我国PEDV毒株结构蛋白遗传变异情况分析
2018-11-22曾旭健包银莉
曾旭健,包银莉
(1.成都农业科技职业学院,四川 成都,610030; 2.四川海林格生物制药有限公司,四川 成都,610030)
猪流行性腹泻是由猪流行性腹泻病 毒(Porcine Epidemic Diarrhea Virus, PEDV)引起的一种猪急性、感染性肠道疾病,临床上以患病仔猪呕吐、水样腹泻、脱水、消瘦为典型症状。在我国,2010年前主要以散发为主。2010年后该病全面暴发,并呈现新的流行特征:患病仔猪出现更加严重的腹泻、呕吐和脱水症状,死亡率高达80%~100%。这些特征提示现行流行毒株与现有疫苗毒株可能存在较大的差异。文章从PEDV结构、流行株结构蛋白遗传变异情况、我国疫苗株基因分群情况以及流行株对疫病防控作用等多个方面对我国PEDV毒株结构蛋白遗传变异情况进行分析,以期为猪流行性腹泻的合理防控提供理论依据。
1 PEDV的结构
PEDV属于α-冠状病毒,呈现典型的冠状病毒结构。基因组全长约为28kb,由至少7个开放阅读框(ORFs)构成,分别编码4个结构蛋白(S蛋白、E蛋白、M蛋白、N蛋白)和3个非结构蛋白(ORF1a,ORF1b,ORF3)。
2 流行株结构蛋白遗传变异情况
PEDV作为一种基因组较大的RNA病毒,容易发生核酸变异,导致新的变异株甚至基因型的出现。在PEDV的主要结构蛋白基因中,S基因的变异最大。
2.1 S蛋白
S蛋白是位于病毒粒子表面的纤突糖蛋白,由1383个氨基酸(aa)组成。根据 PEDV 与其他冠状病毒S蛋白的同源性,可将S蛋白分为S1(1-789 aa)和S2(790-1383 aa)两个部分。其中,S1是PEDV基因组的高变区域,包含多个中和表位和受体结合域,与病毒抗原性和吸附入侵密切相关。S2区变异性小,在病毒与宿主细胞膜融合的信号转导过程中发挥重要作用。
研究指出,目前不同PEDV毒株基因组的差异主要集中在S基因,特别是S1区域。在不同的 PEDV 毒株中,氨基酸的突变、插入、缺失现象均较为常见。通过对我国2014-2017年间各地报道的PEDV流行株的文献进行梳理,发现近年来PEDV流行毒株S基因遗传变异主要有以下特点:1)由表1可见,各地区分离的PEDV流行株在基因水平上与传统的PEDV的CV777株同源性较低,均不高于95%,说明近年来PEDV毒株确实在基因水平上发生了变异;2)变异情况往往涵盖基因缺失、插入和基因突变多种情况;3)突变区域主要分布在S1区;4)部分变异发生在PEDV毒株抗原保护位点,从而导致PEDV流行株免疫原性的改变;大部分变异为随机发生,无规律可循,变异后所导致的毒株变化不得而知或处于研究状态中。
