西南喀斯特生态系统氮素循环特征及其固碳效应
2018-11-28李德军陈浩肖孔操张伟王克林
李德军,陈浩,肖孔操,张伟,王克林
(1. 中国科学院亚热带农业生态研究所,亚热带农业生态过程重点实验室,湖南 长沙 410125;2. 中国科学院环江喀斯特生态系统观测研究站,广西 环江 547100)
氮是构成蛋白质和核酸的物质基础,因此,一切生命体都需要氮素。然而,由于生物圈中储存的生物可利用性氮的量非常有限[1],陆地自然生态系统植物的生长普遍受氮素限制[2]。生态系统中的氮绝大部分是以有机化合物的形态存在。由于有机化合物分子中的碳与氮原子存在一定的比例关系,这就决定了生态系统不同组分,如林冠、树干、凋落物、微生物量及土壤有机质中的碳与氮含量之间存在一定的比例范围,或化学计量学关系[3]。碳、氮之间的计量关系在很大程度上决定了生态系统的碳、氮循环密切耦合[3-4]。这一点在全球变化背景下具有重要意义。一方面,陆地生态系统固碳被认为是从大气中吸收二氧化碳,进而减缓全球变暖的一条经济而有效的途径,然而,由于碳、氮循环之间紧密的耦合关系,陆地生态系统固碳在很大程度上受制于土壤的供氮能力[5-6]。在氮状况较好的区域,土壤供氮能力相对较强,生态系统的固碳潜力相对较高;反之,生态系统的固碳潜力则较低[6]。另一方面,由于氮是植物生长的必需元素,也是主要的限制性元素,因此,土壤氮的供应能力在很大程度上决定着生态恢复的成效[7]。在氮状况好的区域,土壤氮供应能力较强,植物生长更好,重建的生态系统就更容易或能更快发展到演替的顶级阶段;反之,重建的生态系统可能长期停留在演替的早期阶段[7]。可见,评估生态系统的氮状况对预测全球变化背景下生态系统的固碳潜力及生态演替的方向与进程均具有重要的意义。
然而,目前对区域尺度生态系统氮状况的认识仍非常有限。通常认为温带及北方生态系统受氮限制,而热带与亚热带成熟森林则表现出天然氮饱和特征[8]。然而,这些观点主要是基于一些间接、零散的证据,而直接与系统性的证据缺乏[9]。
喀斯特生态系统是地表系统的重要组成部分,约占全球陆地面积的10%及我国国土面积的36%[10-11]。我国的喀斯特生态系统尤以西南喀斯特区最为典型,这一区域也是全球三大喀斯特集中连片分布区之一。喀斯特生态系统具有其特殊性,如土壤钙/镁含量高、基岩裸露率高、土层浅薄且分布不连续、具有发达的裂隙管道系统等[12]。后二个特征导致喀斯特生态系统非常脆弱,一旦遭到干扰和破坏,水土流失严重,进而引发土地退化。事实确实如此,过去由于不合理的人为活动特别是农业耕作,大面积的喀斯特生态系统遭受了不同程度的退化,甚至石漠化[10]。近20年来,我国在西南喀斯特山区实施了一系列生态恢复工程(如退耕还林还草、石漠化综合治理等),石漠化扩展趋势得到了遏制,生态恢复也取得了不同程度的效果[13]。然而,由于工程实施先行而机理研究滞后,我们至今不清楚喀斯特山区生态恢复的主要限制性因素是什么。如前所述,氮是多数陆地自然生态系统植物生长的主要限制元素,也是决定生态恢复成效的关键元素,因此,西南喀斯特山区生态恢复工程亟需回答的关键科学问题是:氮是否为喀斯特区生态恢复的主要限制性元素?其次,喀斯特生态系统具有一系列的特殊性,其氮状况与邻近的非喀斯特生态系统相比有何差异?我们近几年针对上述问题开展了比较系统的研究,以下将从二方面进行简要论述。
