五台山林线过渡带华北落叶松格局过程和尺度分析
2018-11-15贾美玉张金屯丁沫冲贾冰玉
贾美玉,张金屯,*,梁 钰,丁沫冲,曹 格,贾冰玉
1 生物多样性与生态工程教育部重点实验室,北京师范大学生命科学学院, 北京 100875 2 山东农业大学林学院,泰安 271000
高山林线是高山带一个非常独特的研究实体。高山林线的乔木处于高寒、严酷气候胁迫的临界状态,对于全球性和区域性气候变化的反应敏感,它对全球气候变化具有预警作用[1- 2]。高山林线是高山带一条重要的生态界线,是郁闭森林与高山植被之间的生态过渡带[2]。林线过渡带植被对气候的响应按时间长短分为短期、中期、长期。一年左右短期的响应反应在个体特征上,几年到几十年的中期响应反应在生理响应上如年轮、高度和存活率等,百年以上的长期响应反应在多次演替的群落特征上[3]。林线研究的空间尺度也很丰富,多为局域尺度林线的研究[3- 4],也有跨区域尺度的研究[5-6]。对林线过渡带的研究有研究林线树木种群生长变化的[7- 8],有研究林线位置与气候响应变化的[9],但在外貌、格局和动态的研究相对较少,增加以上研究内容可以探究林线群落除环境因素作用以外的主导生态过程,有研究证明纬度和海拔高度对林线树木的限制大于人为干扰对林线的限制[3],2300m以上有树木种群建立但它的种子密度和生长率也会很低[10]。但也有实验结果表明草食动物对林线生态系统的限制作用也不容忽视[11]。对生长在2300m海拔以上并受放牧干扰的人工林生存变得异常艰难,它的种群年龄特征和格局过程的尺度研究也显得尤为重要,研究结果不仅可以捕捉种群发展的特征还可以推断类似地区生物和非生物因素对乔木的作用尺度为后期管理提供参考。
五台山属于温带落叶阔叶林区,是典型高山林线山地,林线附近的乔木为人工种植的华北落叶松[12]。五台山亚高山草甸是华北地区最典型,类型最丰富,草质和生产力最高的山地草甸,对亚高山草甸的研究包括合理利用、生态评价[13-14]以及群落生态学数量分析[15]。如五台山高山草甸群落分析表明地形因素影响群落型及优势种的划分,同时多样性也是影响划分的关键因素[16],五台山草甸中小格局分析揭示格局尺度对草本群落的生态意义[17]。作为著名风景名胜区,干扰的研究集中在旅游和放牧两方面,DCA方法分析了旅游干扰对五台山草甸的影响[18]。放牧对高山、亚高山草甸植被-土壤系统的影响[19],对牧草品质的影响表明短期的中度放牧对牧草品质有利[20-21]。五台山人工华北落叶松林是重要的景观组成,林线的华北落叶松存活状况堪忧,而它的存活率对于草甸的生产力和多样性影响很大,它的格局对于林下草本入侵也至关重要。已有研究利用聚类和排序分析了五台山高山林线的植被景观,得出了阴坡和阳坡的林线的海拔范围和草本植被群落类型分布的海拔范围[22],野外仪器测定分析五台山林线乔木的光合作用对生境差异的响应得出光照和温度相比水分是光合作用更为重要的限制因子[23]。本文在前面研究的基础上,希望通过点格局方法进一步分析华北落叶松的种群特点以及存活影响因素。点格局分析方法可以研究多尺度内森林种群的分布、生态过程对其格局的影响以及种群存活和死亡的动态[24],因此是有效的格局分析方法。
