Synergy and Redundancy in a Signaling Cascade with Different Feedback Mechanisms∗
2018-11-24LiFangWang王莉芳YingXu徐莹JunMa马军andYaJia贾亚
Li-Fang Wang(王莉芳),Ying Xu(徐莹),Jun Ma(马军),and Ya Jia(贾亚),†
1Department of Physics and Institute of Biophysics,Central China Normal University,Wuhan 430079,China
2Department of Physics,Lanzhou University of Technology,Lanzhou 730050,China
3NAAM-Research Group,Department of Mathematics,Faculty of Science,King Abdulaziz University,P.O.Box 80203,Jeddah 21589,Saudi Arabia
AbstractFeedback plays an important role in various biological signal transmission systems.In this paper,a signaling cascade system(including three layers:input(S),intermediate(V),output(X)components)is employed to study thefluctuations and net synergy in information transmission,in which the V component is regulated by itself or the X component,and each feedback on V is either positive or negative.The Fano factor,the net synergy,and the signalto-noise ratio(SNR)of signaling cascade with the four possible feedback types are theoretically derived by using linear noise approximation of the master equation,and the ability of information transmission through the signaling cascade is characterized by using the partial information decomposition of information theory.It is found that the signaling cascade exhibits different responses to the four feedback mechanisms,which depend on the relationships between degradation rates of components.Our results not only clarify the dependence of the Fano factor,net synergy,and SNR on the feedback regulations with the varying of degradation rates of components,but also imply that living cells could utilize different feedback mechanisms to adapt to the external fluctuating environments.
Key words:stochastic theory,biological signaling transmission cascade,feedback
1 Introduction
Living cells can respond to variations of intracellular or extracellular signals,which behave as information transmission processes.These biochemical reaction networks consist of numerous regulatory motifs where the signal cascades include various feedback mechanisms,and the biological messages are transmitted in the networks of signal cascades,[1−2]such as the gene regulatory network,[3−5]the neuronal network,[6−7]the protein-protein interaction network[3,8−9]etc.It is well known that each biochemical reaction is highly stochastic,which leads to fluctuations in biochemical networks[10−13]and phenotypic diversity of clonal cells.[11,14−16]
Recently,the information transmission of biochemical regulatory motifs had been extensively investigated.Shannon’s information theory provides a methodology for quantifying the reliability of information transmission systems.[17]Williams et al.[18]introduced a non-negative decomposition of multivariate mutual information terms,and showed that partial information forms a lattice,which illustrates the general structure of multivariate information.An in-depth theoretical analysis of partial information decomposition(an extension of information theory)was proposed by Barrett et al.[19]It was demonstrated that the approach is applied to the measures of information transfer and information-based measures of complexity in a neuroscience contexts.
In order to understand how cells deal with the fluctuating environment[20−23]and how cells transduce signals via different network motifs in the presence of noise,Ronde et al.[24−25]analyzed how two binary input signals can be encoded in the concentration of a common signaling protein,and the two output signals can provide reliable information about the inputs in the presence of noise.It was clarified that in the signaling cascade with both autoregulation and feedback,the gain-to-noise ratio dependence on frequency is illustrated using the mathematical model and simulation.Selimkhanov et al.[26]demonstrated that signaling dynamics plays an important role in overcoming extrinsic noise by analyzing the ability of temporal signal modulation,and showed that signaling dynamics can mitigate and potentially eliminate extrinsic noise.It was found that extrinsic noise induces information loss through experimental measurements of information transmission in a signal regulated kinase network.The limits on information transmission in signal pathways was studied through a combination of quantitative experiments and theoretical analysis by Hansen et al.[27]It was demonstrated that the amount of information transmitted by the yeast transcription factor Msn2 to single target genes is limited,but information transmission can be increased by adjusting promoter cis-elements or by integrating in-formation from multiple genes in the presence of noise.
