中国四川省峨眉山特有小鲵类(两栖纲,有尾目)龙洞山溪鲵(Batrachuperus londongensis Liu and Tian, 1978)正模标本骨骼μCT三维数据集
2018-11-17贾佳张美华高克勤江建平
贾佳 ,张美华,高克勤,江建平
1.北京大学地球与空间科学学院,北京 100871
2.中国科学院资源地层学与古地理学重点实验室,中国科学院南京地质古生物研究所,南京 210008
3.中国科学院成都生物研究所,成都 610041
数据库(集)基本信息简介
数据库(集)名称 中国四川省龙洞山溪鲵正模标本(CIB 14380)高精度μCT扫描源数据集数据作者 贾佳、张美华、高克勤、江建平数据通信作者 贾佳(jia_jia@pku.edu.cn)标本采集时间 龙洞山溪鲵的正模标本采集于1965年3月23日地理区域 龙洞山溪鲵的正模标本发现于中国四川省峨眉山龙洞河,地理坐标为北纬29°34′42.85″,东经103°17′5.61″,海拔1300米。

?
引 言
蝾螈类又称有尾两栖类,是一类四肢与尾巴正常发育的中小型滑体两栖动物。作为现存最为原始的四足动物,蝾螈类与其他滑体两栖动物(蛙类与蟾蜍、蚓螈类)相比在形态上与已经绝灭的原始两栖动物更为相似,因此对了解脊椎动物由水生环境向陆地环境的过渡、四足动物的早期演化和滑体两栖动物的起源具有重要的研究意义[1]。
小鲵科(Hynobiidae)是一类小型蝾螈(体长约70~260毫米)[2],包括9~11属,67~68个现存物种,主要分布在东亚地区,仅有个别属种分布在中亚(副趾鲵属Paradactylodon)和欧洲东部的俄罗斯境内(极北鲵属Salamandrella)[3-4]。该科成员的幼年个体生活在水中,具有外鳃和尾鳍;成年个体通常经过变态发育之后,外鳃和尾鳍通常消失,可生活在水中或陆地上。唯一的特例是龙洞山溪鲵:它们的某些个体不经过变态发育,却成为了保留幼体特征(如鳃裂)并终生生活在水中的童体类型[2]。小鲵科因其具有多个原始特征,如体外受精、较高的染色体数目(40个以上)、具有小染色体、脊神经由椎体之间穿过(第一条脊神经除外)、下颌中隅骨不与前关节骨愈合等,被广泛认为是现存蝾螈类中的一个原始类群[5-7],对蝾螈类的起源研究至关重要。