本实验选择五台山南台母树林基地树龄30以上的林线过渡带为研究样地,对样地内的华北落叶松种群进行分析。通过分析希望探究①林线过渡带华北落叶松种群生长特征以及死亡率与胸径分布的关系;②不同尺度内华北落叶松活立木和枯立木分布格局,以及空间异质性和种群密度对华北落叶松枯立木和活立木格局分析,进而推测其死亡的主要原因。
1 研究区概况
五台山位于山西省东北部,38°27′—39°15′N,112°48′—113°50′E。五台山位于蒙古高压中心的东南边缘地带。西伯利亚干冷气流吹向海洋的过程中,以强度波动性经过五台山向东南推移。海拔2250m以上山地的年均温约为-1.6—-4.9℃,年降水量717—840mm[25]。五台山所处地区属暖温带半湿润季风气候。五台山地区地形高耸,又临近华北平原,是接受东南海洋夏季风送来的暖湿气团较早和最佳的场所,五台山山顶年降水量高达900mm以上。五台豆村日照时数年平均2684.2h,日照充足[26]。由于地形变化较大,气候条件差异明显。植物资源丰富,拥有华北地区面积最大的亚高山草甸[27]。调查样地在2300m左右,属于山地草甸土。
2 材料与方法
2.1 野外取样
2017年7月初在五台山南台母树林基地以上选择有显著林线森林分布的高坡(2315m),坡向北偏西25°,坡度26°。从有显著林草交错带界限为起始的一边,往下坡方向延伸设置50m×50m的样地,用1m为单位刻度的100m长的样绳把样地分成5条样带,每条样带5个样方,共25个10m×10m的样方,对25个样方进行编号,在每个10m×10m样方中对乔木个体的位置和胸径信息进行调查,选定原点,在样地内记录每一棵华北落叶松(死+活)的位置和胸径信息。在50m×50m的样地内随机设置5个10m×10m的样方进行群落物种组成信息调查,统计出现的乔、灌、草物种。
2.2 数据处理与分析
点格局分析和零模型模拟的方法可以用来探索种群格局、种内两种不同属性个体间关系、种间关系以及生态过程在不同尺度上的效果。本研究选取成对相关函数g(r) 值为纵坐标,个体间距离为尺度作为横坐标进行绘图。