Although there are a number of theoretical and experimental investigations about information transduction of signal modules,little has been dealt with synergy and redundancy in information transmission in a signaling cascade with different feedbacks in the presence of noise.The synergy and redundancy in information transmission are related to many biological phenomena and processes in biological system,such as the activities of neurons,[28−30]eye color,[31]the behaviors of animals and biological processes responsible for disease.[30−34]However,the fluctuations in the signaling cascade can not be neglected since biochemical reaction networks are always in changing environments.Maity et al.[35]demonstrated that the network motifs with different types of regulation affect the degradation rate dependence of the noise and mutual information differently.Biswas et al.[36]studied the signal transmission in a generic two-step cascade using a mathematical model by varying the degradation rate of the input component.Moreover,Pedraza et al.[37]found that thefluctuations in each component are affected by the mean value of the component and its life time.Some experimental evidences[38−39]suggested that degradation time at the single cell level can be measured and adjusted,and stochastic protein expression in an individual cell was investigated by experiments.[40−41]Furthermore,feedback and autoregulation motifs play very important roles in biochemical systems,[39,42−45]for example,buffering noise,improving sensitivity,enhancing adaptability,and changing the response time of the system.The above investigations indicated that feedback strategy and degradation rate can affect the kinetic characteristics of biochemical reaction networks.However,previous researches did not involve the effects of degradation rates of components on synergy and redundancy of information transmission in the signaling cascade with various feedbacks.
To study the stochastic nature of biochemical interaction processes,one has to use the mesoscopic approaches,instead of the macroscopic description wherefluctuations are neglected.The first approach is to solve the probability distribution of all the different molecular components[46]by using the chemical master equation.The famous Gillespie algorithm is usually used to perform stochastic simulations to obtain numerical results of chemical master equation.[47]The second technique is the linear noise approximation(LNA)of the chemical master equation,which is used in this paper.By applying this technique,the chemical master equation is simplified to the Fokker-Planck equation via utilizing van Kampen’s expansion,[46]and the statistical features of stochastic systems are characterized[48−49]rapidly.The third approach is the Langevin method that can obtain a solution for small fluctuations around steady state.
Interesting questions now arise:What are the effects of degradation rate of the signaling cascade with different feedbacks on the fluctuations and information transmission?How do different types of feedback influence synergy and redundancy in the information transmission in the presence of fluctuations? What is the relationship between net synergy and the SNR of the system in information transmission?
Based on the models[19,25,35−36]of information transmission,in this paper,a signaling cascade model with four feedback types is employed to investigate the fluctuation effects and information transmission by using LNA method.The article is organized as follows.In Sec.2,we present a signaling cascade with different feedback types,and then the Fano factor and net synergy are derived via utilizing the linear noise approximation.In Sec.3,we discuss the performance of the signaling cascade involving in different feedbacks.We end with conclusions of the paper in Sec.4.
2 Model and Method
We consider the information transmission through a signaling transmission cascade with different feedbacks as shown in Fig.1,where there are three components:the input component S,the intermediate component V,and the output component X.In gene transcription regulatory networks,transcription factors can be regarded as the signals,and regulate its target gene.Therefore,here S can act as an input signal(e.g.,the transcription factor)that regulates the output signal X(e.g.,the target gene)by the intermediate component V.
In the deterministic case,the kinetic process of the signaling cascade can be described in the following dimensionless form:

where s,v,and x are the expression level(or concentration)of components S,V,and X,respectively.The two terms on the right side of Eqs.(1)–(3)describe all the reactions involving in the production events and degradation events of S,V,and X,respectively.The parametersµs,µvand µxdenote the degradation rate of S,V,and X,respectively.
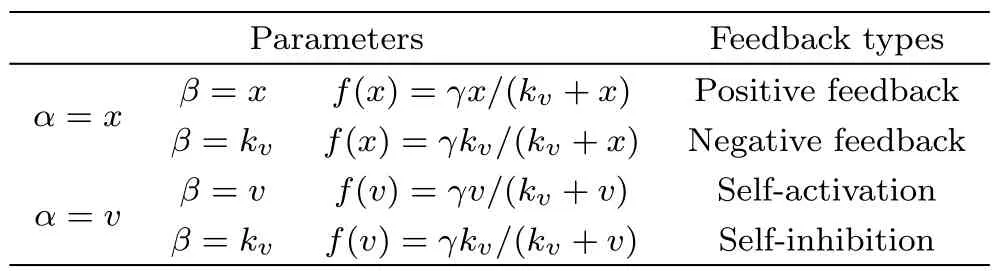
Table 1 Four freedback mechanisms.
