海拔变化对高原湿地优势植物叶绿素荧光特性的影响
2018-11-15刘国栋许俊萍岳海涛李丽萍
李 晖,田 昆,刘国栋,王 行,孙 梅,许俊萍,岳海涛,李丽萍,戴 强
1 西南林业大学湿地学院, 昆明 650224 2 西南林业大学国家高原湿地研究中心, 昆明 650224 3 曲阜师范大学地理与旅游学院, 日照 276826
有研究表明气候变化是导致高寒地区湿地生态系统退化的重要原因之一,也是引起环境变劣、生物多样性下降的重要因素[1]。高原湿地是我国重要的生态屏障,对维系流域生态安全有着重要作用[2],由于其地理位置的特殊性及对气候变化的高度敏感性[3],逐渐成为科学家们关注的热点。云南高原湿地地形的相对封闭和地理上的隔离,造成了其在发挥至关重要作用的同时,也成为一个更为敏感和脆弱的地带[4]。湿地植物作为湿地生态系统内物质循环和能量流动的首要生物因素,在维持闭合半闭合湿地生态系统的稳定和物质循环中起着重要作用,是其结构和功能的核心[5],也是湿地其他生物生长和新陈代谢所需能量的主要来源[6]。在自然条件下,气候变化对湿地生态系统的影响是一个长期、复杂的过程,开展气候变化与湿地生态系统耦合关系研究的难点是研究时间尺度的问题。国内外研究者多以室内控制实验来开展湿地植物响应气候变化的研究。云南高原地处低纬度高海拔地区,地理位置特殊,纬度和海拔的共同作用使得气候的区域差异和垂直变化十分明显。海拔梯度变化所带来的区域气候条件差异是否会对适应了高海拔冷凉气候的湿地植物叶绿素荧光特性产生影响,以及它们如何去适应或响应这种变化,目前鲜见这方面的研究报道。开展该项研究,以原位移置试验来探讨海拔变化对湿地植物叶绿素光特性的影响,揭示湿地植物对海拔变化的适应与响应机制,补充与完善湿地植物与海拔变化相互作用机制的内涵,为我国高原湿地的保护提供预警或决策依据。
1 材料和方法
1.1 研究区概况
青藏高原东南缘的横断山地势由北向南倾斜,南北走向带上沿海拔梯度下降孤立分散着许多高原湿地,横断山腹心地带的纳帕海湿地是该区域代表性湿地,也是国内外生物多样性保护关注的热点区域[7],纳帕海湿地位于长江流域,海拔3260m,年平均气温5.4℃,年平均降水量620mm,干湿季分明,分布着世界广泛生长的植物水葱(Scirpustabernaemontani)和茭草(Zizaniacaduciflora),适应其冷凉气候,这两种植物成为了纳帕海湿地的优势植物群落。随着海拔降低,同一走向带上分布着另一个典型湿地拉市海,海拔2400m,年平均气温11.8℃,年降水量在900—1200mm,也是干湿季分明,其中6—9月降水量占全年的80%以上[8]。地势继续往南倾斜降至海拔1891m的代表性湿地滇池,其年平均气温升至15.1℃,年均降雨量1075mm,夏季雨量占全年降水量的60%以上,也具有干湿季分明的特点,这种随海拔梯度变化引起的气候因子差异为位移试验研究提供了条件。不同海拔梯度主要气象因子特征见表1, 研究区域位置见图1。
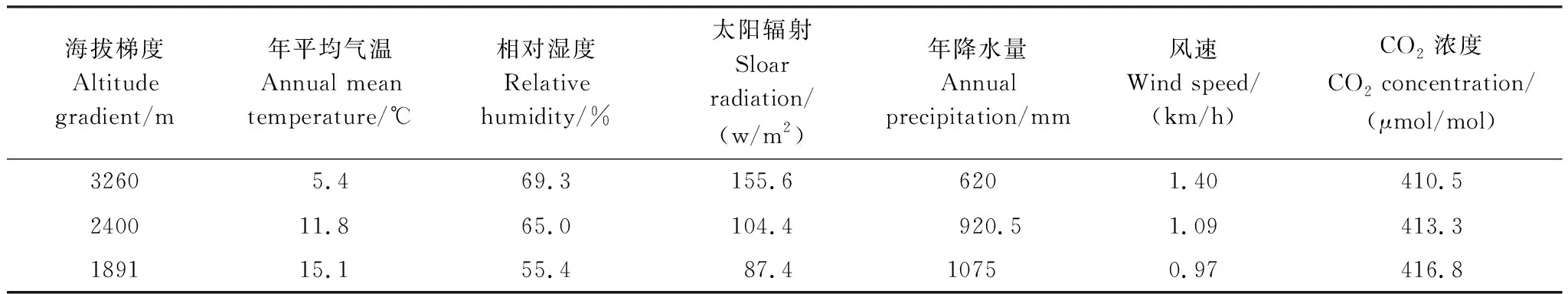
表1 不同海拔梯度主要气象因子特征
图1 研究区域位置图Fig.1 Location of the study sites
1.2 实验材料和实验设计
