祁连山东部青杄年内径向生长动态对气候的响应
2018-11-15牛豪阁于爱灵张军周勾晓华
牛豪阁,张 芬,于爱灵,王 放,张军周,勾晓华
兰州大学资源环境学院,西部环境教育部重点实验室, 兰州 730000
树木年轮因其具有时间序列长、定年准确、分辨率高、复本量大、连续性强、地域分布广泛的特点[1],在我国西北地区的气候重建研究中有广泛应用[2- 5]。树轮-气候响应研究是树轮气候重建的基础[6],但是这一研究主要基于树轮宽度和气候要素的数理统计关系[7- 8],缺乏对树木生理基础的研究。而且在温带地区,雨季到来时间和生长季温度的变化会显著影响形成层活动,但是年际间的轮宽却仍可能保持不变[9-10],这导致基于轮宽的气候响应分析会覆盖生长季内各月的气候信息,不能揭示树木短期的径向生长变化与气候要素的关系。因此需要引入生长监测的手段,在更高分辨率的基础上(生长季内各月)分析树木径向生长对气候的响应特征,进而分析被监测树种的生存策略[11- 13]。
2010年以来,该地区对树木生长的监测逐步建立起来,并取得丰硕成果[14- 17]。但是,基于Dendrometer监测的树木径向生长包括细胞吸水膨胀失水收缩的可逆过程和细胞膨胀分裂的不可逆过程[18],且通过Dendrometer监测的是树干的直径和圆周变化而不是直接的木材形成,很难将真正的形成层细胞生长和由水分波动导致的膨胀收缩分离开[19]。Deslauriers等对意大利阿尔卑斯山东部不同海拔高度云杉和落叶松的监测研究显示,在低海拔地区(1020 m),利用10 d时间尺度的Dendrometer数据所确定的生长季与微树芯结果有很好的一致性[12];Köcher等通过径向生长监测的方法研究了5种阔叶树径向生长对环境要素的响应,发现在7 d、21 d时间尺度下提取的径向生长量与影响树木生长的主要因素—相对湿度的显著相关关系更加稳定[20]。这些研究说明通过延长时间尺度可以削弱细胞水分日波动的影响,更加准确地分析气候要素对树木径向生长的影响。且Li等[21-22]在藏东南色季拉山的监测研究发现,在一周的时间尺度上就可以监测到形成层细胞的径向生长活动;Mäkinen等也发现一周时间尺度的微树芯采样可以记录到木质部细胞的形成[23]。基于此,本研究将时间尺度延长到7 d、10 d,在尽量减少树干水分波动干扰的基础上,分析青杄径向生长与气候要素的关系。
为了避免监测时间较短导致树木径向生长对气候的响应结果不可靠,我们对吐鲁沟青杄进行了连续3年的监测(2013—2015),目的在于:(1)研究不同时间尺度下树木径向生长对气候要素的响应差异;(2)研究3年生长季内各月气候状况与树木径向生长的关系。
1 研究区概况
吐鲁沟国家森林公园(36°40′—36°44′N,102°36′—102°46′E;海拔1950—3300 m)位于兰州市永登县连城镇(图1),位于祁连山东部的大通河流域,属温带大陆性气候,青杄分布的西界。该地区的年日照时数为2655.2 h,年太阳总辐射为469 kJ/cm2,年平均气温7.4℃,无霜期125—135 d,年蒸发量为1542 mm,圈湾、苗圃年降水量分别为679 mm和657 mm,主要集中于4—9月, 7月降水最多[24]。