Note that f(α)in Eq.(2)is a function in the form of f(α)=(γβ)/(kv+α),in which the relationships between parameters α and β can represent four feedback mechanisms as listed in Table 1.The positive and negative feedbacks correspond to the scheme(a)in Fig.1,and the self-activation and self-inhibition feedbacks correspond to the scheme(b)in Fig.1,respectively.
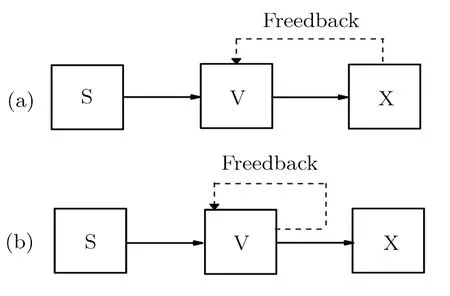
Fig.1 General schematic diagram of the signaling cascade with(a)the feedback(positive or negative)of output component X and(b)the self-feedback(activation or inhibition)of intermediate component V.
To characterize the fluctuationproperties due to finite molecular numbers of components S,V,and X,we introduce joint probability distribution P(Ns,Nv,Nx,t),where Ns,Nv,and Nxrepresent the numbers of components S,V,and X at time t,and are expressed by Ns=Ωs,Nv=Ωv,Nx=Ωx.The parameter Ω denotes the size of system.The chemical master equation[46,50−51]corresponding to Eqs.(1)–(3)obeys the following form

By utilizing van Kampen’s Ω-expansion method,[46]the number of the i-th component(i=s,v,x)is approximated asand the joint probability distribution is given by P(N1,N2,...,NR,t)=Ω−R/2Π(ξ1,ξ2,...,ξR,t).The Fokker-Planck equation is obtained by collecting the terms of Ω0in the expansion of Eq.(4)and written as

in which Aijis the stationary Jacobian matrix of Eqs.(1)–(3),and Bijis the stationary diffusion matrix:

To obtain the values of variance and covariance of different components of the signaling cascade,the Lyapunov equation at steady state is given by[48,52]
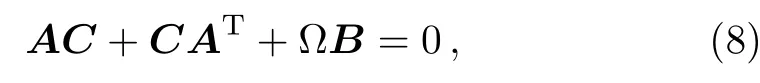
where the matrix C encompasses the variances and covariances,which represent the stochastic fluctuations in the numbers of components S,V and X and correlation degree of the fluctuations among components of the system,respectively.Substituting Eqs.(6)and(7)into Eq.(8),the matrix C can be obtained as follows


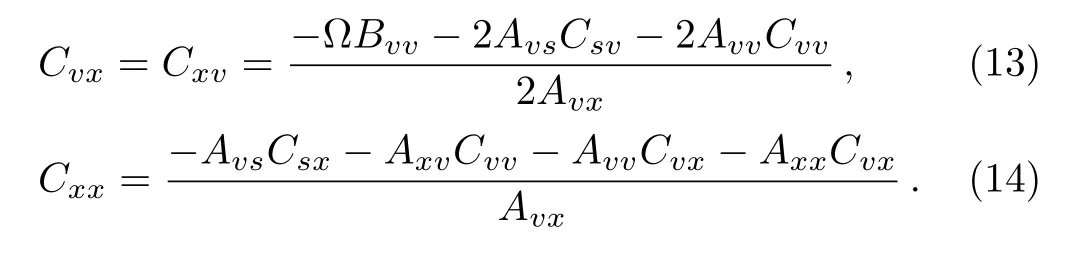
2.1 Fano Factor
The Fano factor,which is a measure of the relative size of the internal fluctuations in the system,[53]is defined as the ratio of the variance to the mean.The Fano factors of component S,V,and X are expressed as

2.2 Net Synergy
According to Refs.[18–19,27,36],a partial information decomposition(PID)is adopted in the paper.The information I(s;v,x)is composed of four terms:(i)The unique information that V provides about S:U(s;v|x);(ii)The unique information that X provides about S:U(s;x|v);(iii)the redundant information that both V and X provide about S:R(s;v,x);(iv)The synergistic information that the combination of V and X provides about S:S(s;v,x).The mathematical expressions of these quantities are expressed as I(s;v,x)=U(s;v|x)+U(s;x|v)+R(s;v,x)+S(s;v,x),I(s;v)=U(s;v|x)+R(s;v,x),I(s;x)=U(s;x|v)+R(s;v,x),and then the net synergy is written as∆I(s;v,x)=I(s;v,x)−I(s;v)− I(s;x).Thus,the net synergy is represented as∆I(s;v,x)=S(s;v,x)−R(s;v,x),in which the relationship between synergy and redundancy is given.I(s;v,x),I(s;v),and I(s;x)denote the mutual information between three components S,V,and X,quantifying the amount of information one component includes about another,and characterize the information transfer of the system between the source(S)and the targets(V and X).Consequently,the net synergy can be rewritten as
where conditional variances of the system are given by

2.3 Signal-to-Noise Ratio
To measure the fidelity of information transmission of the signals,signal-to-noise ratio(SNR),which is taken as a measuring method to quantitatively evaluate the fidelity of information transmission in the signaling pathway,[54−55]is defined as

3 Results