利用云南北高南低的立体地形及其海拔差异引起的气候条件变化,以纳帕海世界广泛分布的代表性优势植物水葱(Scirpustabernaemontani)和茭草(Zizaniacaduciflora)群落为研究对象,随其生长土壤移至低海拔地区,研究其随海拔下降,气候条件改变的生长变化,以位移试验来探讨气候变化对高原湿地植物光合作用的影响。将不低于120株的水葱和茭草的植物-土壤单元分别原位移至海拔依次降低的丽江拉市海和滇池流域西南林业大学校园实验池内,在移出地纳帕海做同样移出试验。三地的实验池条件一致,每个池长300cm,宽150cm,高100cm,土层厚度50cm,水深依据移出地自然水位保持在25cm左右。每种参试植物设置3组重复。经过1年左右的适应期,以移出地纳帕海为对照,于植物生长的关键时期分别测量三地实验池内植物萌芽期、生长期和凋落期的叶绿素含量、叶绿素荧光参数和叶绿素荧光光响应曲线数据,同时,在3个实验地分别安装全天候气候自动记录仪(Portlog,USA),同步监测气象特征因子。分析比较随海拔梯度下降的不同区域气候因子中的关键因子,以及随区域气候条件变化的植物光合特征差异。
1.3 测定指标与方法
叶绿素含量的测定:在三地植物生长的萌芽期、生长期和凋落期内,选取标记过的植物,齐根剪下,用密封袋装好置于便携式冷冻箱中带回实验室,用蒸馏水洗净,剪碎后称取0.2g装入25mL试管中,加80%乙醇10mL左右,加塞放入60—80℃水浴中恒温提取叶绿素,至叶片全部褪绿为止,冷却后,用80%乙醇定容至刻度,摇匀后分别于663nm和645nm波长下测定吸光值,而后带入公式计算出叶绿素a、叶绿素b及总叶绿素的含量[9]。
叶绿素荧光参数的测定:在三地植物生长的萌芽期、生长期和凋落期内,选择晴朗的天气,于9:00—11:30,使用Li- 6400XT便携式光合仪(Li-COR,Lincoln,NE,USA),选取标记过的植物,分别进行实际光化学量子效率(PhiPSⅡ)、电子传递速率(ETR)、光化学猝灭系数(qP)及非光化学猝灭系数(NPQ)的测定;于太阳落山1h后,进行光化学量子效率(Fv/Fm)的测定。测定时,叶室内的测量光强设定为1000μmol m-2s-1,人工控制CO2浓度为400μmol/mol。
叶绿素荧光光响应曲线的测定:在三地植物生长的萌芽期、生长期和凋落期内,选择晴朗的天气,于9:00—11:30,利用LI- 6400XT便携式光合仪(Li-COR,Lincoln,NE,USA),选取标记过的植物,进行叶绿素荧光光响应曲线的测量,光强梯度从2000μmol m-2s-1开始,依次降为1500、1000、500、200、100μmol m-2s-1和50μmol m-2s-1,人工控制CO2浓度为400μmol/mol。
1.4 数据分析
采用Excel 2010及SPSS 20.0进行相关数据分析,不同数据间的差异比较运用单因素方差分析法(One way ANOVA)在Sig=0.05显著水平下进行。叶绿素荧光光响应曲线运用Origin软件拟合曲线。不同气候因子与植物叶绿素荧光特性各参数之间的关系采用主成分分析法进行相关性分析。
2 结果与分析
2.1 不同生长期三地主要气候因子分析
影响湿地植物生长的因素很多,随海拔梯度变化,植物生长环境因子也相应不同,在考虑了消除土壤、水质等主要因子影响的基础上,气候因子中太阳辐射、降雨量、相对湿度、温度变化和空气CO2浓度变化等均是影响植物光合生长的主要因子,利用设置于3个研究地的全天候气候自动记录仪,通过同步观测记录到的气象数据,分析其对湿地植物光合生长的影响及其贡献率。采集到的气象数据表明,纳帕海、拉市海和昆明三地的年平均气温分别为7.13℃、13.3℃和16.9℃,在植物生长周期内,相邻两地相差最大4.4℃,最低0.6℃(表2),年降雨量分别为564.9、966.3mm和971.8mm,与50年来三地的气候变化趋势一致。不同生长期内,三地植物的气候因子呈现出随海拔降低气温上升、太阳辐射下降和空气CO2浓度升高的趋势,且气温和空气CO2浓度上升规律明显,相邻两地气温上升最高4.4℃,与IPCC本世纪末气候变化温度预测值接近,而降雨和相对湿度变化规律不明显(表2)。

表2 不同生长期三地主要气候因子差异
2.2 水葱、茭草叶绿素含量差异
