不同种植方式下两种草本营养元素对土壤厚度和水分减少的响应
2018-11-14赵雅洁宋海燕梁千慧李素慧刘锦春
赵雅洁,李 周,宋海燕,张 静,梁千慧,李素慧,刘锦春
三峡库区生态环境教育部重点实验室,重庆市三峡库区植物生态与资源重点实验室,西南大学生命科学学院, 重庆 400715
碳(C)是植物各种生理生化过程的底物和能量来源,植物主要通过光合作用同化空气中的CO2获取生长所需的C;而氮(N)和磷(P)是蛋白质和核酸等生物分子的重要组成元素,植物主要通过根系从土壤中吸收生长所需的N和P[1- 2]。3种元素作为植物生长发育过程中所必需的营养元素和细胞结构物质中重要的组成元素,在植物的生长和生理生化调节机能中发挥着重要的作用[3- 4]。研究表明,植物可以通过调节C、N、P的代谢和循环,改变元素的吸收、积累和分配来响应和适应环境的变化[1,5]。如在干旱环境下,由于植物吸收和运输养分能力降低,植物N和P含量和积累量显著降低,植物体内营养元素在不同器官中的分配也发生变化[6- 8];而土壤浅薄或土壤厚度的减少使得土壤中所含的养分总量降低,同时浅薄土壤保水能力差,水分蒸发更快,可能进一步加重干旱胁迫,因而可能也会导致植物对营养元素的积累量降低,元素分配比发生改变[9- 10]。
在我国西南喀斯特地区,岩溶干旱和“土层厚薄不均”是限制植物生长的两个关键生态因子[11- 12]。面对水分和土壤资源的缺乏及分布不均,喀斯特地区的植物具有较强的形态可塑性和生理调节能力,最大程度地减少不利环境因素的影响,提高自身的适应能力和竞争力[13- 15]。例如,喀斯特地区的适生物种通常具有较厚的角质层或蜡质层,发达的表皮毛和根系,较高的根冠比和水分利用效率以及较高的渗透调节能力和抗氧化保护能力[14- 15]。但另一方面,为了更大程度地利用有限的资源,喀斯特地区的植物可能存在一定的生态位分化,通过生态位互补减少资源缺乏对共存物种的抑制作用[16- 18]。我们的前期研究表明,在喀斯特土壤上混种深根植物苇状羊茅和浅根植物黑麦草,确实发现两者的总生物量和地上生物量并没有受到影响,也就是说两者之间的竞争效应不明显[11];同时,两者的光合作用受资源减少的抑制作用也均得到缓解[12]。然而,土壤和水分双重资源的减少还是加剧了苇状羊茅和黑麦草的地下竞争,但两者能够通过不同的生长策略(根系生长、生物量分配)和生理策略(光合调节、水分利用)来适应不利环境。
目前,国内外学者开展了大量关于喀斯特植物营养元素对干旱生境的适应性研究,但有关土壤资源的匮乏对该地区植物营养元素特征的影响较少,而土层浅薄和岩溶干旱双重胁迫共同对该地区植物营养元素的影响研究更少。因此,在以“岩溶干旱”和“土层浅薄化”为特征的喀斯特生境下,深根植物苇状羊茅(FestucaarundinaceaSchreb.)和浅根植物黑麦草(LoliumperenneL.)的营养元素将受到怎样的影响?两者混种后分别采取何种元素调节策略提高各自的适应和竞争能力?因此,本研究在前期研究的基础上,继续以深根植物苇状羊茅和浅根植物黑麦草为研究对象,通过模拟控制实验,探讨:1)土壤厚度和/或水分减少对苇状羊茅和黑麦草叶片、根系C、N和P元素含量、积累和分配特征的影响;2)两者混种后营养元素的积累和分配策略。以期更深入而全面地了解喀斯特地区植物对环境的适应,从而为该地区的植被恢复和重建提供参考。