1 退耕后喀斯特土壤氮状况随演替变化特征及其固碳效应
1.1 基于小流域尺度退耕演替序列或不同退耕模式的证据
基于小流域尺度(古周生态恢复示范区和木论国家级自然保护区)退耕后次生演替序列(包括草丛、灌丛、次生林和原生林),以农田作为对照,采用空间代替时间的方法研究了氮状况随演替变化特征[7,14]。结果表明,退耕后土壤(表层15 cm)总氮随演替较快累积,经过70年左右恢复到原生林水平,平均累积速率约为12 g N/(m2·yr)[7]。由于碳-氮循环密切耦合,氮的累积也相应保障了退耕后土壤有机碳随演替进程快速累积,经过40年左右能恢复到原生林水平,平均累积速率约为138 g C/(m2yr)[15]。我们发现退耕后土壤有机碳和总氮累积遵循以下模式(公式1)[7,15]:

式中:St0和St分别表示退耕前(t0)及退耕后某一时间点(t)土壤碳(或氮)储量,A表示土壤碳(或氮)储量在退耕前与达到平衡态之间的差值;B代表增长常数;yrs表示退耕后恢复年限;C表示形态参数。创新性的提出了土壤碳(或氮)储量达到平衡态所需时间(T,单位为年)的估算方法(公式2)[7,15]:
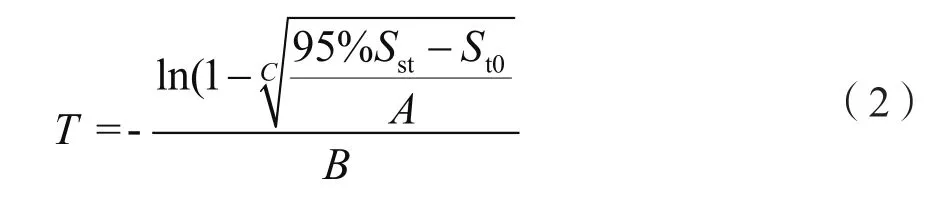
式中:Sst表示平衡态时土壤碳(或氮)储量。
土壤总氮快速累积的一个重要前提是需要有充足的外源氮输入。大气氮沉降与生物固氮是多数陆地自然生态系统外源氮输入的二条主要途径[16]。我们发现这二条途径仅能解释土壤氮累积速率的40%~76%[7]。进一步分析发现基岩中含有一定量的氮,在基岩快速风化溶蚀过程中,岩石中的氮释放到土壤中,进而被土壤直接固定或被植物吸收后随凋落物进入土壤并成为土壤总氮组分[7]。上述三个氮源可解释土壤氮累积速率的40%~102%[7]。可见,岩石氮释放是解释喀斯特山区退耕后土壤总氮累积的一条重要机制[7]。Houlton等[17]基于全球表层岩石的分析也表明西南喀斯特山区碳酸盐岩氮含量相对较高。另有研究则表明西南喀斯特山区岩石的化学风化速率居于全球高水平[18]。较高的氮含量和风化速率无疑是西南喀斯特山区碳酸盐岩氮释放速率相对较高的主要原因。
然而,土壤氮的累积是外源氮输入与不同途径土壤氮流失综合作用的结果,因此,仅有外源氮的输入并不足以解释退耕后土壤氮的快速累积[7]。在土壤对氮的截留/固持能力较弱的情况下,外源氮进入土壤后将很快通过不同途径流失,并不会导致土壤氮的净累积。因此,退耕后土壤总氮能快速累积意味着喀斯特土壤具有较高的氮固持能力。经过深入分析,我们发现土壤总氮与交换性钙及镁含量之间存在非常强的线性关系[7]。由于土壤总氮90%以上为有机氮,因此,我们提出土壤有机质通过钙镁离子与矿物表面生成矿物复合体是喀斯特土壤有机质稳定的重要途径,进而有助于土壤有机氮(或总氮)累积[7]。这也是解释退耕后随演替进程土壤总氮快速累积的另一重要机制[7]。