In order to clearly demonstrate the impacts of feedback regulation on the noise and net synergy,we consider four different settings of the degradation rates of components:(i)µs< µv:µs=0.1,µv=1;(ii)µs≪ µv:µs=0.1,µv=10;(iii)µs= µv:µs=1,µv=1;and(iv)µs> µv:µs=1,µv=0.1.For these cases,one can study the stochastic fluctuations of components around equilibrium state in the signaling cascade.Thus,the mean expression level of these components S,V,and X are assumed to keep at a certain value,respectively.Similar assumptions were previously made,especially when analyzing some real systems.[56−57]
In this paper,we set γ =10,ks= µss,and kx=(µxx)/v in the following four cases.The setting of parameter kvis as follows:(i)Positive feedback α=x and β =x:kv=(xγs− xvµv)/(vµv);(ii)Negative feedback α =x and β =kv:kv=(vxµv)/(γs − vµv);(iii)Selfactivation α =v and β =v:kv=(γs − vµv)/µv;(iv)Self-inhibition α =v and β =kv:kv=(v2µv)/(γs−vµv).
Here,the net synergy and mutual information are measured in bits.Note that in the following sections,the case ofµs≪ µvis not considered in the negative feedback and self-inhibition.This makes sense because the production of the intermediate component V is repressed strongly by the joint interaction between fast degradation rateµvof V and the suppression arising from X or V itself.
3.1 Effects of Degradation Rates on Fano Factorof Output Component X for DifferentFeedbacks
By virtue of the theoretical expressions derived in Sec.2,the Fano factor of output component X as a function ofµxis depicted in Fig.2.In most cases,it is found that with the increasing of the degradation rate of output componentµx,the Fano factor of X increases for very slowµx,and saturates to a plateau value for fastµx.However,in the case ofµs≪ µv,the Fano factor of X is independent of the change ofµxfor self-activation,referring to the red dash dotted line in Fig.2(c).
In the case ofµs= µv,for positive feedback and selfactivation,the minimum Fano factor value of X is achieved whenµs≪ µv,and the maximum Fano factor value of X is obtained whenµs>µv,corresponding to Figs.2(a)and 2(c).For negative feedback and self-inhibition,the system exhibits different responses to the variations of the degradation rates of components.For instance,at a given value ofµx,when µs< µv,the Fano factor of X is large and becomes small whenµs> µv,as shown in Figs.2(b)and 2(d).
The above results indicate that in the case ofµs> µv,for positive feedback and self-activation,significant relative fluctuations in the output component X are generated and the rapid variation of input signal S can be sensed by output component X.For negative feedback and self-inhibition,although the input component S is at a faster degradation rate compared with the intermediate component V,the fluctuations in the output component X are suppressed and X is not sensitive to the change of input signal.These results are in accord with some closely related phenomena in biological systems,such as osmo adaptation,[58]bacterial chemotaxis,[43]and the responses of S.Cerevisiae to the changes in external carbon source.[59]
In the case ofµs= µv,for positive feedback and self-activation,at a given value ofµx,the fluctuations in the output component X are comparatively smaller than those in the cases ofµs< µvand µs> µv,but larger forµs≪ µv.By contrast,for negative feedback and self-inhibition,when µs= µv,the fluctuations in X are at an intermediate level between the other two cases,i.e.,µs< µvand µs> µv.The results of Fig.2 suggest that the fluctuations in the output component X can be influenced by the input signal and other intermediate components through correlation intensity of the fluctuations,which can be adjusted by various feedback regulation.Therefore,cells can take advantage of appropriate responses to adapt to changing environments when facing challenges caused by random fluctuations.