水葱和茭草移至拉市海和滇池后,随着海拔降低,区域气候条件发生改变,与移出地纳帕海相比,在植物萌芽期,拉市海和滇池的水葱叶绿素a分别增加了10.2%和50.5%,叶绿素b分别增加了8.66%和23.47%、总叶绿素含量分别增加了22.54%和68.77%,叶绿素a/b分别增加了1.42%和21.91%(图2)。茭草的叶绿素a分别增加了45.19%和87.88%、叶绿素b分别增加了16.77%和35.06%、总叶绿素含量分别增加了11.17%和29.2%,叶绿素a/b分别增加了12.99%和26.4%(图3)。
两种植物生长期和凋落期叶绿素含量的变化与萌芽期相同,但两种植物生长期这些数值的增长率较萌芽期高且变化明显(图2、图3),凋落期这些数值的增长率则较萌芽期低(图2、图3)。相较移出地纳帕海,生长期拉市海和滇池水葱叶绿素a的增长率比萌芽期分别高出22.83%和50.92%,叶绿素b的增长率分别高出12.99%和15.66%,总叶绿素含量的增长率分别高出12.16%和24.16%,叶绿素a/b的增长率分别高出7.93%和11.02%,茭草叶绿素a的增长率比萌芽期分别高出37.56%和45.38%,叶绿素b的增长率分别高出20.11%和27.91%,总叶绿素含量的增长率分别高出13.77%和16.46%,叶绿素a/b的增长率分别高出20.51%和16.72%。凋落期水葱叶绿素a的增长率比萌芽期分别下降6.18%和24.87%,叶绿素b的增长率分别下降5.22%和2.39%,总叶绿素含量的增长率分别下降5.91%和20.5%,叶绿素a/b的增长率分别下降0.85%和18.15%,茭草叶绿素a的增长率分别降低25.71%和41.59%,叶绿素b的增长率分别降低7.21%和13.78%,总叶绿素含量的增长率分别降低9.78%和14.26%,叶绿素a/b的增长率分别降低3.94%和5.78%。可见,两种植物3个时期的叶绿素含量都随区域气候条件的改变而增加,光合能力增强,且海拔较低的滇池增加速率更大。
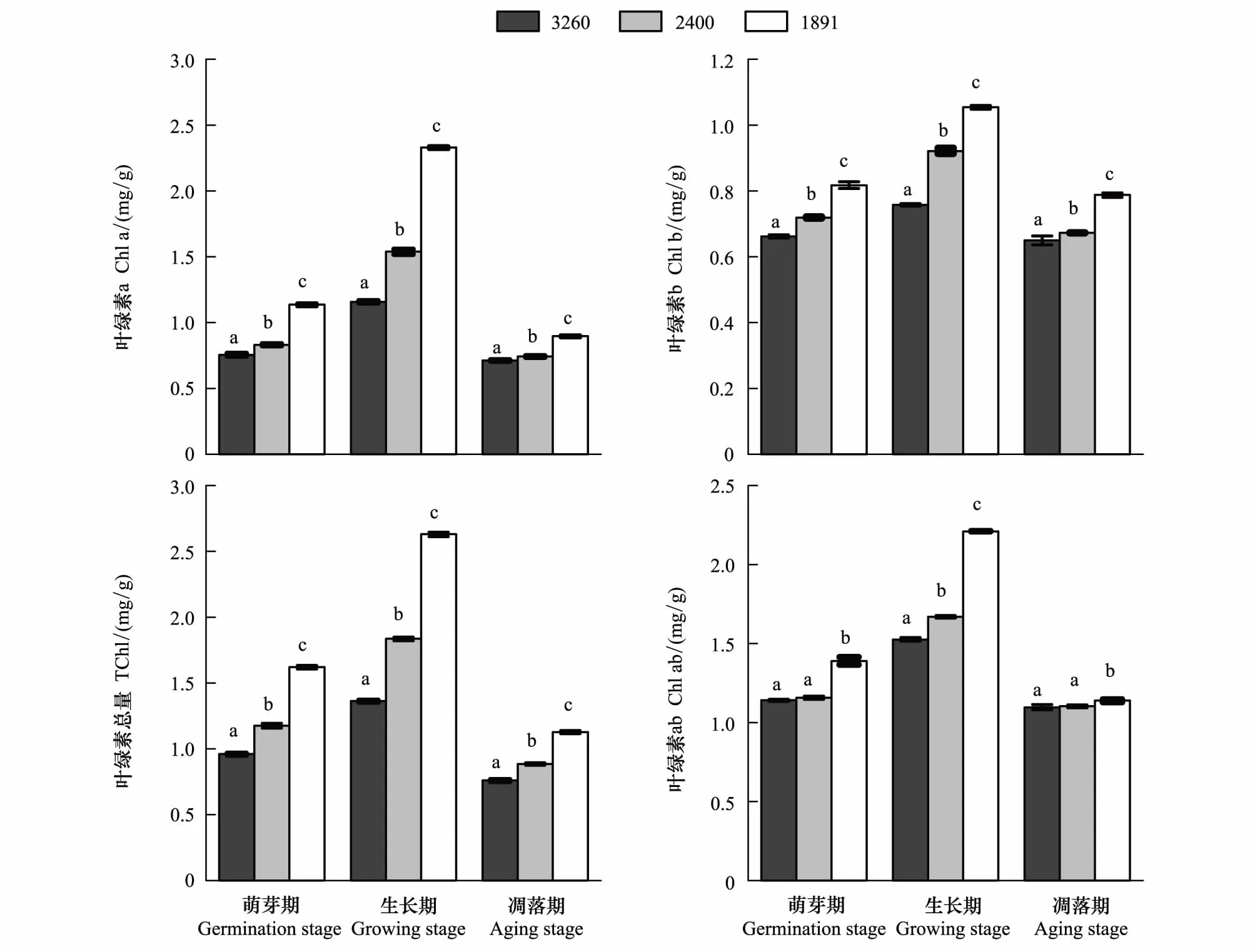
图2 不同海拔梯度水葱生长周期的叶绿素含量变化(平均值±标准差)Fig.2 Changes in chlorophyll content of Scirpus tabernaemontani growth cycle stages at different altitude (mean ± SD)不同海拔相同生长期标有不同英文字母,表明两者间差异显著(P<0.05)
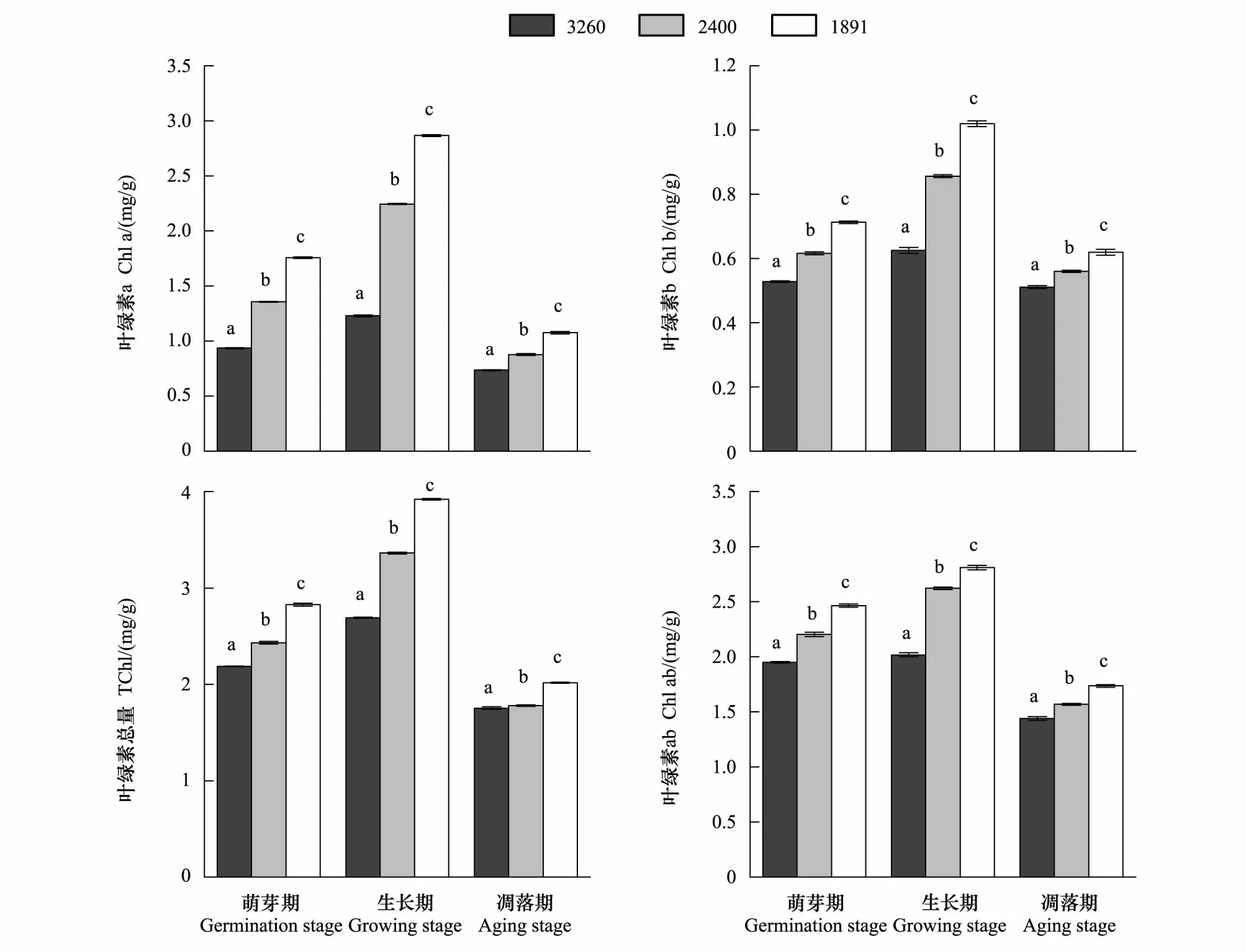
图3 不同海拔梯度茭草生长周期的叶绿素含量变化(平均值±标准差)Fig.3 Changes in chlorophyll content of Zizania caduciflora growth cycle stages at different altitude (mean ± SD)不同海拔相同生长期标有不同英文字母,表明两者间差异显著(P<0.05)
2.3 水葱、茭草叶绿素荧光参数变化
与移出地纳帕海相比,在植物萌芽期,移至拉市海和滇池的水葱Fv/Fm值分别增加了1.99%和5.98%,PhiPSⅡ值分别增加了36.65%和95.78%,ETR值分别增加了36.71%和96.17%,qP值分别增加了32.17%和67.55%,NPQ值则分别减少了17.13%和59.1%(图4)。茭草的Fv/Fm分别增加了9.4%和12.06%,PhiPSⅡ值分别增加了43.52%和87.29%,ETR值分别增加了42.71%和87.12%,qP值分别增加了23.59%和51.47%,NPQ值分别减少了19.28%和34.09%(图5)。