土壤总氮的累积导致土壤中不同形态无机氮含量随演替进程而发生变化[7]。在草丛阶段以铵态氮为主,而在其他阶段则以硝态氮为主。土壤中的硝态氮与铵态氮之间的比值是反映土壤氮状况的一个重要指标,比值小于1意味着生态系统缺氮,而大于1则意味着生态系统富氮。基于该指标我们提出在草丛阶段缺氮,而其他阶段则富氮。土壤15N比值也支持灌丛、次生林和原生林土壤氮循环相比草丛阶段更开放[7]。叶片N∶P比值也是反映生态系统氮状况的一个常用指标,该比值大于16表示植物受磷限制,小于14表示受氮限制,而在二者之间意味着植物受氮和磷同时限制[19]。我们对不同演替阶段植物群落叶片N∶P比值的分析表明,草丛阶段植物受氮限制,灌丛阶段受氮和磷同时限制,而次生林和原生林阶段植物受磷限制[14]。此外,土壤初级氮转化也是评估土壤氮状况的有效方法。基于氮同位素稀释法,我们定量了土壤初级氮矿化速率(GNM)、初级硝化速率(GN)、硝态氮异化还原为铵的速率(DNRA)、微生物对铵态氮固持速率(GAI)和微生物对硝态氮固持速率(GNI)。GNM和GN在退耕后早期速率下降,之后随演替进程而增加;但其他三个过程速率随演替进程没有明显的变化规律。GN∶GAI比值是反映土壤氮状况的一个重要指标,该比值小于1意味着生态系统缺氮,而大于1则意味着生态系统富氮。基于该比值的分析,我们发现草丛阶段微生物受氮限制,而后续阶段则表现出明显的氮饱和特征。
进入土壤中的凋落物所含的氮主要为蛋白质类物质,包括蛋白质、几丁质及肽聚糖等大分子多聚体,其中蛋白质所占的比例最高[20]。对于这些多聚体微生物无法直接利用,必须经过胞外酶的解聚作用变成小分子有机氮后微生物才能利用,进而经过氮转化过程生成铵态氮和硝态氮[20-21]。这些生成的小分子有机氮、铵态氮和硝态氮均比较容易被植物吸收利用或从土壤中流失。基于此,我们假定二种可能的情景。第一种情景是进入土壤中的凋落物绝大部分经过解聚及初级氮转化后生成的小分子有机氮、铵态氮和硝态氮,进而被植物吸收或从土壤中流失。在这种情景下,退耕后随演替进程土壤总氮的累积速率理论上应非常低。第二种情景是进入土壤中的凋落物仅有小部分经过解聚及初级氮转化后生成的小分子有机氮、铵态氮和硝态氮。在这种情景下,退耕后随演替进程土壤总氮的累积速率理论上应比较高。通过分析参与土壤大分子多聚体解聚过程的几种重要的、与氮转化相关的胞外酶,我们发现蛋白水解酶活性与初级氮矿化速率显著相关,表明蛋白质解聚环节是初级氮矿化作用的限速步骤。蛋白水解酶活性也与土壤总氮含量显著相关。由于蛋白水解酶活性理论上与被解聚的多聚体呈正相关关系,因此,蛋白水解酶活性与土壤总氮含量之间显著的相关关系意味着被解聚的多聚体有机质与土壤总氮含量呈正比例关系,即次生演替过程中随着土壤总氮增加,不同演替阶段的土壤总氮按一定比例被解聚成小分子有机氮。基于此,我们认为多聚体有机质解聚过程受限制也是解释演替过程中土壤总氮累积的一条重要机制。
基于不同退耕恢复模式的研究也支持退耕后土壤总氮与有机碳能较快累积,但退耕模式对总氮与有机碳累积速率具有重要影响[22-23]。长期退耕模式试验地位于广西壮族自治区环江毛南族自治县古周村小流域,包括三种退耕模式,即种植经济树种香椿、种植牧草桂牧一号及任豆与桂牧一号混种,以未退耕的玉米-大豆轮作地为对照。退耕16年后,所有退耕处理的有机碳水平均得到了显著提升,但种植香椿处理的土壤有机碳增加最显著;种植香椿和任豆/桂牧一号混种二种退耕模式下土壤总氮与总磷水平得到了显著提升;基于磷脂脂肪酸(PLFAs)分析的结果表明,三种退耕模式下土壤微生物及其不同功能群丰度均显著增加[22]。