Fig.2 (Color online)Effects of degradation rates on Fano factor of output component X for different feedbacks:(a)Positive feedback α =x and β =x.(b)Negative feedback α =x and β =kv.(c)Self-activation α =v and β =v.(d)Self-inhibition α =v and β =kv.
3.2 Characteristics of Information Transmission in Signaling Cascade with Different Feedbacks
(i)The Case of Positive Feedback α =x and β =x
For positive feedback,the dependence of the net synergy and mutual information on degradation rates of components are plotted in Fig.3.In the case ofµs< µv,the net synergy∆I(s;v,x)is a monotonically decreasing function ofµxfor slow µx,and then it tends to a constant value asµxcontinues to increase for fastµx,as illustrated in Fig.3(a).Moreover,the net synergy∆I(s;v,x)<0,which demonstrates that redundant property of the target variables is dominant in the signaling cascade with positive feedback.It is observed from Fig.3(e)that I(s;v,x)is almost independent of the change ofµx.Meanwhile,the mutual information I(s;x)displays an opposite tendency with the variation ofµxin contrast with the net synergy,and∆I(s;v,x)≈−I(s;x),implying that the net synergy is mainly determined by the mutual information I(s;x)between the input component S and the output component X.
In the case ofµs≪ µv,it is found from Fig.3(b)that the change tendency of net synergy∆I(s;v,x)with the alteration ofµxis similar to that in the case ofµs< µv,but with∆I(s;v,x)>0,which reflects that synergistic feature of the target variables is prevalent over the redundancy in information transmission.According to Fig.3(f),it is seen that I(s;v,x)is decreased by increasingµx.Meanwhile,with the increasing ofµx,the mutual information I(s;x)increases at beginning for very small values ofµx,reaches a peak value,and then decreases to a constant for large values ofµx,showing that there exists an optimal value ofµxat which the maximum value of mutual information I(s;x)can be obtained in such a case.
In the case ofµs= µv,with the increasing ofµx,the net synergy∆I(s;v,x)exhibits an approximately threshold behavior and the threshold is aroundµx=0.83,as shown in Fig.3(c).This signifies that redundant feature of the targets plays a leading role in information transduction forµx>0.83 due to∆I(s;v,x)<0,but forµx<0.83,the net synergy∆I(s;v,x)>0.Figure 3(g)shows that there is a peak in the curve of I(s;v,x)and I(s;x)is a monotone increasing function ofµx.
In the case ofµs> µv,Fig.3(d)shows that the net synergy also displays an approximately threshold behavior as a function ofµx,and the threshold is aboutµx=4.66.Forµx>4.66,the net synergy is slightly reduced with increasingµxand then reaches a constant value for fastµx.In Fig.3(h),it is obvious that the variations of mutual information I(s;v,x)and I(s;x)with the change ofµxis analogous to that inµs=µv.However,the change of the net synergy is confined to a smaller range compared with those in the other three cases,since the mutual information is less in this case.
(ii)The Case of Negative Feedback α =x and β =kv
For negative feedback,the influence of degradation rates of three components on the net synergy and mutual information are illustrated in Fig.4.It is found that forµs< µv,the changes of the net synergy and mutual information are similar with increasingµxin Figs.3(a)and 4(a).However,the absolute value of net synergy is slightly smaller in the latter than that in the former case at the same value ofµxsince the net synergy∆I(s;v,x)is mainly determined by the mutual information I(s;x).The mutual information I(s;x)shown in Fig.3(e)is more than that in Fig.4(d)for a given value ofµx.
In Fig.4(b),when µs= µv,the net synergy is a monotone decreasing function ofµx,without showing a threshold behavior,and has a faster decline than that in Fig.3(c),which is attributed to the difference in the mutual information between Figs.3(g)and 4(e).That is,the faster the degradation rateµx,the larger the absolute value of net synergy.