2.2 M 蛋白
M蛋白是PEDV的囊膜糖蛋白,由226个氨基酸组成。编码M蛋白的基因核苷酸序列具有高度的保守性。然而也有部分文献指出,M基因也出现了一定的进化和变异的趋势。这种趋势主要表现为核苷酸序列上的基因突变。董波等对2015年1月-2017年6月期间采自闽西地区规模化养猪场12份样品分析发现,闽西毒株M基因与CV777 同源性为98. 7% 。此外,董波等指出在M基因上虽有11个基因位点发生突变,但是反应在氨基酸水平上,其仅表现出3个氨基酸的改变。这些数据表明,不同PEDV毒株的M基因不论是在基因水平还是在氨基酸水平,仍然处于相对保守阶段。2018年,丁卫星等对收集的2株PEDV临床毒株(D1和D3)的M基因进行克隆,分析后发现,与CV777相比,D1株存在12个核苷酸的插入,导致存在4个氨基酸的插入。这个发现与以往的基因突变形式是不一样的。基因插入是否会成为M基因变异的新的形式还有待进一步的研究。总体而言,现阶段M基因虽有突变,但仍是一个相对保守的基因。
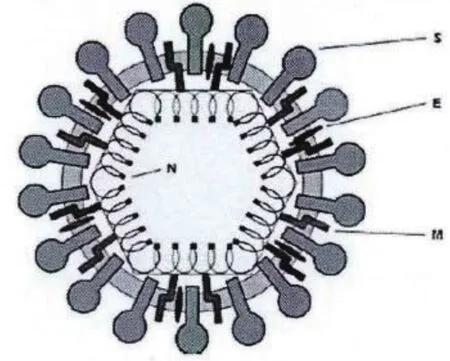
图1 PEDV结构示意图(毕静,2013)

图2 PEDV基因组结构示意图(王恩雨,2018)
2.3 E蛋白
E 蛋白是位于病毒囊膜上的小包膜蛋白,由76个氨基酸组成。研究发现E基因在同群冠状病毒中具有很高的同源性。

表1 近年来PEDV流行毒株S基因遗传变异情况汇总
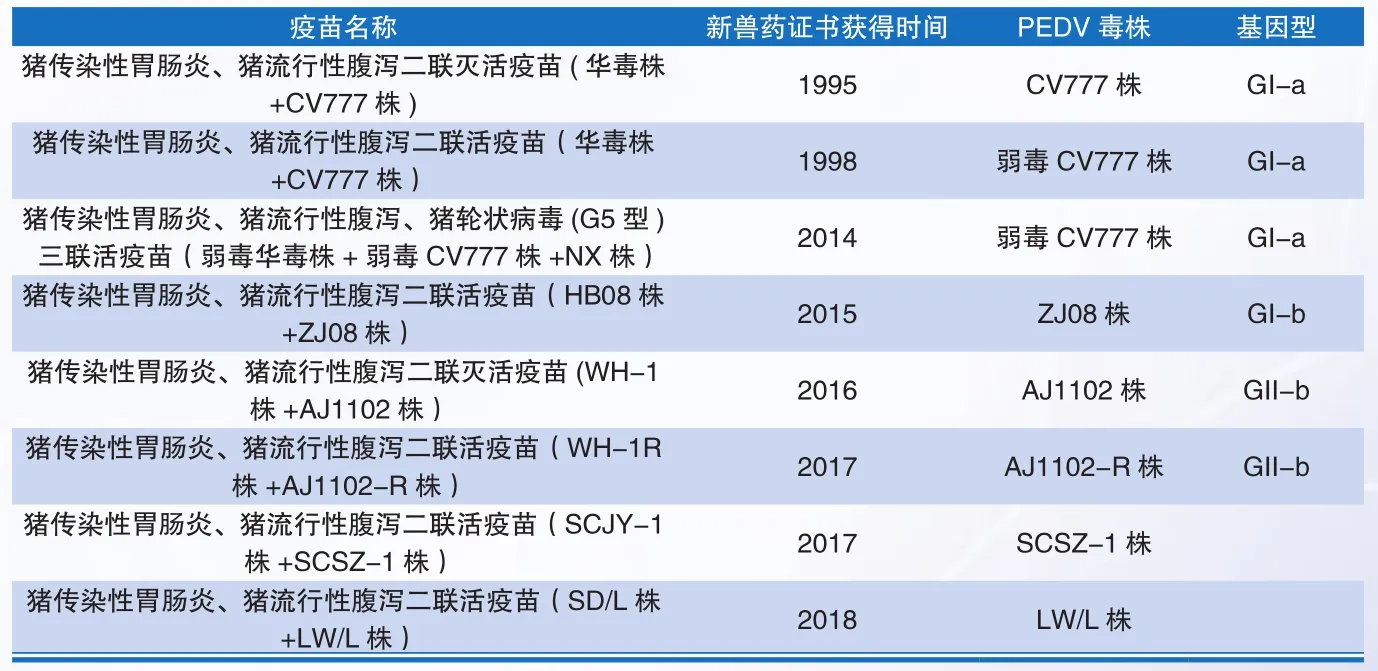
表2 我国PEDV疫苗株基因分群情况一览表
2.4 N蛋白
N蛋白是核衣壳蛋白,由441个氨基酸构成。Chen等对中国部分省的32个猪场当前流行的 PEDV的N基因进行测序分析,结果显示毒株之间同源性大于95.0%,和中国参考株同源性为95.6%~99.7 %,与致弱参考毒株CV777,DR13和83P-5 的核苷酸同源性为95.9 %~100.0% 。此外,所有的分析结果显示,目前PEDV毒株之间N基因的变异为核苷酸的突变,不涉及核苷酸插入和缺失。这些研究结果表明N蛋白还是非常保守的。
3 我国疫苗株基因分群情况
自首个猪传染性胃肠炎、猪流行性腹泻二联灭活疫苗上市以来,目前,我国共有8个PEDV联苗获得了新兽药证书,涉及PEDV疫苗毒株7个,分别为CV777株及其致弱毒株弱毒CV777株,AJ1102株及其致弱毒株AJ1102-R株,弱毒株ZJ08株,SCSZ-1株和LW/L株。