然而,前人对小鲵科的起源时间和地点[8-12]、某些特征的演化历史[13-14]和内部谱系分支关系[11,15-17]存在较大分歧。小鲵科曾一度被认为是一个并系类群(paraphyletic group[18])或复系类群(polyphyletic group[14]);而它的单系性,即包括同一共同祖先演化而来的所有后裔的演化分支,直到最近才得到分支系统学研究的支持(参见[11,15])。造成以上争议的原因可分为以下四点:第一,该科化石记录稀缺。目前仅发现5个新生代(6600万年至今)化石属种,而且它们多保存为零散的脊椎和肢骨,鲜有可靠的形态特征信息[12]。近年来,中国辽西、冀北和内蒙古东部地区中侏罗世到早白垩世(距今1.25~1.65亿年前)地层中产出了6个在形态特征上与现存小鲵类极为相似的化石属种,即东方塘螈(Laccotritonorientalis)、凤山中华螈(Sinerpetonfengshanensis)、钟健辽西螈(Liaoxitritonzhongjiani)、道虎沟辽西螈(Liaoxitritondaohugouensis)、围场皇家螈(Regalerpetonweichangensis)和北方诺敏螈(Nuominerpetonaquilonaris)[19-20]。这些化石保存完整,为探讨小鲵科的起源提供了珍贵材料,然而它们与现存小鲵类的分支关系并不明确(见下文),仅有钟健辽西螈和北方诺敏螈被认为是小鲵科的基干属种[20-22];第二,对小鲵科的现存属种,特别是对较为进步的中国特有小鲵类(如山溪鲵类)的骨骼形态学研究不足,加重了从充盈原始特征的小鲵类中寻找近裔共性(synapomorphy)的难度。第三,基于分子数据对该科的分支分析结果不一致。随着基因测序技术的突飞猛进,现存小鲵类的分子数据正以愈加低廉的成本快速地积聚。独立演化的基因位点为探讨小鲵类的分支关系提供了大量有效信息。但是前人根据线粒体DNA[11,23]、细胞核DNA[17]以及结合二者[15-16,24-25]所得出的分支分析结果依然存在差别,不能提供一致的小鲵科内部支系演化关系。第四,目前还没有基于形态特征数据对小鲵科进行分支分析的研究。前人曾对小鲵科具有演化和分类意义的形态特征有过总结[10,26-27],然而却并未严格地按照分支分析理论[28]进行研究,而且所得结果与基于分子数据得到的分支分析结果差异悬殊。