首先运用完全空间随机CSR(Complete spatial randomness)模型对样地内华北落叶松的活立木和枯立木分别进行多尺度上的格局判断,此处结果可以比较多个尺度上活立木和枯立木分布特征;再者异质性泊松过程HP(Heterogeneous Poisson process)模型模拟样地内的空间异质性对格局的影响,通过异质性模型模拟结果的检验可以了解空间异质性对华北落叶松格局的作用是否显著;最后运用随机标签RL(Random Labeling)模型对活立木、枯立木个体进行尺度上的相关性分析,我们可以得到它们的相关关系以及进一步推测华北落叶松死亡原因。Monte-Carlo 方法用于检验实际值是否显著偏离零模型,一个点进行199个次模拟,在199次模拟中计算第五个最低值和最高值来获得α=0.05 的显著水平的值。在CSR模型进行格局判断结果中,当实际值超出置信区间的上包迹线为聚集分布,超出下包迹线为均匀分布。GoF test(Goodness of fit test)可以用于检验模型描述实际格局值的能力。比较选定范围内观察值和理论值的偏差,用秩检验判断显著性,当观察值的秩(rank)大于190说明观察显著偏离理论值。说明在α=0.05 水平上实际值显著偏离模型。模型不能很好解释形成格局的原因[28]。
K函数和g函数公式如下[29]:
式中,A为样方面积,n为研究个体的总点数,r为两点之间的距离即尺度,uij是i和j之间的距离,当uij
g(r)=dK(r)/dr×2πr
式中,g(r)>1,聚集分布;g(r)<1,均匀分布;g(r)=1,随机分布。
以下为3个零假设模型的介绍[28,30]:
a.完全空间随机CSR(Complete spatial randomness)模型
完全空间随机模型是最简单、常用的零模型,它假设个体之间不存在相互作用。它也叫同质性泊松分布,即研究区内所研究个体的密度是恒定的,且所研究个体在研究区域内的分布符合泊松分布,研究区域内每个点出现研究个体的概率是相同的。完全空间随机模型可以描述种群的真实格局,但对格局形成的生态过程缺乏解释。
b.异质性泊松过程HP(Heterogeneous Poisson process)模型
空间异质性常存在于研究区内,这时我们需要运用异质性泊松过程考虑样方内空间一次特征的零模型假设,进而模拟植物空间分布的二次特征。在异质性泊松过程中,所研究个体在研究区域内的密度是不恒定的,而随位置变化而变化,由λ(x,y)表示密度,其是运用圆形动窗在样方内进行插值运算而估计出的值,从而代表空间的一次特征,计算公式为:
式中,C(x,y)(R)是以(x,y)为圆心,R为半径的圆形动窗,Point[X]表示区域X内的点数,Area[X]表示区域X的面积。在这个模型中R需要谨慎考虑,R过大结果会接近CSR模型,R过小接近真实的分布,失去了空间一次特征假设的意义。经过10、5、3m的尝试后决定R=3m最为合适。此模型用于分析华北落叶松死亡的原因是不是与生境相关。因此采用华北落叶活立木做为背景,零模型假定活立木与枯立木的生境没有显著性差异。