Fig.3 (Color online)Effects of degradation rates on the net synergy and mutual information for positive feedback α =x and β =x.(a)and(e): µs< µv.(b)and(f): µs ≪ µv.(c)and(g): µs= µv.(d)and(h):µs> µv.

Fig.4 (Color online)Effects of degradation rates on the net synergy and mutual information for negative feedback α =x and β =kv.(a)and(d): µs< µv.(b)and(e): µs= µv.(c)and(f): µs> µv.
From Fig.4(c),it can be found that when µs> µv,the net synergy also displays an approximately threshold behavior with the change ofµx,and the threshold is at aboutµx=0.63.Moreover,forµx>0.63,the net synergy is reduced significantly by increasingµx,unlike the case of Fig.3(d).Figure 4(f)shows that the mutual infor-mation I(s;v,x)and I(s;x)are less in this case,leading to the small variation range of net synergy compared with the other two cases of negative feedback.Our results reveal that positive feedback and negative feedback always play different roles in the signaling cascade,and can act as the effective mechanisms of modulating noise and net synergy by varying the degradation rates of components in changing environments.
(iii)The Case of Self-Activation α =v and β =v
According to the above analysis,a natural question is whether these results of feedback regulation still hold in the case of autoregulation.Hence,in the following sections,we will explore how the degradation rates of components affect information transmission in the signaling cascade with self-activation or self-inhibition.The numerical results are plotted in Figs.5 and 6.
Figure 5 shows that the net synergy and mutual information as a function ofµxfor self-activation.Forµs< µv,the changes of net synergy∆I(s;v,x)and mutual information by alteringµxare similar to those in the case of positive feedback,suggesting that redundant property of the targets is dominant in this case once again.Forµs= µv,the variation trend of net synergy and mutual information by varyingµxare analogous to the case of positive feedback.However,the peak value of net synergy is larger compared to positive feedback.
Fig.5 (Color online)Effects of degradation rates on the net synergy and mutual information for self-activation α =v and β =v.(a)and(e):µs< µv.(b)and(f):µs≪ µv.(c)and(g):µs= µv.(d)and(h):µs> µv.

Fig.6 (Color online)Effects of degradation rates on the net synergy and mutual information for self-inhibition α =v and β =kv.(a)and(d):µs< µv.(b)and(e):µs= µv.(c)and(f):µs> µv.
Forµs≪ µv,the changes of the net synergy and mutual information differ from those in the case of positive feedback.From Fig.5(b),it is observed that the net synergy is decreased monotonically with an increase inµx.Specifically,the notable distinction between positive feedback and self-activation is the sign of the net synergy∆I(s;v,x),i.e.,∆I(s;v,x)>0 in Fig.3(b)but∆I(s;v,x)<0 in Fig.5(b),implying that synergy is prevalent over redundancy in the former case,but redundancy is remarkable in the latter.By making a comparison between Figs.3(f)and 5(f),it can be seen that for selfactivation,the mutual information I(s;v,x)and I(s;x)are almost independent of the change of degradation rateµx,but just slightly increase for very slow µx.However,for positive feedback,increasingµxcan cause a reduction in I(s;v,x)and a prominent peak appears in the curve of I(s;x).
Forµs> µv,according to Figs.5(d)and 5(h),the net synergy also exhibits an approximately threshold behavior,and the variation range of net synergy is smaller compared with the other three cases for self-activation.Moreover,the changes of the mutual information with increasingµxare analogous to the case of positive feedback,whereas the maximum value of net synergy∆I(s;v,x)is much smaller and I(s;v,x)is less than that in the case of positive feedback.
(iv)The Case of Self-Inhibition α =v and β =kv
For self-inhibition,the effects ofµxon the net synergy and mutual information in the information transmission are presented in Fig.6.Whenµs≤ µv,the changes of the net synergy and mutual information with the increasing ofµxare almost the same as those of negative feedback,corresponding to Figs.6(a),6(b),6(d),and 6(e).
Forµs> µv,the peak value of net synergy∆I(s;v,x)shown in Fig.6(c)is larger than that in Fig.4(c).Although the net synergy displays an approximately threshold behavior,the respective threshold values are different in the two cases.Meanwhile,the remarkable peak of I(s;v,x),which appears in Fig.6(f)differs from that in Fig.4(f).However,the changes of mutual information I(s;x)are nearly the same for self-inhibition and negative feedback.