生长期和凋落期两种植物的叶绿素荧光参数均随着区域气候条件的改变而增加,但生长期这些数值的增长率或下降幅度比萌芽期高,凋落期则较萌芽期低(图4、图5)。生长期拉市海和滇池的水葱Fv/Fm值分别增加了3.05%和6.77%,PhiPSⅡ值分别增加了69.61%和103.55%,ETR值分别增加了50.35%和103.19%,qP值分别增加了65.55%和92.51%,NPQ值分别减少了52.74%和62.6%,茭草的Fv/Fm分别增加了10.66%和15.16%,PhiPSⅡ值分别增加了80.72%和119.43%,ETR值分别增加了80.77%和119.3%,qP值分别增加了30.46%和72.74%,NPQ值分别减少了29.87%和50.76%。凋落期水葱的Fv/Fm值分别增加了1.46%和3.83%,PhiPSⅡ值分别增加了17.13%和90.7%,ETR值分别增加了17.14%和90.53%,qP值分别增加了9.82%和49.01%,NPQ值分别减少了8.28%和42.77%,茭草的Fv/Fm分别增加了5.33%和10.91%,PhiPSⅡ值分别增加了10.41%和35.35%,ETR值分别增加了10.32%和35.54%,qP值分别增加了13.02%和41.63%,NPQ值分别减少了8.33%和10.48%。
由于水葱和茭草存在种间差异以及两种植物的适应能力不同,两者叶绿素荧光参数值的增长幅度与下降幅度不同。随海拔降低,两种植物3个时期的叶绿素荧光参数也随区域气候条件改变而升高或降低,这就导致PSⅡ反应中心的最大光能转化效率和捕光能力升高,吸收的光能更多地用于光合作用,提高了两者的光合效率。
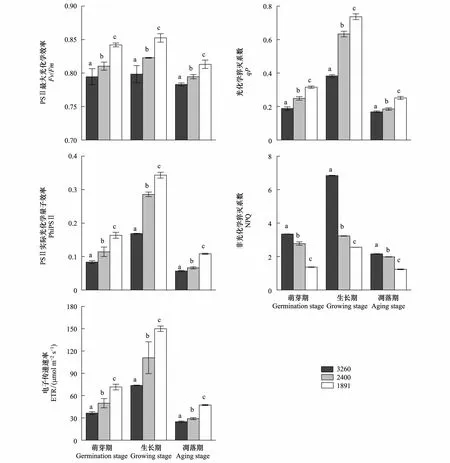
图4 不同海拔梯度水葱生长周期的叶绿素荧光参数变化(平均值±标准差)Fig.4 Changes in chlorophyll fluorescence parameters of Scirpus tabernaemontani growth cycle stages at different altitude (mean±SD)不同海拔相同生长期标有不同英文字母,表明两者间差异显著(P<0.05)
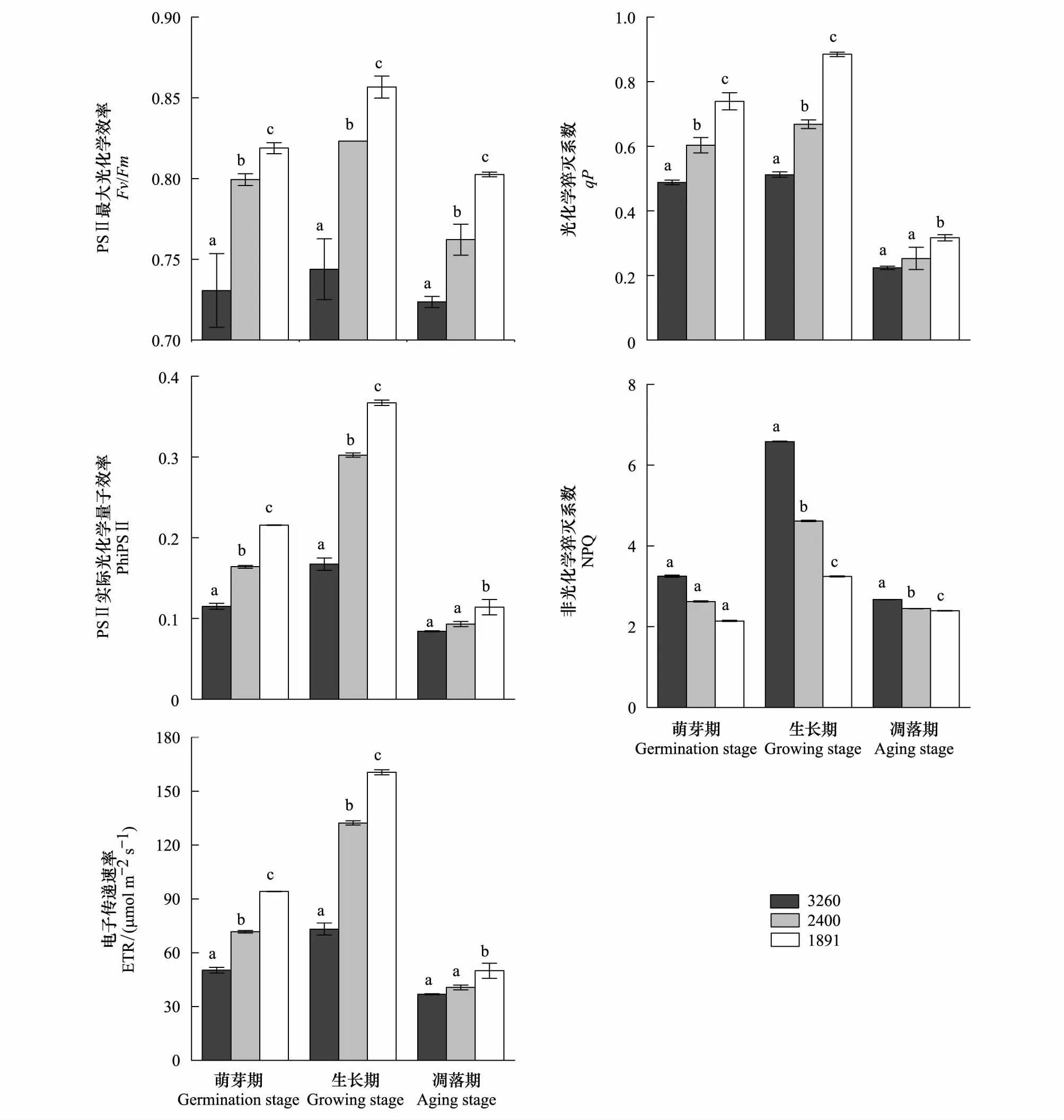
图5 不同海拔梯度茭草3个生长期的叶绿素荧光参数变化(平均值±标准差)Fig.5 Changes in chlorophyll fluorescence parameters of Zizania caduciflora growth cycle stages at different altitude (mean ± SD)不同海拔相同生长期标有不同英文字母,表明两者间差异显著(P<0.05)
2.4 水葱、茭草叶绿素荧光光响应曲线变化
植物萌芽期,三地水葱和茭草的PhiPSⅡ-PAR和PhiCO2-PAR曲线都为反比例函数(图6)。水葱和茭草的三条PhiPSⅡ-PAR和PhiCO2-PAR曲线在50至500之间都呈急速下降趋势,在500至2000之间呈缓慢下降趋势。随海拔降低,区域气候条件改变,与移出地纳帕海相比,在植物萌芽期,移至拉市海和滇池流域的水葱PhiPSⅡmax分别增加了13.96%和24.09%,PhiCO2max分别增加了89.87%和114.77%,茭草的PhiPSⅡmax分别增加了25.42%和46.48%,PhiCO2max分别增加了27.08%和84.33%,茭草的PhiPSⅡmax增加率较水葱高,而PhiCO2max增加率却比水葱低,由于PhiPSⅡ与植物的叶绿素含量成正比,PhiCO2与CO2浓度和气孔导度有关,而茭草的叶绿素含量比水葱多,气孔导度却比水葱低,导致茭草叶片中CO2的含量比水葱低,所以茭草的PhiPSⅡmax增加率比水葱高,而PhiCO2max增加率却比水葱低。三地水葱和茭草的PhiPSⅡ和PhiCO2存在线性关系,滇池、拉市海和纳帕海水葱的PhiPSⅡ和PhiCO2的相关系数分别为0.962、0.994和0.986,茭草的PhiPSⅡ和PhiCO2的相关系数分别为0.967、0.984和0.998。水葱和茭草所表现出的差异说明两者的适应能力不同。