c.随机标签(Random labeling)模型
随机标签模型属于双变量假设模型的一种,它可以用来分析两个物种或者同一物种两类型的格局在研究尺度上的相关关系。它假设两个变量的空间分布格局由相同的随机过程(或受到相同的限制条件)产生的。每一次循环模拟中,给变量1的点随机打上标签,计算其周围研究尺度内出现变量1或变量2的个体的概率。此模型用于分析活立木与枯立木的相关关系以及枯立木形成原因。
运用Programita 软件进行点格局分析,分析结果在Origin 9.1软件中对g(r) 、模拟值和置信区间在0—25m上进行作图。
3 结果与分析
3.1 华北落叶松群落结构与物种组成
调查50m×50m样地内,共433棵华北落叶松,其中活立木366棵,枯立木67棵。根据计算可知平均密度为5.77m2/棵,所有活树的平均高度为4.96m,枝下高为2.84m。其中郁闭度接近80%,群落中木本植物物种丰富度不高但草本植物种类多样,灌木有华北覆盆子(Rubusidaeus)、瘤糖茶藨子(Ribeshimalense),草本植物有多叶早熟禾(Poaplurifolia)、胭脂花(Primulamaximowiczii)、紫苞风毛菊(Saussureapurpurascens)、瞿麦(Dianthussuperbus)、异燕麦(Helictotrichonschellianum)、黑柴胡(Bupleurumsmithii)、小红菊(Dendranthemachanetii)、疏齿银莲花(Anemoneobtusiloba)、高原毛茛(Ranunculustanguticus)、早春苔草(Carexsubpediformis)、高山紫菀(Asteralpinus)、三叶委陵菜(Potentillafreyniana)、雪白委陵菜(Potentillanivea)、互叶金腰子(Chrysospleniumalternifolium)、峨参(Anthriscussylvestris)、紫花碎米荠(Cardaminetangutorum)、珠芽蓼(Polygonumviviparum)、北京假报春(Cortusamatthioli)等。
3.2 华北落叶松胸径分析
华北落叶松总体的均值(11.03cm)和活立木个体的均值(11.54cm)相差很小,枯立木个体的均值(8.27cm)与活立木以及总体相差很大(图1)。把华北落叶松不同胸径根据大小依次划分为5个等级,计算不同胸径区间内个体数占总体数的比例,结果表明:活立木个体在胸径小于10cm内数量最多,而且从图2线图和柱状图都可以看出从胸径值小的区间到胸径值大的区间活立木呈现显著的递减趋势。枯立木个体胸径只在小于15cm范围内分布,但在胸径小于10cm内分布较多(图2)。
3.3 华北落叶松的完全空间随机模型和空间异质性过程模型模拟分析