保护区内随着海拔上升,植被呈现明显的垂直分带特点:海拔2100 m以下主要分布山杨(Populusdavidiana)等阔叶树;随海拔升高依次为油松(Pinustabulaeformis)、青杄(Piceawilsorii)和青海云杉(Piceacrassifolia),在青海云杉林内分布有红桦(Betulaalbo-sinensis)、糙皮桦(BetulaBetulaceae),海拔3000 m以上乔木为祁连圆柏(Sabinaprzewalskii),灌木为杜鹃(Rhododendronspp.)、金露梅(Potentillafruticose)、鲜黄小檗(Berberisdiaphana)等等。青杄监测点位于吐鲁沟内的苗圃(36°41′32.57″N,102°44′22.92″E,海拔2145 m),坡度为20°,郁闭度在0.3左右,坡向为半阳坡。
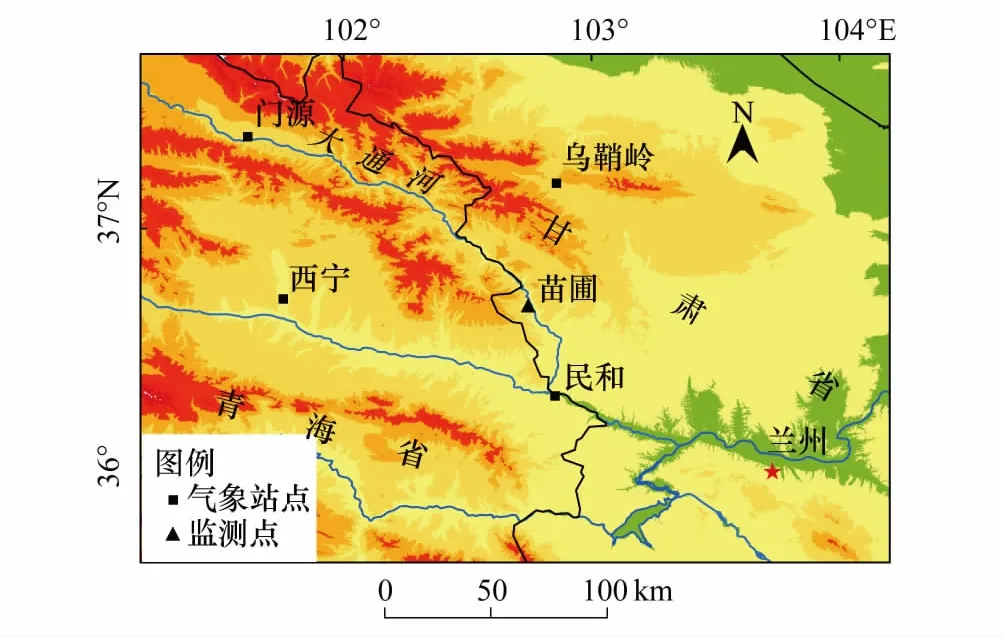
图1 吐鲁沟青杄监测点Fig.1 Monitoring point of Picea wilsorii in Tulugou
2 数据采集和处理方法
2.1 青杄径向生长数据采集处理
选取长势良好的青杄2棵(附近无径流、无工程干扰),两棵样木的树高为24 m左右,胸径分别为25.4、30.8 cm,树龄分别为80、94 a。在2013—2015年使用点触式树木径向生长监测仪(Point Dendrometer, Ecomatic Germany; 型号:DR; 精度:±2 μm)监测树木茎干的年内变化。首先,在树干胸径高度处剥去树皮,以便减少树皮水分变化对树干径向监测的干扰。然后,安装Dendrometer,并将仪器的记录时间间隔设置为30min,一天共记录48个数据[5]。Dowens根据树木径向的收缩和膨胀将树木的径向变化分为3个阶段:收缩阶段、恢复阶段和增长阶段,从上一个最大值到当前最小值的这段时间是收缩阶段,从当前最小值膨胀到上一个最大值的时间是恢复阶段,继续膨胀到下一个最大值的时间称为增长阶段[25]。Deslauriers将树木径向生长分为收缩阶段、膨胀阶段和径向增长阶段,该划分方法将恢复和增长阶段合并在膨胀阶段中[12]。根据这些阶段划分可以计算出树干一天的收缩量、膨胀量、增长量,进而提取树木的径向生长量。提取方法主要有3种:平均值法[26]、最大值法[27]、周期法[25]。Deslauriers对比发现3种方法提取的径向生长量有很好的一致性[12]。因此,本文使用平均值法提取青杄不同时间尺度(1 d、7 d、10 d)的径向生长量,即用下一个时间段径向生长量的平均值减去当前时间段径向生长量的平均值,正值表示存在绝对的径向生长量,负值表示没有径向生长,记为0。
Gompertz函数在树木径向生长拟合中有广泛的应用,可以用来确定树木的生长季[28-29]。将每天记录的48个数据求平均,得到青杄累积径向生长的日均值序列。通过t检验发现两棵树在3年生长季中的径向波动(P2013=0.491,P2014=0.068,P2015=0.056)和径向增长(P2013=0.136,P2014=0.098,P2015=0.162)没有显著的差异,所以将两棵青杄的数据平均,得到青杄的平均生长状况。使用Gompertz函数对青杄3个生长季每天的径向生长变化进行拟合,然后对拟合函数求导得到青杄在各年的生长速率,并将生长速率大于4 μm的时段作为青杄的生长季[5]。