两种植物生长期和凋落期曲线和参数的变化均呈上升趋势,与萌芽期相同,不同的是两种植物生长期PhiPSⅡmax和PhiCO2max的增长幅度比萌芽期高且变化显著(图7),而凋落期PhiPSⅡmax和PhiCO2max的增长幅度却比萌芽期低(图8),这与生长期水热条件好且植物处于快速生长阶段,而凋落期植物处于衰落阶段有关。在植物生长期,移至拉市海和滇池流域的水葱PhiPSⅡmax分别增加了21.68%和30.34%,PhiCO2max分别增加了120.46%和153.94%,茭草的PhiPSⅡmax分别增加了36.03%和54.81%,PhiCO2max分别增加了54.62%和15.67%。凋落期水葱PhiPSⅡmax分别增加了7.23%和20.46%,PhiCO2max分别增加了64.37%和88.71%,茭草的PhiPSⅡmax分别增加了12.07%和24.90%,PhiCO2max分别增加了21.95%和80.48%。
两种植物生长期和凋落期的PhiPSⅡ和PhiCO2也存在线性关系。植物生长期,滇池、拉市海和纳帕海水葱的PhiPSⅡ和PhiCO2的相关系数分别为0.958、0.998和0.978,茭草的PhiPSⅡ和PhiCO2的相关系数分别为0.912、0.965和0.923。植物凋落期,水葱的PhiPSⅡ和PhiCO2的相关系数分别为0.976、0.968和0.914,茭草的PhiPSⅡ和PhiCO2的相关系数分别为0.988、0.989和0.972。原因与植物的叶绿素含量、植物所处的环境CO2浓度及植物的气孔导度有关,不同的相关系数说明PhiPSⅡ和PhiCO2的相关程度不同以及线性拟合的好坏。
随着海拔降低,两种植物3个时期的PhiPSⅡ-PAR、PhiCO2-PAR曲线和PhiPSⅡmax、PhiCO2max值依次升高,致使植物的光合活性提高,光合能力得到增强,为种群的发展壮大提供了有利条件。

图6 3个不同海拔梯度萌芽期水葱和茭草PhiPSⅡ-PAR、PhiCO2-PAR曲线(平均值±标准差)Fig.6 PhiPSⅡ-PAR and PhiCO2-PAR curves of Scirpus tabernaemontani and Zizania caduciflora in germination stage at different altitude (mean±SD)

图7 3个不同海拔梯度生长期水葱和茭草PhiPSⅡ-PAR、PhiCO2-PAR曲线(平均值±标准差)Fig.7 PhiPSⅡ-PAR and PhiCO2-PAR curves of Scirpus tabernaemontani and Zizania caduciflora in growing period at different altitude (mean±SD)

图8 3个不同海拔梯度凋落期水葱和茭草PhiPSⅡ-PAR、PhiCO2-PAR曲线(平均值±标准差)Fig.8 PhiPSⅡ-PAR and PhiCO2-PAR curves of Scirpus tabernaemontani and Zizania caduciflora in withered period at different altitude (mean±SD)