利用完全空间随机模型对0—25m尺度上的华北落叶松格局进行分析发现:活立木个体在0.5、2.5m和21m上大于上包迹线为聚集分布,在14.5cm上小于下包迹线为均匀分布;枯立木个体在0—15.5m上大于上包迹线为聚集分布,22.5m上小于下包迹线为均匀分布(图3)。
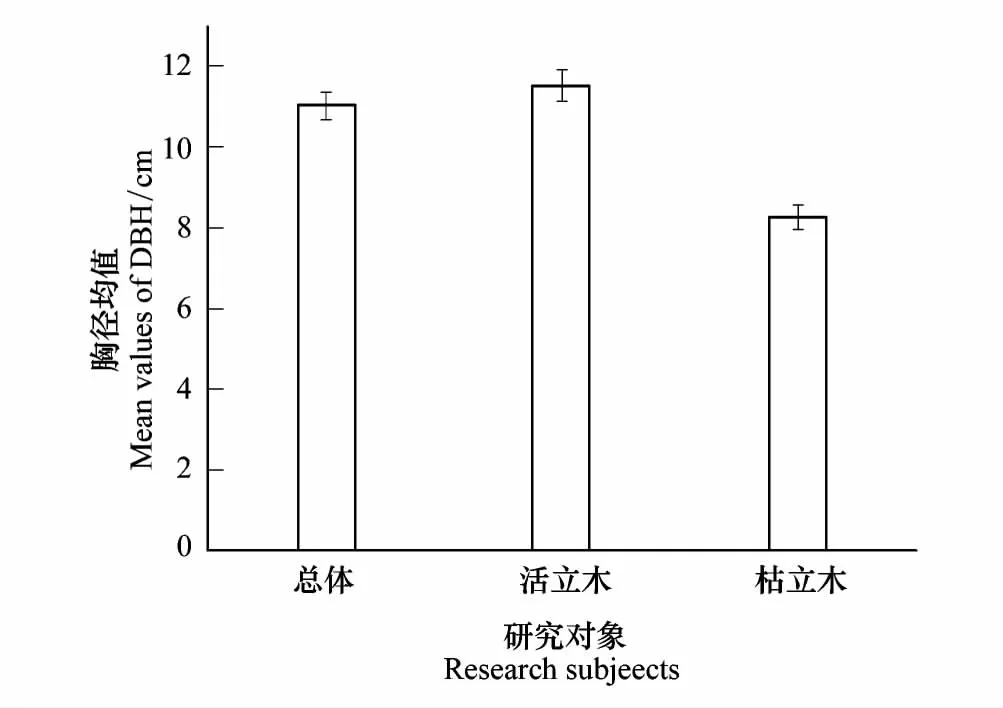
图1 华北落叶松胸径均值Fig.1 Average DBH of Larix principis-rupprechtii
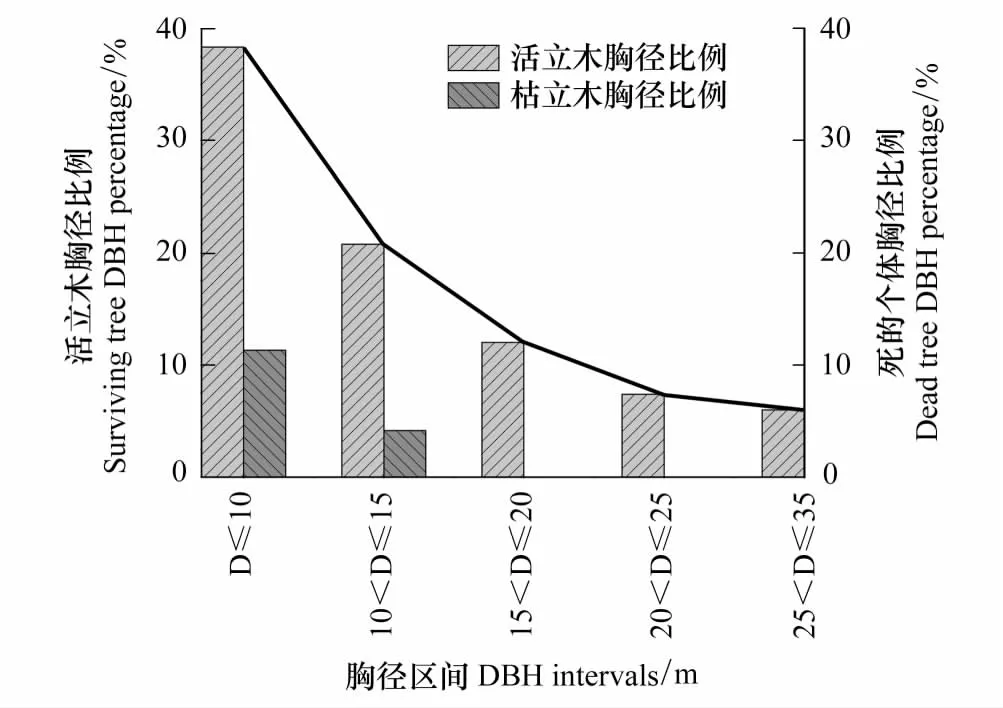
图2 活立木和枯立木的胸径比例Fig.2 DBH ratio of survival and dead tree
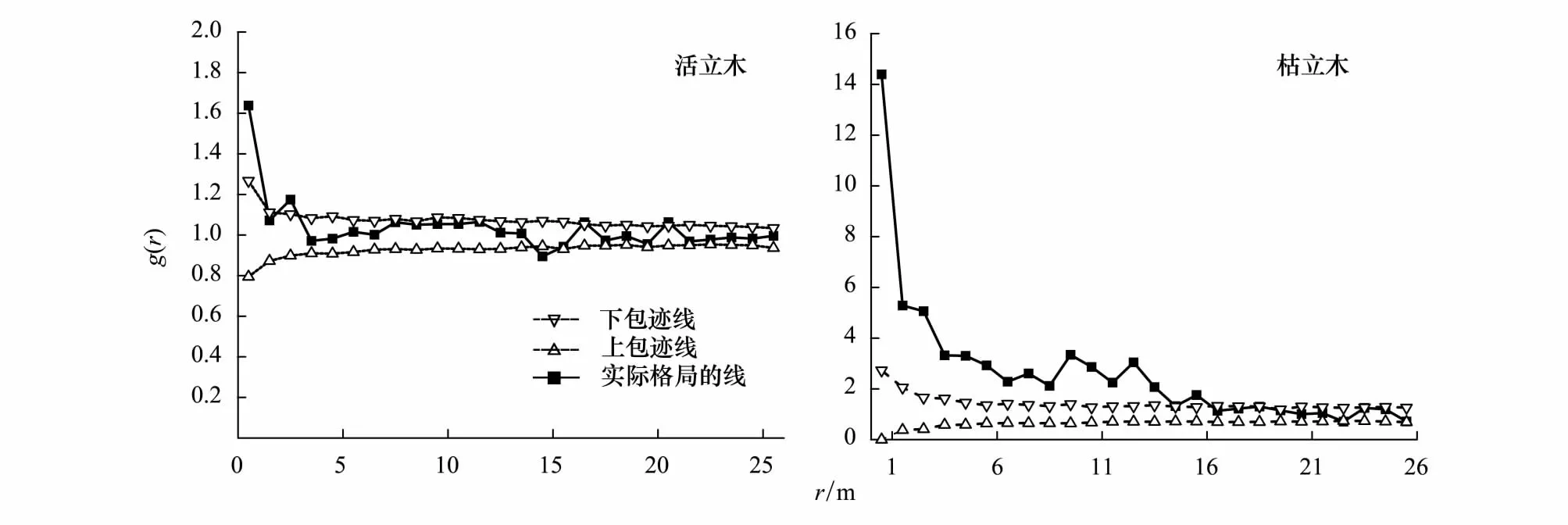
图3 华北落叶松活立木和枯立木的格局与尺度分析Fig.3 Pattern on scales analysis of survival and dead Larix principis-rupprechtii
以华北落叶松活立木的分布密度作为空间异质性模型的一次特征值,选择带宽3m,Kernel方法,在0—25m上的空间异质性模拟结果表明:活立木个体格局值大部分落在空间异质性过程模型的包迹线内且HP模拟值曲线与实际格局值曲线走势相似,空间异质性模型可以通过显著水平95%检验(P=0.065)很好模拟活华北落叶松个体格局。枯立木个体的格局大部分值落在空间异质性过程模型的包迹线外且HP模拟值的曲线与实际格局值曲线相差很大,实际格局与空间异质性过程模型模拟格局的差异检验结果的概率值(P=0.005)小于0.05,说明枯立木格局不能被现在活立木分布为参考搭建的空间异质性模型的一次特征值的空间异质性过程模型所解释(图4)。
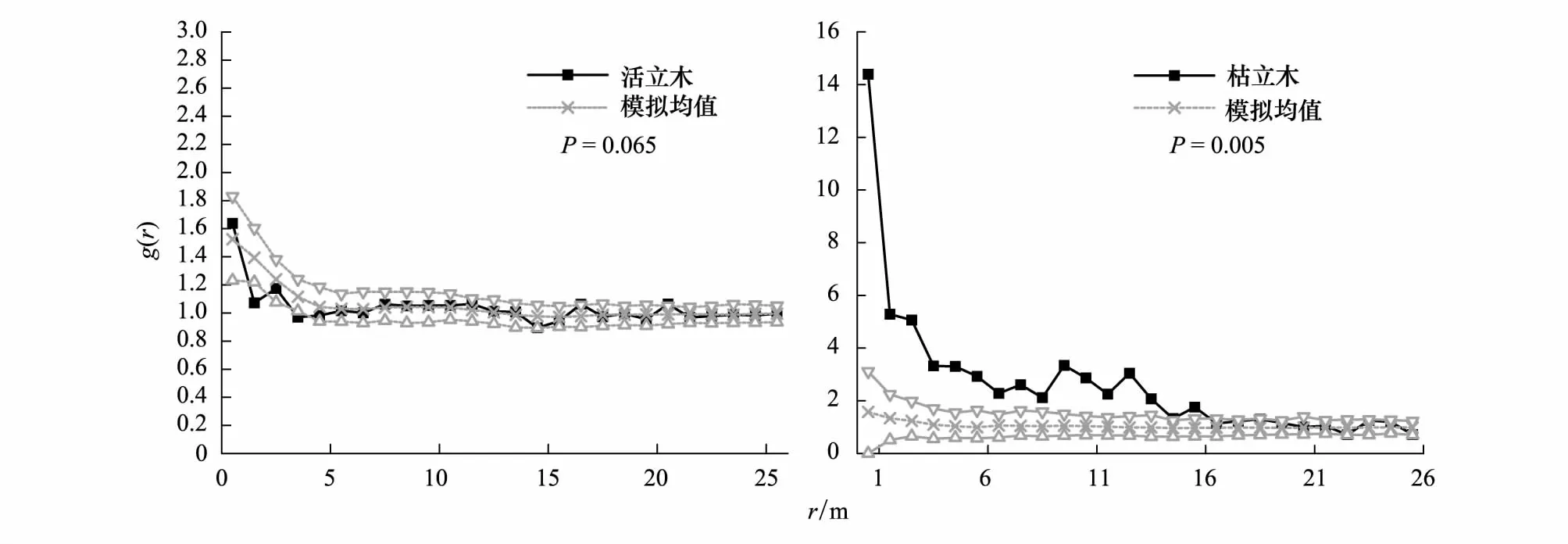
图4 空间异质性过程模型模拟实际格局Fig.4 Heterogeneous Poisson process simulated observed patternHP 指空间异质性过程(Heterogeneous Poisson process)
3.4 随机标签(Random labeling)模型对华北落叶松枯立木、活立木格局的分析