Gompertz函数模拟公式如下式:
Y=Aexp[-e(β-kt)]
式中,Y为径向生长原始数据天均值;A为上渐近线;β表示与Y初始值有关的参数;k表示內秉生长率;t表示时间;将拟合方程求导,得到青杄的径向生长速率。
2.2 气象数据采集及处理
在距离树木径向生长监测点50 m的空地上安装自动气候站(YM-03),记录气候要素的变化特征,包括空气温度、相对湿度、降水量、太阳辐射、土壤温度及湿度、风速风向等;记录时间间隔为10 min,数据储存在数据采集器中。该监测点2013—2015年降水量分别为576.1、665.8 mm和641.8 mm,其中,6—9月的降水量分别占全年降水量的77%、64%和76%。2013、2015年平均最高气温出现在8月(19.31、18.17℃),2014年平均最高温在7月(18.38℃),2013年平均最低温出现在1月(-5.82℃),2014、2015年平均最低温在12月(-5.91℃、-4.03℃)(图2)。
本文计算了3个生长季内不同时间尺度(1 d、7 d、10 d)的平均气温(T)、平均最高温(Tmax)、平均最低温(Tmin)、降水量(P)、平均土壤湿度(SH)、土壤温度(ST)和饱和水汽压亏缺(VPD),用于分析不同的时间尺度气候要素与青杄径向生长的关系。为了分析各气候要素对青杄径向生长的贡献,本文在一天的时间尺度上对二者做了逐步回归分析;同时以31天为窗口计算它们的滑动相关系数,分析青杄年内径向生长变化与气候要素关系随时间的变化。
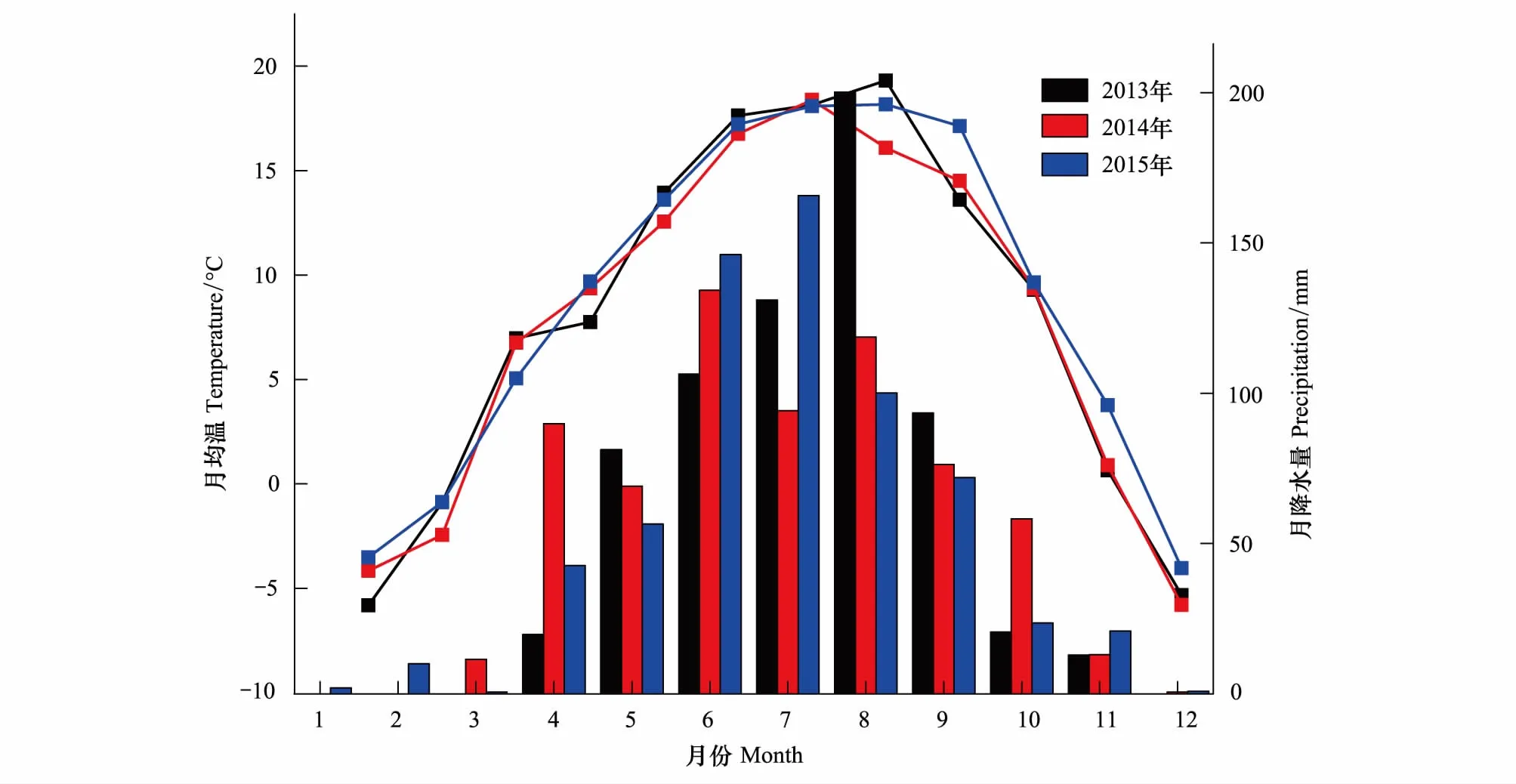
图2 吐鲁沟气象站2013—2015的气温、降水变化Fig.2 Climate diagram for the Tulugou meteorological station during 2013—2015
3 结果
3.1 青杄径向生长模拟
通过Gompertz函数对两棵青杄3年的平均生长序列进行拟合(图 3),3年径向生长的拟合方程R2均达到99%,表明方程很好的拟合了青杄的径向生长变化。结果显示青杄的最大径向生长速率集中在5月底6月初,2013到2015年的最大生长速率分别在5月26日、6月6日和6月6日;2013年生长开始和结束的时间分别是4月20日和8月6日,2014年分别是4月30日和8月21日,2015年分别是4月27日和8月24日。2013年青杄拟合一开始生长速率就超过了4 μm/d,可能是受到春季复水的影响,即径向生长增加是由细胞吸水膨胀导致,并非有新的形成层细胞生成[23,30]。
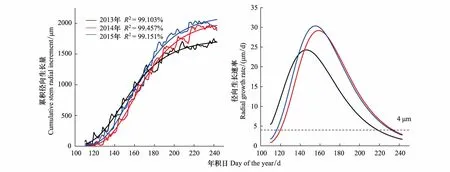