2.5 气候因子与植物荧光特性相关性分析
对三个气候条件下水葱和茭草的叶绿素荧光特性与气候因子进行相关性分析,结果表明:植物生长周期水葱和茭草的各项指标的变化与降雨量和相对湿度相关性较小(表3),由于两种植物均生长于水中,因此降雨量和相对湿度不是影响植物生长各项指标变化的最主要的气象因子。而植物生长周期水葱和茭草的叶绿素a主要与气温(r=0.796,P<0.01;r=0.737,P<0.01)、太阳辐射(r=0.699,P<0.01;r=0.734,P<0.01)和空气CO2浓度(r=0.629,P<0.01;r=0.719,P<0.01)呈极显著正相关,叶绿素b主要与气温(r=0.821,P<0.01;r=0.852,P<0.01)、太阳辐射(r=0.716,P<0.01;r=0.687,P<0.05)和空气CO2浓度(r=0.718,P<0.01;r=0.700,P<0.01)呈极显著或显著正相关,总叶绿素主要与气温(r=0.775,P<0.01;r=0.747,P<0.01)、太阳辐射(r=0.726,P<0.01;r=0.717,P<0.05)和空气CO2浓度(r=0.6719,P<0.01;r=0.617,P<0.01)呈极显著或显著正相关,叶绿素a/b与气温(r=0.751,P<0.01;r=0.693,P<0.01)、太阳辐射(r=0.674,P<0.01;r=0.561,P<0.01)和空气CO2浓度(r=0.544,P<0.01;r=0.408,P<0.01)呈极显著正相关,Fv/Fm与气温(r=0.770,P<0.01;r=0.725,P<0.01)、太阳辐射(r=0.569,P<0.01;r=0.589,P<0.01)和空气CO2浓度(r=0.387,P<0.01;r=0.539,P<0.01)呈极显著正相关,PhiPSⅡ与气温(r=0.828,P<0.01;r=0.752,P<0.01)、太阳辐射(r=0.714,P<0.01;r=0.735,P<0.01)和空气CO2浓度(r=0.571,P<0.01;r=0.650,P<0.01)呈极显著正相关,ETR与气温(r=0.799,P<0.01;r=0.752,P<0.01)、太阳辐射(r=0.685,P<0.01;r=0.715,P<0.05)和空气CO2浓度(r=0.598,P<0.01;r=0.650,P<0.01)呈极显著或显著正相关,qP与气温(r=0.852,P<0.01;r=0.654,P<0.05)、太阳辐射(r=0.687,P<0.01;r=0.495,P>0.05)和空气CO2浓度(r=0.507,P<0.01;r=0.384,P<0.05)呈极显著、显著正相关或不相关,NPQ与气温(r=0.562,P<0.01;r=0.627,P<0.01)、太阳辐射(r=-0.017,P<0.01;r=0.115,P<0.01)和空气CO2浓度(r=-0.520,P<0.01;r=-0.445,P<0.05)呈极显著正相关、极显著负相关或显著负相关,PhiCO2与气温(r=0.636,P<0.01;r=0.717,P<0.01)、太阳辐射(r=0.727,P<0.01;r=0.662,P<0.01)和空气CO2浓度(r=0.512,P<0.01;r=0.407,P<0.01)呈极显著正相关(表3)。可见气温、太阳辐射、CO2浓度是影响植物光合最主要的气候因子。
3 讨论