如上所述,尽管分子数据以其骤增的体量占较大优势,现生小鲵科成员的形态特征依然是研究小鲵类支系演化相关问题不可或缺的组成部分。另一方面,大部分小鲵类化石仅保存了骨骼信息,只有少数情况下较好地保存了软体信息。因此形态特征,尤其是骨骼形态特征,是唯一能够链接化石与现存属种的桥梁,对探讨化石属种与现存小鲵类支系演化关系至关重要。此前,对现存小鲵类的形态特征研究主要依靠双染色法(参见[29-31])。该方法通过使用染色试剂将硬骨和软骨分别染成红色和蓝色,因而能够很好地呈现出骨骼(软骨和硬骨)的轮廓[32],不足之处在于无法获取标本内部的三维结构信息,而且对标本造成了不可逆的损坏。高精度计算机断层扫描(high-resolution X-ray tomography, 或μCT扫描)是利用高能X光射线扫描标本并在短时间内获取标本三维结构的技术。该技术可以完好无损地获取重要标本的骨骼虚拟信息,方便研究人员的观察。
针对上述问题,本文选取栖息于中国四川省峨眉山龙洞河地区的龙洞山溪鲵(Batrachuperuslondongensis)为研究对象,对其正模标本(CIB 14380)进行了μCT扫描。这是μCT扫描技术在中国现存小鲵类骨骼研究中的首次应用,也是我们利用μCT扫描技术实现对现存小鲵类骨骼系统研究的一系列结果之一[33]。如前所述,龙洞山溪鲵同时具有变态发育和兼性童体型现象,在发育方式上有别于其他所有现存小鲵类。然而,该属种从被正式确立[34]至今已有40年,此前却并未有针对其骨骼系统的详细研究。μCT扫描揭示出龙洞山溪鲵在头骨、舌器和肢骨上存在多个特殊的骨骼形态特征(见2数据样本描述)。
本数据集提供了龙洞山溪鲵正模标本(CIB 14380)的μCT扫描原始数据、三维重建图片和视频、头部的3D pdf文件和可用于3D打印的stl文件。以上原始数据的共享旨在为相关科研工作者提供可用于对比研究的龙洞山溪鲵的原始资料,同时也为公众了解龙洞山溪鲵的骨骼信息提供了渠道。
1 数据采集和处理方法
1.1 标本与数据采集
1.1.1 标本采集
龙洞山溪鲵的正模标本于1965年3月23日由刘承钊等人采集。采集地点位于四川省乐山市峨眉山零公里附近龙洞河(N29°34′42.85″、E103°17′5.61″),海拔 1300 米。此后该标本浸泡在福尔马林溶液中,并保存在成都生物研究所两栖爬行动物标本馆模式标本室,野外编号为CIB 65I0013,正式馆藏编号为CIB 14380。该标本为一雄性成年个体,全长265毫米,吻臀距(吻端至肛孔后缘的长度)129毫米,头长(吻端至枕髁后缘之间的长度)27.5毫米,头宽(头部两侧的最大距离)23.8毫米。头部后外侧保留鳃裂,属于童体型个体。我们将该标本从福尔马林溶液中取出,使用佳能(Canon EOS 5D)单反相机对其进行照相(见图 1)。
1.1.2 μCT数据采集