图3 Gompertz函数模拟的青杄3年径向生长变化及生长速率Fig.3 Gompertz function modeled curves of cumulative radial variation and radial growth rate from 2013 to 2015左图细折线表示原始数据的累积径向生长量,粗曲线表示拟合的累积径向生长量
3.2 径向生长量与气候要素的关系
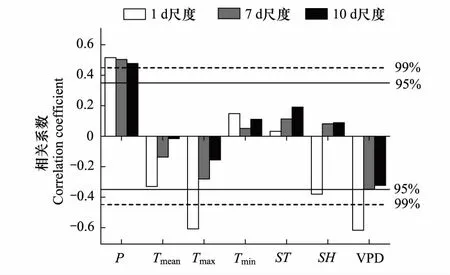
图4 青杄3年生长季内1d、7d和10d径向生长量与气候要素的相关分析Fig.4 Correlation coefficients between stem radial increment of different time scales1d、7d和10d对应的自由度分别为335、45和32,根据自由度32设定置信线;P, Tmean, Tmax, Tmin, ST, SH, VPD分别表示降水量,平均温,最高温,最低温,土壤温度,土壤湿度,饱和水汽压亏缺
将3年生长季内青杄的日径向生长量与气候要素做相关分析,在1 d尺度上,与降水显著正相关,与最高气温和VPD显著负相关。在7 d和10 d时间尺度上,青杄的径向生长变化对气候要素的响应模式与1 d尺度一致,但是相关系数有所降低(图4)。
利用逐步回归分析建立回归模型,分析各气候要素对青杄径向生长的贡献,也显示降水对青杄的径向生长有主要积极作用,温度尤其是最高温对其径向生长限制明显,方程如下:
YSRI=81.873+1.682P-2.026Tmax-0.154VPD
回归方程中的所有参数都通过0.05显著性水平检验,具体方程统计信息如表1。

表1 回归方程统计信息
为了更加直观地分析青杄径向生长与气候要素的关系,本文将2013到2015年3年生长季内的日径向生长量与对应时段的气温和降水进行了对比(图5),发现高温少雨的天气条件对应低的径向生长量,低温多雨对应高径向生长量,充沛的降水有利于青杄的径向生长。
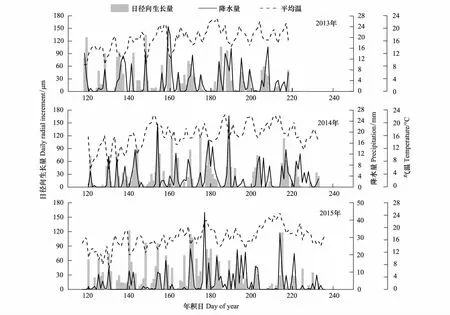
图5 2013—2015年生长季内日径向生长量与气温、降水量对比Fig.5 The comparison between daily stem radial increment and climate factors (temperature, precipitation) in growing season from 2013 to 2015
以上结果显示不同时间尺度的径向生长都与降水有稳定的正相关关系,为了分析青杄年内径向生长与气候要素关系随时间的变化,本文以31 d为窗口,每隔一天进行滑动,计算了二者在一天时间尺度上的滑动相关系数(图6)。3年生长季内各月径向生长与气温、饱和水汽压亏缺(VPD)都为负相关,与降水都为正相关,但是相关系数随时间的推移波动较大,年际间也有很大差异。3年的径向生长都与5月、7月降水显著正相关,与6月降水的相关性波动较大,2013和2014年径向生长与6月降水显著正相关,2015年相关系数急剧降低;31天滑动降水量显示2015年6、7月的降水比2013、2014年明显要多。且6月青杄的径向生长速率较快(图3),所以6月气温降水的波动会对其径向生长产生重要影响。
4 讨论