1 材料与方法
1.1 试验设计
本试验选择深根植物苇状羊茅和浅根植物黑麦草为研究对象。供试土壤是取自重庆市沙坪坝区海石公园黄色石灰土,基本理化性状:pH为7.4,有机质为0.34%,全氮为0.28 g/kg,全磷为0.39 g/kg,全钾为23.7 g/kg,田间持水量为39.8%[12]。试验采用土壤厚度、水分和种植方式三因素完全随机区组设计。土壤厚度(T)包括对照土壤厚度(15 cm,TCK)和浅土(5 cm,TS),以自制的两种相同底面积(100 cm2)不同深度的长方体栽培容器实现,分别依次装入1500 g和500 g的干土,以上两种土壤厚度是基于野外实地考察和前期预实验而设计的。于2015年4月4日选取长势和大小基本一致的幼苗(株高为20 cm左右)移栽到容器中,放置在西南大学生态园透明雨棚下(29°49′N,106°30′E,海拔249 m)。种植方式(PP)分为单种和混种,每盆2株。
待幼苗全部存活并适应生长一段时间后,于2015年4月14日进行水分处理。根据重庆地区1981—2011这30年间4—6月降雨量和本试验具体的容器规格计算出日平均降雨量40 mL,以40 mL作为对照水分处理(120 mL/3天,WCK)。基于预实验,本研究以在对照的水平上减少70%(12 mL)的水分处理作为干旱处理(36 mL/3天,WD),此时水分胁迫为中度干旱胁迫[19]。水分处理历时69 d,每3天浇一次水(共浇水22次),每个处理5个重复。
同时,每种土壤厚度和水分处理设置3个无植物生长的空白处理,进行同步的水分处理,用于测定土壤含水量。每次水分处理前,通过五点取样的方法进行土样的采集,使用称重法进行土壤含水量的测定,其中对照组(CK:TCKWCK)和浅土组(S:TSWCK)的土壤含水量为21.3%和20.3%,均属于偏湿生境,干旱组(D:TCKWD)和浅土+干旱组(SD:TSWD)的土壤含水量分别为9.1%和7.3%,均属于中旱生境[19]。
1.2 指标测定
于2015年6月22日,对地上部分和地下部分进行收获,将植物地上部分和根系于80℃烘箱中烘干后,用球磨仪进行研磨,干燥箱中备用。同时每一处理下选取5株植株,测量植株的生长状况,用直尺测量株高,用数字化扫描仪(STD1600Epson USA)和WinRhizo根系分析系统(Regent Instrument Inc.,Canada)对叶面积和总根长进行测定。
(1)C和N元素:称取0.005 g样品粉末,用锡舟包好,使用元素分析仪(Vario EL cube,Elementar,GER)测定C和N百分比含量,经过换算得出C、N元素含量(mg/g)。
(2)P元素:称取0.05 g样品粉末于消解罐中,并加入2 mL双氧水和6 mL硝酸,用微波消解仪(SeepWave MWS- 4,Berghof,GER)消解后,转移并定容到50 mL离心管中,再用iCAP6300电感耦合等离子体发射光谱仪(ICP-OES,Thermo Fisher,UK)测定出P元素浓度(mg/L),经换算得P元素含量为(mg/g)。
(3)器官营养元素积累量=各器官营养元素含量×各器官对应的生物量;根系营养元素分配比=根系营养元素积累量/(地上部分营养元素积累量+根系营养元素积累量)。