1.2 基于区域尺度的证据
为了探明基于小尺度演替序列的结论是否适用于区域尺度,我们在桂西北区域开展了不同岩性下土壤碳、氮随演替或土地利用变化的研究,包括三种岩性(白云岩、石灰岩和碎屑岩)和五种土地利用类型(耕地、草丛、灌丛、次生林和人工林),其中草丛、灌丛和次生林代表退耕后不同演替阶段[24-28]。总体上,土壤有机碳和总氮随退耕后演替进程而增加[24-25],这与基于小尺度退耕演替序列的结果一致。然而,进一步的分析发现,岩性对土壤有机碳和总氮随退耕后演替的变化特征具有显著影响[24-25,27]。在白云岩区,自然生态系统转变为耕地后土壤总氮与有机碳流失相比石灰岩区更严重;反过来,白云岩区退耕后土壤总氮与有机碳累积相比石灰岩区更明显[24-25]。造成白云岩与石灰岩区土壤总氮与有机碳随土地利用变化存在差异的主控因素为土壤交换性钙水平。如前所述,交换性钙是决定喀斯特土壤有机质稳定的重要因素,交换性钙水平的高低一定程度上决定着土壤有机碳和总氮的水平[7,15]。一方面,白云岩和石灰岩的溶解速率存在很大差异,后者是前者的3-60倍[29]。另一方面,喀斯特耕地土壤侵蚀速率相比其他土地利用类型较高[30,31]。在白云岩区,由于耕地土壤侵蚀过程中钙流失速率大于岩石溶解过程中钙的释放速率,导致耕作过程中土壤钙含量下降严重,土壤总氮与有机碳水平相应下降;而石灰岩区由于岩石溶解过程中钙释放速率高,耕作过程中流失的钙能得到及时补充,土壤钙含量在不同土地利用类型之间差异低于白云岩。退耕后,土壤侵蚀速率下降,钙的淋失速率相应降低。在白云岩区,土壤中的钙通过岩石溶解释放而逐渐得到补充,土壤总氮与有机碳相应增加;而在石灰岩区,土壤中钙由于在耕作过程中维持比较高的水平,因此,退耕后变化不大,土壤总氮与有机碳含量变化幅度不如白云岩区[25]。进一步的分析发现,岩性不仅影响土壤有机碳含量,也影响有机碳的稳定性;但退耕后的演替仅影响土壤有机碳含量[26]。
植树造林被视为增加土壤固碳、减缓全球变暖的有效途径[5]。然而,目前关于造林影响土壤有机碳库变化的内在机理的认识尚不充分。为了探明植树造林对土壤碳、氮和磷含量的影响是否受岩性的调节,在典型喀斯特区采用配对采样的方式,分别在石灰岩和碎屑岩区采集了农田和人工林表层土壤样品(0~15 cm),分析了土壤有机碳、全氮和全磷含量[27]。结果表明,造林对土壤有机碳和氮库的影响受岩性调节,但对土壤全磷的影响不受岩性调节。相比石灰岩区,在碎屑岩区造林短期内对土壤有机碳库的提升效果更明显。相反,在石灰岩区,造林导致土壤氮库显著下降;而在碎屑岩区,造林对土壤氮库没有显著影响。在两种岩性区,造林均会导致土壤磷库的下降。此外,在两种岩性区造林都会引起土壤碳、氮和磷元素的摩尔比增加,这表明造林后土壤固碳会逐渐受土壤氮和磷的限制。进一步的分析发现,在二种岩性条件下,造林后土壤稳定性碳组分在总有机碳中的比例均显著下降,意味着造林后土壤有机碳稳定性下降。我们的研究表明植树造林对土壤有机碳和氮库的影响受岩性调节,为解释造林对土壤碳和氮库在不同区域之间的差异化影响提供了新的机制[27]。
通过对无机氮(DIN,铵氮+硝氮)、可溶性有机氮(DON)、净矿化速率和净硝化速率的分析,我们发现土壤硝氮、总可溶性无机氮、净矿化速率和净硝化速率均随植被正向演替(即从草丛到次生林)而增加[28]。土壤NO3-∶NH4