4.1 青杄不同时间尺度径向生长量与气候要素的响应关系
不同的时间尺度上,青杄的径向生长量都与降水显著正相关,与温度、VPD负相关,说明该地区水分蒸散严重,其径向生长受到了水分的限制[1]。但是树木的径向生长包括细胞吸水膨胀、失水收缩的可逆过程和细胞膨胀分裂的不可逆过程[20,31],在1 d时间尺度上,细胞从傍晚开始吸水膨胀,持续到早上膨胀结束,白天蒸腾失水收缩[32],这种细胞失水复水过程会记录水分变化信号,在短时间尺度上,该信号可能会叠加到细胞生长膨胀分裂所揭露的水分变化信号上,所以在1 d的时间尺度上,径向生长量与水分的相关性较高(图4)。
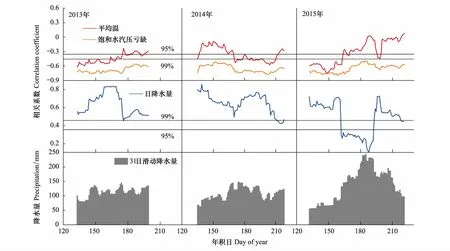
图6 2013—2015年青杄生长季内日径向生长量与饱和水汽压亏缺、温度和降水31天滑动相关Fig.6 31-day moving correlation between the daily stem radius change and the daily climate factors红色折线表示天均VPD与径向生长量的31天滑动相关;黑色折线表示日均温与径向生长量的31天滑动相关;蓝色折线表示日降水量与径向生长量的31天滑动相关;蓝色柱状图表示31天降水量的滑动
为了减弱细胞失水复水对径向生长所记录的气候信号的干扰,本文将径向生长量的时间尺度延长到7 d和10 d,结果与降水的相关度有所降低,说明通过延长时间尺度可以在一定程度上减弱细胞失水复水的干扰。但相关系数仍在显著度水平以上,证实了该地区树木的径向生长确实受到了水分限制。充沛的降水有利于细胞膨胀增大和分裂,从而促进树木的生长[33-34]。相反,如果极度缺水,会造成细胞膨压减小,影响细胞的新陈代谢[35],进而抑制树木的径向生长。
4.2 青杄年内径向生长对气候要素的响应
虽然上文已经明确研究区青杄径向生长主要受水分限制,但是由于不同月份径向生长量和水分条件不同,因此青杄径向生长与降水和温度的关系存在很大的年内差异,在一天时间尺度上,以31 d为窗口,每隔一天进行滑动相关分析,发现3年的径向生长与5、7月降水显著正相关,与6月降水的相关性存在年际差异,2015年6月的径向生长与降水相关不显著(图6)。
5月青杄径向生长处于加速阶段,但雨季还未到来,并且温度缓慢回升,树木径向生长受到水分的限制[36]。7月降水较多,然而空气温度较高,树木蒸腾旺盛,消耗大量的水分,可能对树木生长造成干旱胁迫。并且长期的野外监测发现该地区7月有很多集中在下午的短时降水,7月温度较高,短时降水后天气转晴,蒸发迅速,导致水分的利用效率较低。对7月下午的短时降水天数和降水量进行了统计,并将全天都有降水和连续几天都有降水的情况排除在外,结果显示(表2):3年7月下午的短时降水天数和降水量都占很大比例,尤其是2015年不仅下午短时降水的天数超过50%,其绝对降水量也超过一半。所以即便7月的降水较多,但是高温和这种短时降水集中的情况使水分蒸散严重,很大程度上降低了树木对降水的利用效率,导致该时间段的水分仍然是树木径向生长的制约因子。