龙洞山溪鲵正模标本的 μCT扫描工作依托成都生物研究所的扫描仪 Quantum GX microCT Imaging System(PerkinElmer®,Massachusetts,USA)进行。
把拍照之后的正模标本移入密封袋,小心将其尽量缕直并卷成长条状。将标本固定在扫描仪的小动物床(Small Bed)上,并保持标本的长轴与动物床的长轴平行。设置电压为50千伏,电流为60毫安,通过调整扫描视域(Field of View)下的采集(Acquisition)和重建(Reconstruction)两个参数,使标本尽可能充满扫描区域。在预览(Live)模式下,通过调整 X光射线源的旋转角度(Rotation Control)来确保所扫描的结构在任意角度均处于扫描范围之内。之后,选择高精度扫描模式(High Resolution)下的57分钟档位对标本的每一段进行扫描。由于龙洞山溪鲵的正模标本较长,为了获取较高的分辨率,需要分段对其进行扫描(图 1):通过调整动物床的位置将标本沿其长轴进行水平移动,待移动到下一段后,重复上述设置对其进行高精度扫描。移动过程中,确保相邻两段扫描存在一定重叠区域,以便后续的图像拼接。
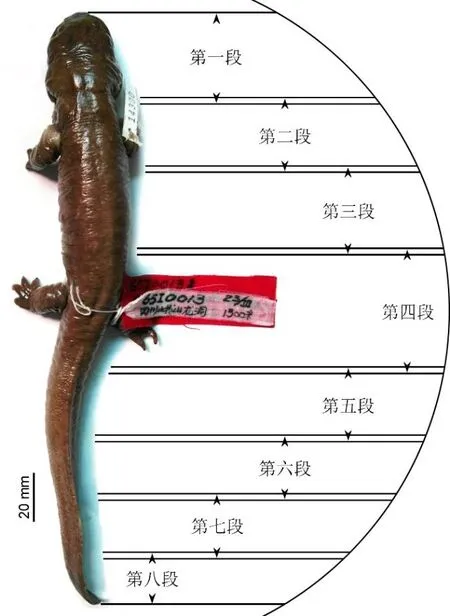
图1 龙洞山溪鲵正模标本CIB 14380照片及μCT分段扫描设计方案
结束每段扫描后,扫描仪生成10.3 GB的原始数据,包括803张raw格式的文件。通过扫描仪配套软件(Quantum GX microCT software)将每一段扫描之后的原始数据导出成横切标本长轴的512张16位tiff格式图片,总大小为256 MB,即本数据集第一部分数据的以下8个文件:skull90um.zip,forelimb120um.zip,trunk120um.zip,hindlimb144um.zip,tail_1_120um.zip,tail_2_120um.zip,tail_3_120um.zip 和 tail_4_120um.zip。
值得注意的是,该扫描仪(Quantum GX microCT Imaging System)与其他常用扫描仪(如GE Phoenix v|tome|x s Industrial High-Resolution CT & X-Ray System或Nikon XT H系列等)相比具有两处不同:首先,该扫描仪在运行过程中标本保持静止,而X光射线源和X光感受器围绕标本旋转360度;这与其他扫描仪的扫描过程恰好相反。