研究,有助于了解种群的生长现状和发展趋势。同时,对于该种群濒危机制的探索和研究奠定理论基础,为梓叶槭种群的天然更新及退化喀斯特区森林生态系统的恢复与重建提供理论依据。