采用随机标签对活的华北落叶松个体和死的华北落叶松个体进行分析,成对相关函数g12除在21m附近出现负相关外,在0—25m尺度上都在包迹线内,说明活立木个体与枯立木个体大部分距离内无显著相关关系,在一定距离(21m)处才会出现负相关。g21-g11在0—15m尺度上小于下包迹线,在活立木个体附近的枯立木个体的邻体密度低于枯立木个体附近的枯立木个体的邻体密度。g12-g22在3m处大于上包迹线,表示枯立木个体周围的活立木个体的邻体密度高于在活立木个体周围的活立木个体的邻体密度;21m处小于下包迹线,表示在枯立木个体周围的活立木个体的邻体密度低于在活立木个体周围的活立木个体的邻体密度,其他尺度上差异不显著。g1,1+2-g2, 1+2在0—12m尺度上大于上包迹线,枯立木个体周围的华北落叶松(死+活)邻体密度高于活立木个体周围华北落叶松(死+活)邻体密度,华北落叶松枯立木周围密度高于活立木周围的密度,0—15m上枯立木聚集分布,因此0—12m内高密度的华北落叶松在枯立木周围是华北落叶松死亡的原因(图5)。

图5 随机标签模型分析0—25m尺度上活立木、枯立木的相关关系Fig.5 Random labeling simulated relation of survival and dead tree on 0—25m scales“1 ” 代表枯立木 dead individuals,“2 ”代表活立木 surviving individuals
4 讨论
4.1 林线过渡带内华北落叶松种群特征
华北落叶松在研究样地内的平均胸径为11.54cm,而参考相关对华北落叶松人工林生长规律的研究表明,华北落叶松胸径年平均年生长量为4.6cm[31],研究样地内树龄大于30年,计算得到现在它的胸径至少应该是13.8cm,可见林线过渡带内的华北落叶松生长速度变慢。而且随胸径径级的增加活立木的数量越来越少,这说明随着年龄的增加林线内的华北落叶松胸径生长变慢也是其林线环境做出的适应特点之一。枯立木的胸径在8.27cm,说明不是最近几年的死亡,枯立木的胸径分布在小于15cm内尤其在小于10cm内比例最高,而华北落叶松成熟年龄在43年左右,说明幼苗变幼树的过程中死亡率最高,这与青藏高原东缘林线乔木种群生长特征有相似的结果[32]。
4.2 林线过渡带内华北落叶松活立木、枯立木的格局与尺度的响应
针对0—25m个体距离为研究尺度,我们发现活立木在3m内有显著聚集分布,华北落叶松冠幅比较大,猜测为幼树在这个尺度内聚集分布,这样可以受到成树的庇护[33],这是适应恶劣环境的对策之一[34],在21m上也有聚集分布,可能是成树的聚集尺度。在15m附近有均匀分布出现,可能成树之间在这个尺度上更容易生存。枯立木的聚集分布很明显,在15m以内都是聚集的,显然死亡不是随机的,这与青藏高原林线内冷杉格局研究结果支持的随机死亡假说不一致[35]。利用活立木分布模拟的空间异质性研究结果表明,环境异质性对活立木格局形成有显著影响[36-37],环境异质性模拟的格局与枯立木的格局不吻合,说明死亡主要不是现有的空间异质性导致的,在林线这种敏感的环境中任何微小的环境改变都会导致树木的死亡[38],过去的空间异质性可能是导致华北落叶松死亡的原因。尤其是气候变动大的年份等,这需要结合气候数据进行进一步分析验证。