For each of the four feedback types mentioned above,the most amount of mutual information I(s;x)can be obtained whenµs< µv.By comparison,the least amount of mutual information I(s;x)can be achieved whenµs> µv,which is in agreement with Ref.[35].This indicates that information transmission capacity in the signaling cascade is hindered by slow degradation rate of the intermediate component V in contrast to input signal S.It is important to note that the change of mutual information I(s;x)with the increasing ofµxis related to the correlation degree between the input signal S and output component X.
According to the above results,it is obvious that different feedback mechanisms in the signaling cascade can give rise to different fluctuation effects,net synergy and mutual information by changing the degradation rates of three components,thus easily generating diverse responses in a genetically identical population.This is the reason why certain types of feedback arises frequently in some conditions,which may be related to their noise behavior and information transmission abilities.In turn,these behaviors are determined by their topologies.Hence,the choice of feedback framework is a mechanism of efficiently modulating stochastic fluctuations and information transduction abilities via varying degradation rates of components in the signaling cascade,which is important for the cells to survive in changing environments.
3.3 Effects of Degradation Rates on SNR for Different Feedbacks
To clearly demonstrate the relationship between the net synergy and the SNR in the signaling cascade involving in various feedbacks,in this section,we take into account the dependence of the SNR on degradation rates of components in the signaling cascade with different feedbacks,as shown in Fig.7.
Whenµs< µv,the response of the SNR to the change ofµxis similar in each of the four figures,i.e.,the SNR of the system increases with the increasing ofµxfor slowµx,and then approaches to a constant for fastµx.Moreover,there is an opposite variation tendency between the net synergy and the SNR in the signaling cascade,which is in accordance with Ref.[36].Meanwhile,in this case,the large values of the SNR of the system are attained for each of the four feedbacks.The reason is that some information is corrupted or lost due to inevitable stochasticfluctuations during the information transmission process.If some important information is corrupted or lost,then the correspond substitution can be provided at the output component as a result of redundant information.Hence redundancy in information transduction can enhance the SNR of the signaling cascade.
When µs≪ µv,there is an optimal value ofµxat which the maximum value of the SNR can be obtained for positive feedback.Correspondingly,there is also an optimal value ofµxat which the mutual information I(s;x)can reach its maximum in such a case(See the red dashed line in Fig.3(f)).However,these results of positive feedback are invalid in the case of self-activation.In fact,cells need the signaling cascade that encompasses different feedbacks to carry out some certain functions.It suggests that each possible feedback strategy may have a highfitness under certain environments,and the tunability of information transduction in feedback regimes would be utilized by organisms in order to better adapt to changes in their environments.Thus,the diverse performance of the system might explain why certain network motifs,or their combinations,preferentially occur in biological systems and are retained in evolution.
When µs= µv,the trend of change in the SNR in the signaling cascade with the alteration ofµxis similar to the case ofµs< µvfor each of the four feedbacks.For positive feedback,there exists a point of intersection(whereµx=3.71)for the SNR curves betweenµs≪ µvandµs= µv.That is,the SNR of the system in the former case is higher than that in the latter before reaching the intersection point at a given value ofµxand becomes lower after the point of intersection.
When µs> µv,the low SNR of the signaling cascade is obtained for each of the four feedbacks in Fig.7.Moreover,the SNR of the system is not enhanced even though the Fano factor value of output component X is small(see Figs.2(b)and 2(d))for negative feedback and self-inhibition.This suggests that the slow degradation rate of the intermediate componentµvimpedes information transmission in the signaling cascade,and limits the amount of information transmission from the input component to the output component again.

Fig.7(Color online)Effects of degradation rates on the SNR for different feedbacks:(a)Positive feedback α=x and β =x.(b)Negative feedback α =x and β =kv.(c)Self-activation α =v and β =v.(d)Self-inhibition α=v and β=kv.
3.4 Comparisons the Results with Different Methods
Fig.8 (Color online)Fano factors and the net synergy in the caseµs< µvas a function of the degradation rate obtained by theoretical computation(lines)and by the Gillespie algorithm(symbols)with the same and different parameter Ω for four feedbacks:positive feedback(black solid lines with open circles),negative feedback(red solid lines with open upward triangles),self-activation(blue solid lines with open diamonds),and self-inhibition(dark cyan solid lines with open stars).