Wang Dong等于2016年对60株PEDV中国流行株和12株PEDV经典株进行了全基因组进化树分析,结果显示,这些菌株可以划分为两个基因群:GI和GII。其中它们又可进一步划分为GI-a、GI-b、GII-a和GII-b四个亚群。在这个分群体系中,经典株集中在GI群,流行株主要集中在GII群中。对PEDV疫苗株基因群进行分析,结果显示,CV777株及其致弱毒株弱毒CV777株作为经典株,属于GI-a亚群;ZJ08株属于GI-b亚群;AJ1102株及其致弱毒株AJ1102-R株则属于GII-b亚群。文中未对新近获得新兽药证书的疫苗株进行序列分析,相关文献显示,SCSZ-1株和LW/L株与经典株CV777同源性较低,与流行株同源性可达98%以上。
4 流行株对PEDV防控的作用
PEDV作为一个含有大基因组的RNA病毒,与DNA病毒相比,其病毒基因组更容易发生重组和变异。从遗传进化上而言,变异是病原微生物主动适应环境和宿主的一种表现。当其生存环境发生改变时,其必然进行自我改变以适应新的生存环境。当环境压力增大,病原微生物不论是在变异的程度上,还是在变异的速度上都会有显著地提升。这也就解释了我国猪流行性腹泻疫情以2010年为分水岭、后期呈现暴发形势的原因。
众所周知,对于疫病防控而言,选用与导致该疫病发生的病原微生物进行免疫接种往往会获得非常好的免疫保护作用。但是在现实情况下,完全依赖流行株对疫病进行防控是非常不可取也无法完全实现的。一方面,基因的变异是一种生物自发的行为,环境压力不同导致的变异情况也就不同,从而出现同一时间段不同地区不同猪场的流行株存在差别。一味的套用变异株死/活疫苗,只会让自己的猪场疫病情况更加复杂。另一方面,疫苗的研发是有滞后性的。目前虽然可以利用分子方法对流行株的基因组进行分析,但是无法对变异进行预测。而疫苗开发是需要时间的。一个新的疫苗从研发、生产到最终的上市需要大量的时间,而在这段时间中毒株的变异仍然在发生。因此人们永远用不上由最新的流行株生产的疫苗。再次,毒株是疫苗不可或缺的一部分,但不是唯一的构成。一种疫苗的好坏不仅与毒株有关,还与佐剂、生产工艺、疫苗质量等因素息息相关。而对于疫苗的临床使用效果而言,疫苗的使用方法、免疫程序、保存条件等因素更是至关重要的。
对于猪流行性腹泻的防控而言,我们不仅要选择适宜的疫苗,更要采用合适的免疫方式,结合最优的免疫程序,做好环境和饲养的管理以及其他生物安全防护方面的措施。
(参考文献略。如有需要,请与作者联系。)