其次,该扫描仪具有分体积重建功能(Subvolume Reconstruction),可以在原始扫描数据的基础之上通过人为选择感兴趣区(Region of Interest),并将所选区域进行插值法运算,从而提高该区域的分辨率(Voxel Size)。本文对龙洞山溪鲵正模标本的头骨,即第一段的原始扫描数据(文件skull90um.zip)进行了分体积重建,其分辨率由原来的90 μm提升至67.1 μm,即本数据集第一部分数据的第9个文件,skull67_1um.zip。
1.2 数据预处理
对正模标本进行的图像拼接(Merge)、图像分割(Segmentation)、三维重建(3D Reconstruction)、图像和视频生成和 stl文件导出均利用软件 VG Studio Max 2.2(Volume Graphics, Heidelberg, Germany)进行。
第一步图像拼接。首先将每一段扫描生成的512张tiff格式图像堆栈分别导入软件VG Studio Max 2.2中,并分别保存为单独的vgl格式文件。打开VG Studio Max软件的一个空白文件,使用该软件的拼接功能(Merge)将以上vgl格式文件依次导入。按照龙洞山溪鲵的解剖结构,并结合扫描时相邻两段之间的重叠区域将所有vgl文件拼接起来,最终得到完整的骨骼虚拟模型。
第二步图像分割和三维重建。以龙洞山溪鲵的头骨为例,在VG Studio Max 软件的二维窗口中,将头骨的每一块骨骼单独建立感兴趣区,并将每一块骨骼在所有截面中的轮廓画入相应的感兴趣区。对感兴趣区进行平滑(Smooth)或修正(Refine)处理,将感兴趣区提取(Extract ROI)、渲染(Render)便得到相应头骨骨骼的三维结构模型(图2)。
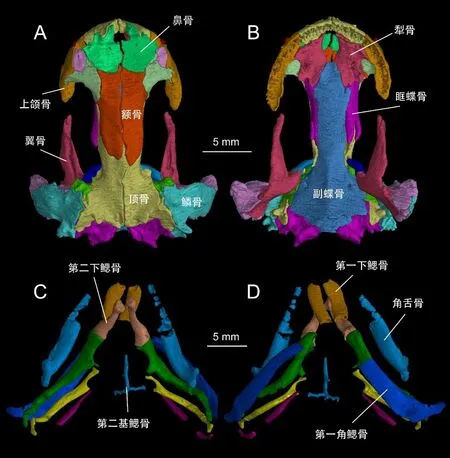
图2 龙洞山溪鲵正模标本CIB 14380头部骨骼三维重建图
第三步图像、视频和stl文件制作。仍以头骨为例,在VG Studio Max软件的三维窗口中,将渲染后的头骨放大并旋转到相应角度(如顶视、腹视、侧视等),选择文件选项卡并保存图像(文件pics.zip)。同样地,在三维窗口中,通过软件自带的关键帧(Keyframer mode)功能,将头骨保存为沿某一轴向旋转的视频文件(文件videos.zip)。将每一个提取出来的文件进行表面测定(Surface Determination),进而导出stl文件(pdf_stl.zip)。
2 数据样本描述