自然条件下,气候变化对湿地生态系统的影响是一个长期、复杂的过程。叶绿素作为光合色素之一,参与光合作用中光能的吸收、传递和转化,其含量的多少间接反映了叶片光合能力的强弱[10]。随着海拔下降,区域气候条件发生改变,水葱和茭草的叶绿素a含量、叶绿素b含量、总叶绿素含量和叶绿素a/b含量也随之增加,郑有飞[11]、周宁[12]、姬江莉[13]和刘杰才[14]等研究发现,温度、太阳辐射、空气CO2浓度、降雨量和相对湿度都会对植物的叶绿素含量产生影响,但湿地植物与气候变化的研究表明降雨量和相对湿度并不是影响因素,而太阳辐射、空气CO2浓度对植物叶绿素含量的影响要小于温度。因此温度的升高是其最主要的环境影响因素,这一结果与蒋丽[15]、徐辉[16]、李娟[10]和任飞[17]等的研究发现一致。温度升高提高了湿地植物酶的活性,使叶绿素酸酯增多,促进了叶绿素的合成,最终使湿地植物的叶绿素含量增加。植物光合色素含量的增加,使光合速率提高,光合产物的量也随之增加,有利于植物光合能力的提升。
叶绿素荧光参数是一组用于描述植物光合作用机理和光合生理状况的变量或常数值,反映了植物“内在性”的特点,被视为是研究植物光合作用与环境关系的内在探针。尽管植物叶绿素荧光信号微弱,却富含了大量的光合信息,它与光合作用关联紧密,能够反映出叶片光合能力的强弱[18]。随着区域气候条件发生变化,水葱和茭草的叶绿素荧光参数也相应发生变化,许珍[19]、倪艳利[20]、韩燕青[21]和赵娟[22]等对影响这些指标变化的因素进行研究后发现,温度、太阳辐射、空气CO2浓度和降雨量都是其影响因素,我们的结果显示降雨量和相对湿度不是影响因素,似乎表明湿地植物的水生生境抵消了降雨和湿度的影响。随着区域气候条件改变,水葱和茭草的Fv/Fm、PhiPSⅡ、ETR和qP值增加,NPQ值减少。Fv/Fm、PhiPSⅡ、ETR、qP值的变化与植物叶绿素含量的高低有关,NPQ值的变化与Fv/Fm、PhiPSⅡ、ETR和qP值的变化有关且变化相反,两种植物的叶绿素含量随着温度的升高而升高,Fv/Fm、PhiPSⅡ、ETR和qP值便随着温度的升高而增加,而NPQ值的变化相反,这五个参数中,Fv/Fm的值比较恒定,一般保持在0.8—0.85之间[23],纳帕海湿地植物生长的三个时期和拉市海湿地茭草生长的初期和两种植物生长的末期,它们的Fv/Fm值都低于0.80,表明两种植物在这几个时期内均受到了温度胁迫,但随着温度升高这种胁迫得到缓解和解除。李磊[24]、董瑜[16]、赵雪[25]和任洁[26]对不同植物的叶绿素荧光参数进行了研究,也发现随着温度升高,Fv/Fm、PhiPSⅡ、ETR和qP值都有所提高,NPQ值有所降低。温度升高所引起的这五个参数的升高或降低,是植物对环境变化的一种适应策略,避免植物的光合机构受到损伤,让光能在植物体内的吸收、传递、耗散、分配等加快,光化学效率提高,光合活性提升,植物的光合能力随之增强。
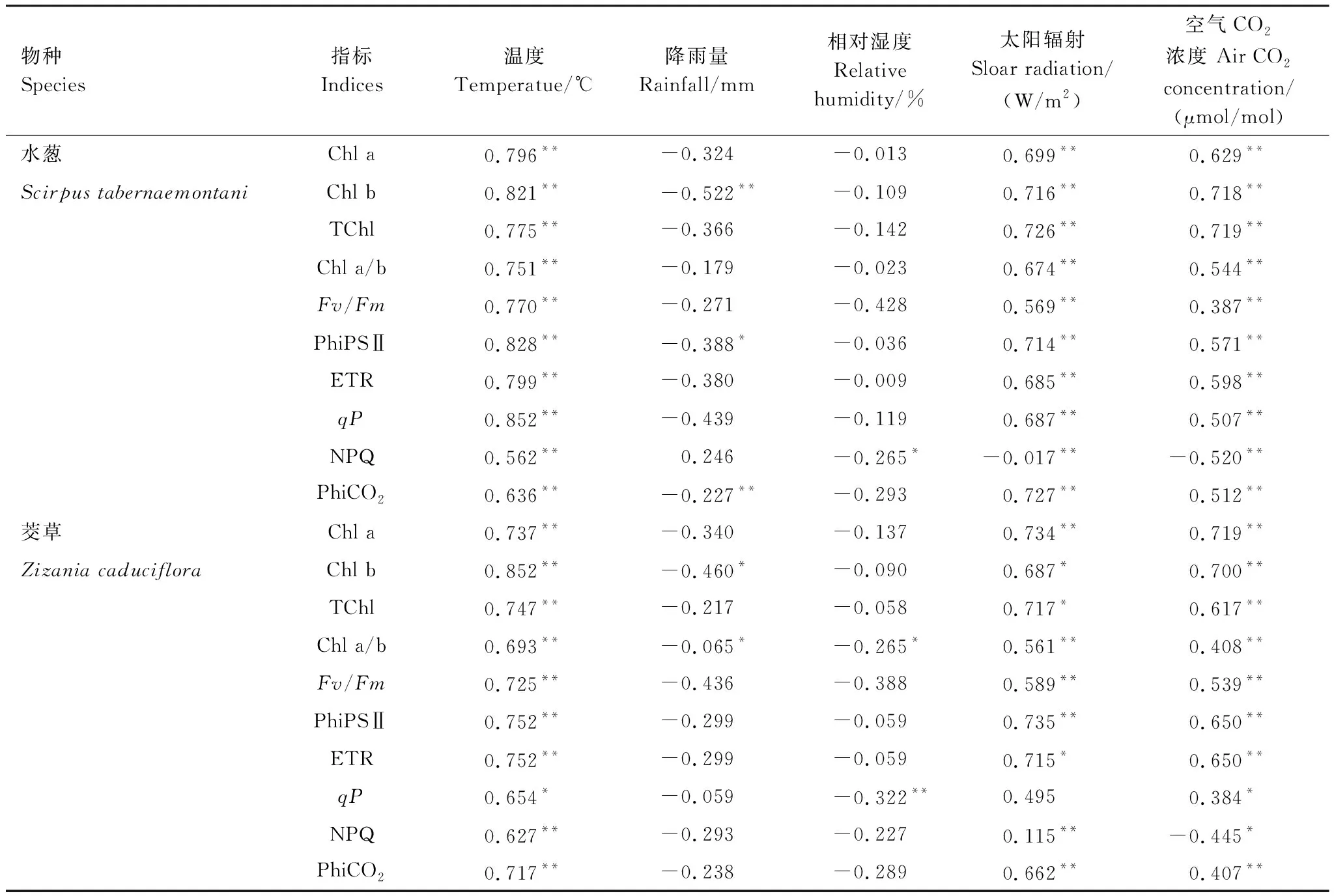
表3 植物生长周期各项指标与环境因子的相关系数
Chl a:叶绿素a,ChlorophyⅡ a;Chl b:叶绿素b,ChlorophyⅡ b;TChl:总叶绿素,Total ChlorophyⅡ;Chl a/b:叶绿素a/b,ChlorophyⅡ a/b;Fv/Fm:PSⅡ最大光化学效率,PSII maximum photochemical efficiency;PhiPSⅡ:PSⅡ实际光化学量子效率,PSII actual photochemical quantum efficiency;ETR:电子传递速率,Electron transfer rate;qP:光化学猝灭系数,Photochemical quenching coefficient;NPQ:非光化学猝灭系数,Non photochemical quenching coefficient;PhiCO2:CO2同化量子产量,CO2assimilation quantum yield;*表示达到0.05显著水平,**0.01极显著水平