+比与DIN∶DON比在草丛阶段与1无明显差异,之后随演替进程而线性增加,在灌丛与次生林阶段(特别是后者)上述比值均显著大于1。因此,基于区域的研究支持基于小尺度演替序列研究得到的结论,即在草丛阶段缺氮,而在次生林阶段则表现出明显的氮饱和特征[28]。
土壤微生物养分限制性分析也是评估生态系统氮、磷养分状况的重要依据[32-35]。通过对桂西北峰丛洼地区域不同演替阶段生态系统土壤胞外酶活性化学计量特征的分析表明,喀斯特生态系统土壤微生物主要受碳和磷的限制,而不受氮限制。进一步分析表明,微生物受碳限制程度在不同演替阶段或岩性条件下存在差异,农田和森林最高,而草丛最低;白云岩区域比石灰岩区域微生物受碳限制程度更高[32]。微生物磷限制随演替变化特征也受岩性影响,在白云岩区域,微生物受磷限制程度从农田至次生林逐渐增强,但在石灰岩区域受磷限制程度在不同演替阶段之间无明显差异[32]。
2 喀斯特与非喀斯特森林氮状况比较
土壤初级氮转化是反映森林氮状况的重要指标。我们前期的研究表明退耕之后喀斯特生态系统土壤总氮快速累积,在森林阶段表现出明显的氮饱和特征。但是仅针对喀斯特生态系统的研究无法判断氮饱和是否是喀斯特森林的独有特征。基于此,我们采用氮同位素稀释法研究了典型喀斯特森林及其邻近红壤区的非喀斯特森林初级氮转化特征[36]。除铵态氮同化速率外,其他测定的初级氮转化速率喀斯特森林均高于非喀斯特森林。非喀斯特森林的初级氮矿化速率较低而铵态氮同化速率与喀斯特森林相似,导致非喀斯特土壤产生的铵态氮能被较好固持。同时,生成的硝态氮大多被DNRA和GNI二个过程有效固持,导致非喀斯特森林硝态氮净产生速率非常低。而在喀斯特森林,生成的铵氮仅有小部分被固持,加上硝化速率高而固持速率低,导致土壤硝氮净生成速率较高。非喀斯特森林的氮饱和指数(GN∶GAI)与1无显著差异,但喀斯特森林则远高于1,意味着喀斯特森林表现出氮饱和特征,而邻近的非喀斯特森林则受氮限制[36]。土壤酶活性化学计量关系能反映土壤微生物对碳、氮和磷的需求状况,从而间接反映土壤养分的可利用状况[34-35]。基于此,我们研究了典型喀斯特森林及其邻近的非喀斯特森林土壤酶活性化学计量特征[33]。基于向量长度的分析表明,喀斯特森林土壤微生物受碳限制程度高于非喀斯特森林。基于临界C∶N比值的分析表明,喀斯特森林土壤微生物受碳限制,非喀斯特森林土壤微生物受氮限制。基于临界C∶P比值和向量角度的分析,发现喀斯特与邻近非喀斯特森林土壤微生物均受磷限制,但非喀斯特森林受磷限制程度高于喀斯特森林[33]。基于区域尺度土壤酶活性化学计量关系的研究表明在区域尺度上喀斯特森林土壤微生物普遍受碳限制,而不受氮限制[32]。
3 结论
我们的研究表明西南喀斯特山区退耕后土壤总氮能比较快速累积,退耕后土壤可利用氮水平随演替进程而增加,喀斯特森林表现出明显的氮饱和特征,而邻近的非喀斯特森林则表现出氮限制特征。我们的研究意味着西南喀斯特山区退耕后的生态恢复仅在早期短时间受氮限制,而中后期则不受氮限制;充足的氮供应可保障生态恢复工程的固碳效应。喀斯特森林氮循环具有其独特性,在全球变化背景下,其响应与适应性也可能异于其他区域/类型森林。