青杄3年的径向生长与降水的关系在5、7月保持一致,差异集中在6月,并且径向生长模拟的结果显示青杄的最大径向生长率集中在5月底6月初,其他地区的研究也发现树木径向生长速率最大在春季末期[12, 37],所以该时段的气候变化对青杄的径向生长有重要影响。本文通过3年的监测研究发现6月降水与径向生长的关系存在明显的年际差异,二者关系在2015年不显著,但是在2013和2014年显著正相关,研究认为这与2013和2014年6月降水偏少有关:通过3年6月份降水量的对比分析发现,2015年6月的降水量最大,可以达到146.1 mm,2013和2014年6月降水相对较少,分别是106.4、134.3 mm,一定程度上限制了青杄的径向生长。

表2 2013—2015年7月下午短时降水天数及降水量在整个月中的比例
2013—2015三年6月降水的年际变率较大,导致树木可利用水分变幅也较大,因而树木径向生长与6月水分的响应关系不稳定,这种不稳定的现象可能会导致一些树轮-气候响应研究中轮宽与6月降水的关系在多年统计上不显著。方克艳等研究了兴隆山(距离吐鲁沟将近300 km)和吐鲁沟青杄的树轮气候响应,结果显示青杄的径向生长受到生长季水分的限制,但是都与生长季内6月的降水关系不显著[38-39];且不同地区的监测结果也有很大差异,张瑞波等对天山地区雪岭云杉一年(2014年)的径向生长监测显示,5月底到6月底的水分状况是其径向生长的主要限制因素[40];但是贺敏慧等在祁连山地区对祁连圆柏的监测发现其径向生长主要与5、7月降水显著正相关,6月份的降水并不是其径向生长的限制因子[36]。刘文火统计了我国西北东部地区树木径向生长对气候的响应情况,发现该地区年表基本上都与各月降水正相关,但是与上一年9月和当年5月降水显著正相关的年表比例最高,分别为73.5%和50.5%,与6月降水显著相关的比例较低(20%)[41]。所以树轮对6月降水响应不稳定的现象,可能在更大范围、不同树种间也同样存在,这有赖于我们在更大范围内对不同的树种进行持续监测。
5 结论与展望
本文在分析青杄日径向生长量与气候要素的基础上,又将时间尺度延长到7 d、10 d,目的在于减弱日径向变化中水分的干扰,不同时间尺度上的分析都反映了水分对青杄径向生长的限制,随着时间尺度的延长,相关系数有所降低,说明延长时间尺度可以在一定程度上避免水分日间波动信号叠加到细胞生长膨胀分裂所揭露的水分变化信号上,从而保证响应分析的准确性。逐步回归分析的结果也显示水分在青杄径向生长中的积极贡献最大,同时为了分析生长季内各月气候要素与青杄径向生长关系,我们又以31 d为窗口在一天时间尺度上进行了滑动相关分析,结果显示:3年生长季5到7月的降水都与青杄的径向生长起显著正相关,关系稳定,但是6月降水对青杄径向生长的影响存在较大的年际波动,在降水量大的年份相关性不明显,在降水量少的年份相关性显著,这可能会导致一些树轮-气候响应研究中轮宽与6月降水的关系在多年统计上不显著。
此外,微树芯法可以更加直接的监测到树木的径向生长,能够精确确定树木的生长季,在很大程度上避免了外界水分的干扰,接下来需要与微树芯结合,进一步研究树木径向生长与水分的关系。
致谢:感谢甘肃省连城国家级自然保护区管理局华发春局长、满自红科长在野外监测过程中给予的帮助,特此致谢。