2.1 数据集文件命名格式
龙洞山溪鲵正模标本(CIB 14380)高精度μCT扫描源数据集的命名规则如下:
(1)该数据集第一部分包括 9个文件。以第一个文件 skull90um.zip和第五个文件tail_1_120um为例,它们的命名规则如下:skull表明该文件包括了对头部的高精度扫描,90um代表该段的扫描分辨率为 90 μm。tail_1表明该文件包括了对尾部第一段的高精度扫描,120um代表该段的扫描分辨率为120 μm。每个文件夹下包括一个xml参数文本文件和512个tiff格式文件。所有tiff格式文件均分别添加了连续的后缀p001-p512。读者利用三维软件(如VG Studio Max等)导入以上tiff文件时,务必按顺序连续读取,以确保骨骼结构的完整性。
(2)该数据集第二部分包括三个文件,分别为pics.zip,videos.zip和pdf_stl.zip,包括了三维重建的图像、视频以及3D pdf和stl文件。读者可用任意版本的Adobe Reader软件打开3D pdf,激活内容并查看龙洞山溪鲵的头部骨骼。如果需要制成实体模型,可将stl文件进一步处理,并利用3D打印机按照同一比例打印即可。
2.2 数据样本
龙洞山溪鲵的系统分类信息如下:
两栖纲 Amphibia Linnaeus, 1758
滑体两栖亚纲 Lissamphibia Haeckel, 1866
有尾超目 Caudata Scopoli, 1777
有尾目 Urodela Duméril, 1806
隐鳃鲵亚目 Cryptobranchoidea Dunn, 1922
小鲵科 Hynobiidae, 1859
龙洞山溪鲵Batrachuperus londongensisLiu and Tian, 1978
龙洞山溪鲵是小鲵科中较为独特的一种,因其既有完全变态发育的成体也有保留鳃裂等幼体特征的童体类型。正模标本(CIB 14380)具有鳃裂,属于童体类型。μCT扫描揭示出正模标本的舌器骨化程度较高,包括成对的第一下鳃骨(hypobranchial I)、第二下鳃骨(hypobranchial II)、第一到第四角鳃骨(ceratobranchial I–IV)、角舌骨(ceratohyal)和第二基鳃骨(basibranchial II)(图2)。与龙洞山溪鲵的变态发育个体和其他小鲵类中通常仅有第二下鳃骨和第二角鳃骨骨化的情况相比,骨化程度显然更高[33-34]。