叶绿素荧光光响应曲线主要为PhiPSⅡ-PAR和PhiCO2-PAR曲线,PhiPSⅡ-PAR和PhiCO2-PAR曲线不仅能够反映植物在相同光强梯度下PhiPSⅡ和PhiCO2的变化,而且还能够进一步揭示温度升高对植物光化学效率和CO2同化速率相对应的量子产量的影响。在本研究中,随着海拔梯度下降,区域气候条件发生改变,太阳辐射和温度升高使得水葱和茭草的叶绿素含量随之升高,CO2浓度升高,气孔导度亦随之增强,所以PhiPSⅡ和PhiCO2升高。PhiPSⅡ和PhiCO2之间存在一次方函数关系,在两者显著性较强的情况下,R2越大,说明PhiPSⅡ和PhiCO2之间的相关性越强,曲线的拟合程度越好。PhiPSⅡ-PAR和PhiCO2-PAR曲线中PhiPSⅡ和PhiCO2值的升高,说明光化学运行效率加快,同化CO2的能力增强,光合作用能力显著增强。
可见,两种植物随海拔下降的区域气候条件改变,尤其是温度的上升,均表现出较好的适应性,但茭草的适应能力强于水葱,表明茭草捕获和传递的光能以及执行能量转化的光合色素含量都大于水葱,并且茭草每个时期的叶绿素荧光参数区域之间的增幅差或降幅差均小于水葱,说明茭草叶绿素荧光参数的变化较水葱稳定,因此茭草对温度升高的适应能力较水葱强。适应性强的植物的光合速率会提高,光合产物的量会随之增加,这就有利于植物光合能力的提升,从而使植物在竞争中保持优势地位,而适应性不强的植物的光合速率会降低,光合能力会下降,面临着生态演替过程中被淘汰的风险。
温度升高增强了水葱和茭草光合能力的同时,也带来了不利影响。研究观察表明,光合增强使得水葱和茭草的株高和茎粗增加,但株高生长速度远远超过茎粗生长,且差异显著。水葱萌芽期株高极显著增加了12.05%和42.17%(P<0.01),生长期株高极显著增加了13.49%和44.76%(P<0.01),凋落期株高显著增加了7.72%和16.73%(P<0.05),茭草萌芽期、生长期和凋落期株高均极显著增加了19.1%和62.69%、27.3%和71.51%及8.48%和33.65%(P<0.01),而水葱3个时期的茎粗分别增加了2.89%和4.74%、4.18%和6.28%及2.76%和3.98%,茭草也分别增加了4.64%和14.99%、7.76%和18.06%及4.41%和9.57%,但它们的差异均不显著(P>0.05)。石冰[27]和刘国栋[28]对增温下水葱、茭草和芦苇的株高、茎粗的研究也得到同样结果。这种由于温度升高引起的茎粗生长和高生长差异,导致茎不够粗壮而发生倒伏,倒伏后植物过早衰亡,不利于种群的生息繁衍。此外,温度升高刺激光合增强,导致水葱和茭草的生物量积累增多,单位面积内的密度增加,植物的生长空间变小,极易发生病虫害,使生长周期提前结束,不利于种群的发展壮大(未发表数据)。
国内外有些研究者认为,温度升高提高了植物光合固碳的能力,使碳输入增加,但同时也使碳输出增加且碳输出大于碳输入。但对于水葱和茭草,温度升高是使其碳输入增加还是使其碳输出增加及它们的碳平衡关系如何?还需进一步深入研究,各气象因子之间互作对各项指标的影响,也还需进一步深入研究。
4 结论
海拔梯度下降的区域气候条件差异尤其是温度、太阳辐射和CO2浓度对湿地植物光合特性产生显著影响,而影响湿地植物生长的最主要的气象因子为温度。水葱和茭草的叶绿素含量、Fv/Fm、PhiPSⅡ、ETR和qP值均随着温度的升高而升高,NPQ值随着温度的升高而降低,表明水葱和茭草具有较强的环境适应能力,反映了作为广布种的宽适应特点[8]。这些值的变化使两者的光合速率提高,光合活性提升,光合能力增强。但茎粗生长和高生长不平衡的不利影响可能导致两种广布种最终难以适应温度上升而衰亡。
致谢:湿地学院/国家高原湿地研究中心的张昆老师对写作给予帮助,西南林业大学提供大型仪器设备平台,特此致谢。