4.3 随机标签对华北落叶松死亡的生态学推测
对活立木和枯立木的相关性在0—25m尺度上分析表明死的华北落叶松与活的华北落叶松在21m尺度附近有显著的负相关,说明现有活立木对枯立木的种内排斥作用出现在21m附近。在0—15m上活立木个体周围的枯立木比枯立木周围的枯立木出现概率低,说明该尺度范围内枯立木周围的死亡率大于活立木周围的死亡率,即在这个尺度范围内华北落叶松的死亡个体很集中,但在15m内现有活立木不对枯立木产生排斥作用,推测现有枯立木出现的原因和现有活立木的分布无关。21m处活立木本身是聚集格局而枯立木自身是随机格局,因此大于枯立木周围的活立木,3m处活立木和枯立木本身格局都是聚集格局,但是枯立木周围的活立木出现概率却大于活立木周围的活立木的出现概率,而0—15m上(含3m)活立木周围的枯立木出现概率低,综合二者结论说明枯立木被活立木所包围,活立木密度在3m距离尺度内大于枯立木的密度。说明活的华北落叶松在林线周围聚集分布且聚集强度在3m内大于死的华北落叶松的聚集分布,推测抱团生活更适应林线生境。最后对枯立木周围华北落叶松的总体与活立木周围华北落叶松总体出现概率的比较表明总体向枯立木方向聚拢发展,而且枯立木聚集分布范围也很广,推测南台林线的华北落叶松存在负密度制约效应这与最近对森林研究的结论一致[39-40],即华北落叶松密度越高存活率降低,后期死亡风险越大。相关研究表明:幼树阶段的负密度制约效果明显,环境异质性对灌木的幼苗作用有限[41]。根据负密度制约假说分析[41,42],上面分析推测种内排斥在0—15m内不是枯立木出现的原因,而作为灌木的华北覆盆子(Rubusidaeus)、瘤糖茶藨子(Ribeshimalense)在15m内对华北落叶松的幼苗或者幼树的种间竞争可能是死亡率增大的原因,也有相关研究提到林线内鬼箭锦鸡儿(Caraganajubata)的盖度通常在40%以上,有时甚至达到90%,可能影响了乔木的生长[22]。因此灌木对乔木格局的影响有待研究验证。
5 结论
林线过渡带生境内的华北落叶松生长缓慢,死的个体胸径都集中在幼苗到幼树阶段,活立木胸径随径级增加也呈现递减趋势。死亡个体聚集分布在0—15m上显著,活的个体仅在3m以内和21m处聚集分布,空间异质性对活的华北落叶松有显著作用,但是幼树阶段的华北落叶松的负密度制约作用显著,在现有调查尺度内由华北落叶松的活立木构建的空间异质性不能很好解释华北落叶松的枯立木的格局,但不同林线类型的存在本身就说明大尺度上空间异质性对林线树木的死亡起很大作用。负密度制约效应对死亡率的影响很大,种内排斥作用在近距离内不明显。本文主要针对华北落叶松为对象展开讨论,针对灌木对幼苗和幼树成活的影响有待研究证实。