图3展示了正模标本的整体骨骼结构。可以看出,龙洞山溪鲵正模标本具有17枚荐前椎,而且前肢的腕骨(carpal,红色)和后肢的跗骨(tarsal,蓝色)骨化较高,与其他童体型蝾螈类(如鳗螈、两栖螈科等)腕、跗骨通常为软骨的情况形成鲜明对比。
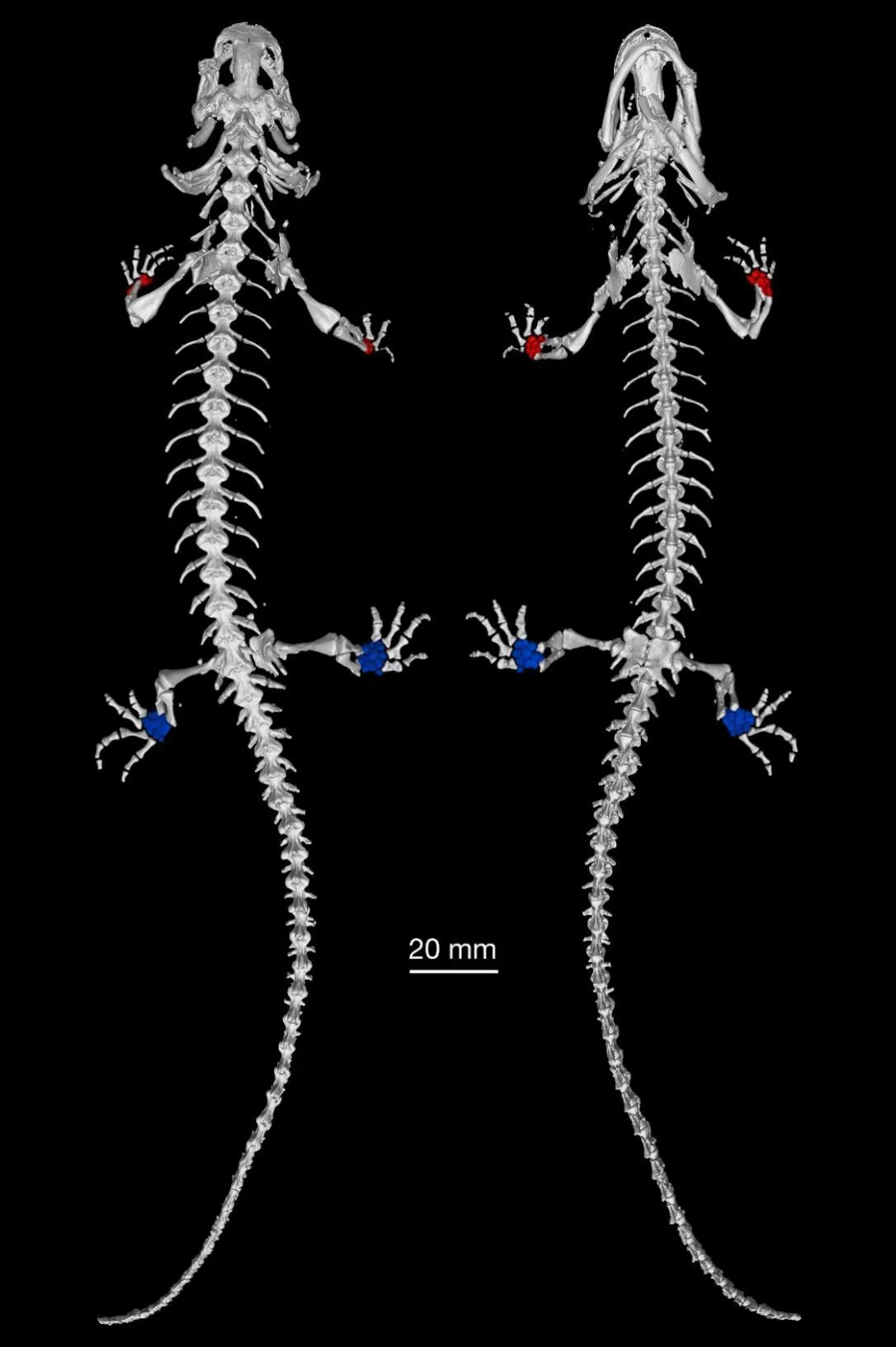
图3 龙洞山溪鲵正模标本CIB 14380整体骨骼三维重建图
3 数据质量控制和评估
本数据集包括利用高精度μCT扫描获取龙洞山溪鲵正模标本的所有原始文件和参数。读者可利用常用的三维软件(Amira、Dragonfly、ImageJ、Mimics、VG Studio等)按照参数文本中的分辨率读取。
μCT扫描所得的骨骼分辨率受以下两方面因素的影响:第一,扫描仪 Quantum GX microCT Imaging System所得到的分辨率取决于扫描视域(Field of View)的大小,而后者受限于采集(Acquisition)和重建(Reconstruction)两个参数的设定(表1)。其中,采集有36和72两个数值选项(无单位),相应的重建有36、25、10、5和72、60、45几个固定值(无单位)。
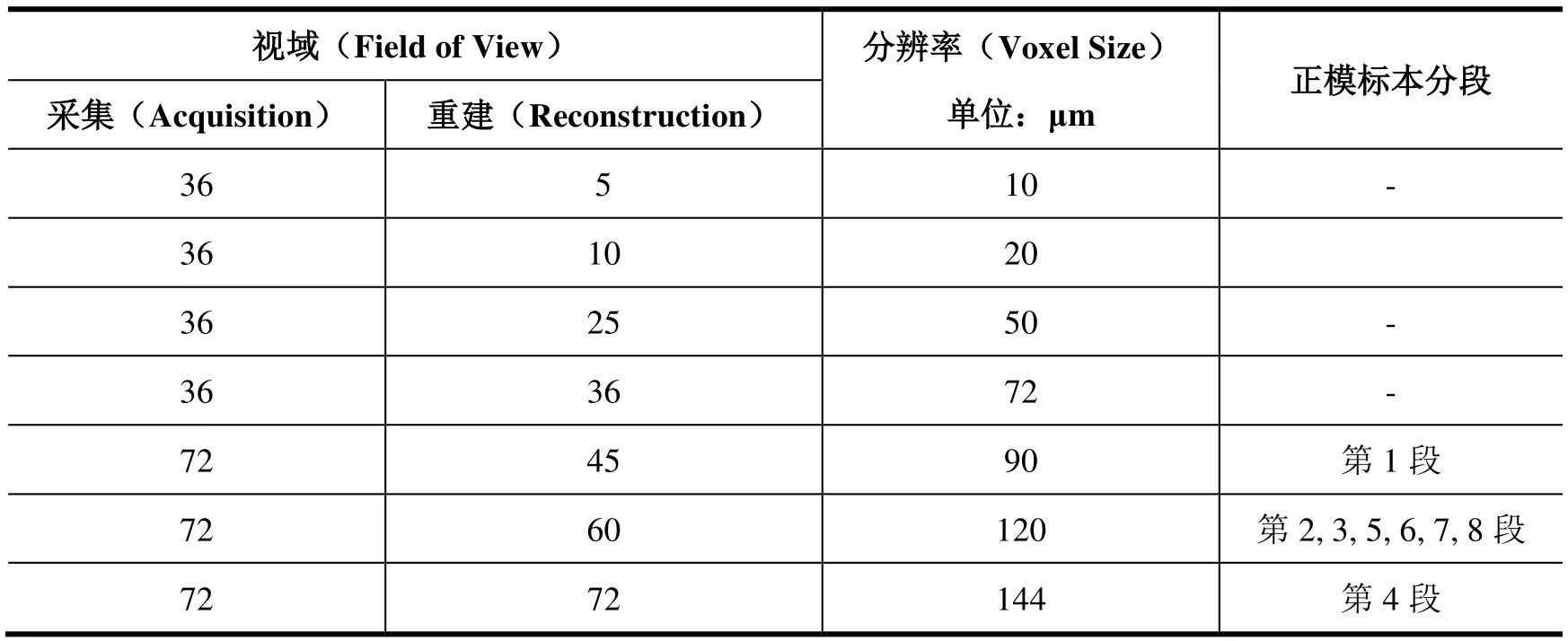
表1 Quantum GX高精度扫描系统小动物床的视域与分辨率大小对照表
第二,正模标本的不同部位由于宽度不同导致分辨率不同。该标本浸泡于福尔马林溶液中已有50余年,其肌肉和软体组织僵化,使得标本早已形成固定姿势。后肢部分由于向两侧伸展,只能在最大视域范围内,即采集和重建参数均为72的情况下进行扫描,因此所得分辨率(144 μm)最低。尽管如此,该标本每一段的骨骼结构在任意扫描截面上的轮廓均清晰可辨(图4)。
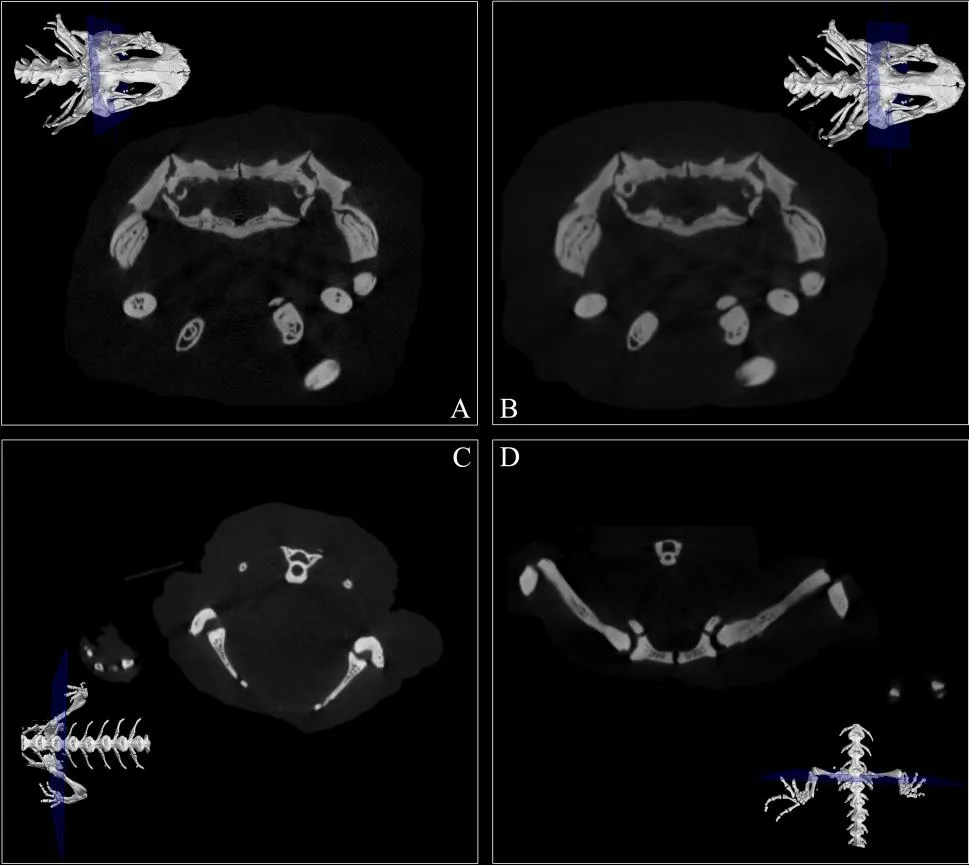
图4 龙洞山溪鲵正模标本CIB 14380分段扫描截面示意图
4 数据价值
本数据集共享了中国四川省峨眉山地区特有小鲵类蝾螈龙洞山溪鲵的正模标本(CIB 14380)高精度μCT骨骼数据,这是目前μCT扫描技术在我国现存小鲵科蝾螈类骨骼研究中的首次应用。本数据集所包含的数据便于处理,为相关科研人员提供了可供对比参照的原始数据。我们注意到国外类似共享扫描数据集的网站如 DigiMorph等已提供了蝾螈类部分属种的扫描数据,但是缺少中国特有属种的信息。龙洞山溪鲵骨骼数据的共享为我国建立相关的数据网站提供了基础数据材料,对我国小鲵科蝾螈类的形态演化研究具有重要价值。