The aim of this paper is to study the fluctuation effects and information transmission in the signaling cascade with different feedbacks.In order to study the fluctuation effects and information transmission in biological systems,there are diverse standard methods as mentioned in the introduction.Thus in this section,we compare numerical results obtained by using LNA method[46,48,52]with those of the Gillespie algorithm(an accurate simulation algorithm).[47]
In Fig.8,for different feedback types,the Fano factor of X and net synergy,which change by varying the degradation rate of the output componentµxwith the same and different parameter Ω,are compared for both approaches,respectively.It is apparent that the results of these two methods are in accordant with each other in the caseµs< µvfor different feedbacks.Hence our theoretical results obtained by the LNA method are valid for our system studied here.These results suggest that the signaling cascade involving in diverse feedback mechanisms can regulate a variety of cellular processes,and exhibit very rich kinetic characteristics.
4 Conclusions
In this paper,a signaling cascade with different feedback mechanisms has been investigated,the theoretical formulas of the Fano factor,the net synergy,and the SNR of the system are derived by using the linear noise approximation,and the effects of the degradation rates of components on the fluctuation and information transmission in the signaling cascade with four feedback mechanisms are analyzed and discussed.
In the case ofµs< µv,the net synergy∆I(s;v,x)<0 for each of the four feedback types,but for positive feedback and self-activation,the absolute value of negative net synergy is larger and the mutual information I(s;x)is more than that of negative feedback and self-inhibition at a given value ofµx.In this case,high SNR of the system is obtained in information transmission.This suggests that for the four feedbacks considered in this paper,when µs< µv,redundant feature of the target variables is prevalent over synergistic property in the information transfer process and redundant information can compensate for either damaged or lost information,thus improve the SNR of the signaling cascade.
In the case ofµs≪ µv,for positive feedback,the minimum value of Fano factor of X is obtained in the signaling cascade.Particularly,there exists an optimal value ofµxat which the maximum value of the SNR can be achieved in this case.However,the SNR of the system is not higher than that in the caseµs< µv.In contrast,there is no optimal value ofµxfor self-activation.
In the case ofµs= µv,for positive feedback and self-activation,the net synergy exhibits an approximately threshold behavior with the change ofµx.However,for negative feedback and self-inhibition,the threshold behavior does not occur,and the net synergy is a monotone decreasing function with the variation ofµxin both cases.
In the case ofµs> µv,for positive feedback and selfactivation,significant relative fluctuations in output component X are obtained,and redundant effect of the target variables is dominant in the information transduction for fastµx.By comparison,for negative feedback and self-inhibition,the minimum value of Fano factor of X is achieved,and redundancy is prior to synergy in the information transduction except for very slowµx.
Above results are in accord with some closely related phenomena in biological systems,such as the osmo adaptation,[58]the bacterial chemotaxis,[43]and the responses of S.Cerevisiae to the changes in external carbon source.[59]Hence different feedback mechanisms exhibit various properties in the fluctuation effects and information transmission by changing the degradation rates of the components in the signaling cascade.
The signaling cascade with different feedback mechanisms studied in this paper is a typical regulatory motif in various network systems,therefore,it may be worth to investigate the impacts of time delays of production rates and degradation rates of components in some biological signal systems(e.g.,the intracellular signal pathways,[60−63]the neural signal systems,[64−69]etc.)the synergy and redundancy in information transmission in our future works.Our results not only give an extension to the previous investigations of information transmission in the signaling cascade,but also provide a good measure for understanding how biochemical networks transduce time-varying input signals and have potential applications in various fields such as climate science,financial industry,and computer networks.
猜你喜欢
杂志排行

Communications in Theoretical Physics的其它文章
- P-V Criticality of Born-Infeld AdS Black Holes Surrounded by Quintessence∗
- Baryogenesis in f(R,T)Gravity∗
- Prospect for Cosmological Parameter Estimation Using Future Hubble Parameter Measurements∗
- Topological Dark Matter from the Theory of Composite Electroweak Symmetry Breaking∗
- Pair Production in Chromoelectric Field with Back Reaction∗
- Impact of Internal Heat Source on Mixed Convective Transverse Transport of Viscoplastic Material under Viscosity Variation