还有一种,比如说同时期的不同题材的作品来进行对比。就拿郁达夫在1927年写的东西作例子。郁达夫在1927年有一个很奇怪的现象,就是他的论文和杂文都是慷慨激昂的,可以举例如《在方向转换的途中》《公开状答日本山口君》《无产阶级专政和无产阶级文学》《诉诸日本无产阶级文艺界同志》。从这四篇论文或杂感来看,他是慷慨激昂的。甚至有一篇题目叫《无产阶级专政和无产阶级文学》,1927年写这样的文章也就很慷慨激昂了,但是在1927年他的小说比如《过去》《迷羊》《清冷的午后》,从这些小说来讲,都是很失望的,甚至是绝望的。那么这样一对比,将这些资料一排比,我觉得也能够得出论点。
1.3 统计处理
利用双因素方差分析(Two-way ANOVA)揭示土壤厚度和种植方式、水分和种植方式、土壤厚度+水分和种植方式对两物种C、N、P元素含量、积累及分配的影响。采用独立样本T检验(Independent-samplesTtest)分析相同土壤厚度和水分处理下同一物种各个指标在不同种植方式下的差异显著性。本实验中的数据分析均利用SPSS 22.0软件完成。采用Origin 8.6软件制图。
2 结果与分析
2.1 生长状况
无论单种还是混种,与对照组相比(CK),苇状羊茅和黑麦草的株高、叶面积、总根长在浅土(S)、干旱(D)和浅土+干旱(SD)处理下均不同程度的降低(表1)。
在各资源水平下(CK、S、D、SD),两物种的株高和叶面积在单种和混种下无显著差异。混种后苇状羊茅的总根长在对照水平下(CK)显著高于单种,而在3种低资源水平下(S、D和SD)显著低于或与单种无显著差异。黑麦草的总根长在各资源水平下在单种和混种间均无显著差异(表1)。
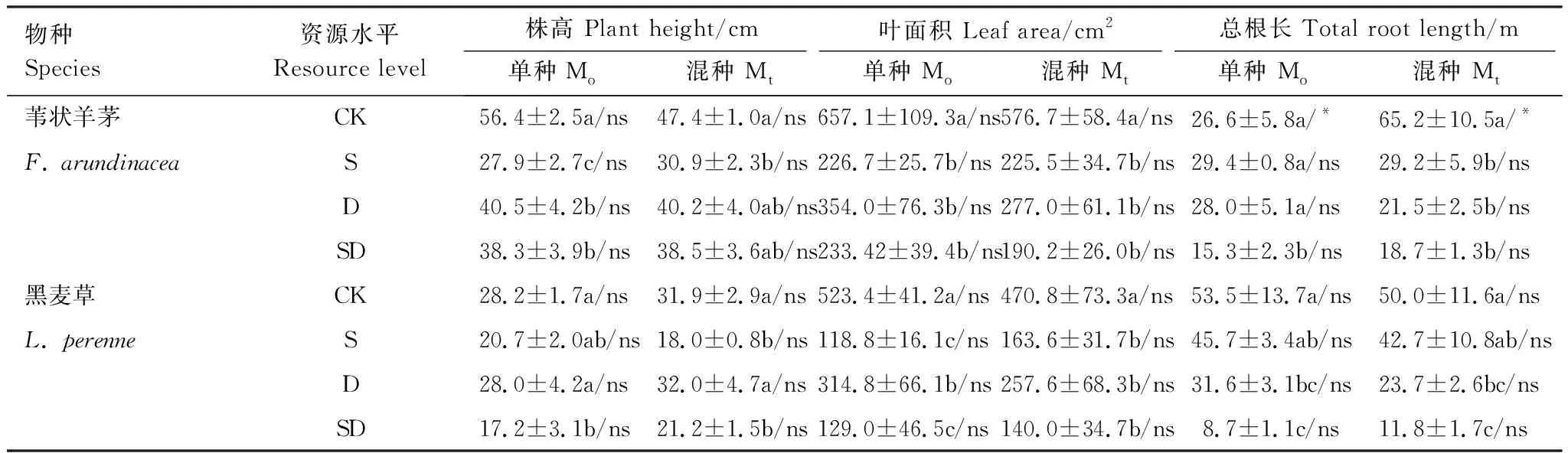
表1 苇状羊茅和黑麦草植株的生长状况
CK:对照组,Control;S:浅土组,Shallow soil group;D:干旱组,Drought group;SD:浅土+干旱组,Shallow soil + drought group;Mo:单种,Monoculture;Mt:混种,Mixture;不同小写字母表示同一器官下不同资源水平间差异显著(P<0.05);ns/*,同一资源水平下单种和混种间差异显著性(ns,P>0.1; *,P<0.05)
2.2 营养元素含量
2.2.1 C含量
无论是单种还是混种,与对照相比,苇状羊茅的地上部分C含量在3种低资源水平下(S、D、SD)显著增加,而根系C含量无显著变化。在对照资源水平下(CK),混种后苇状羊茅的地上部分C含量比单种显著增加6.66%,而根系C含量与单种无显著差异。而在3种低资源水平下(S、D和SD),混种后苇状羊茅的地上部分和根系C含量仅在干旱组显著高于单种(图1)。
在单种和混种条件下,黑麦草的地上部分C含量在3种低资源水平下显著低于对照组,而根系C含量与对照组无显著差异。在对照资源水平下,混种后黑麦草的地上C含量与单种无显著差异,而根系C含量显著高于单种,增加的幅度为2.14%。在低资源水平下,黑麦草的地上部分和根系C含量在单种和混种之间无显著差异(图1)。
两因素方差分析表明,苇状羊茅地上部分C含量仅在T和PP的交互作用下有显著差异,根系C含量仅在W和PP的交互作用下有显著差异。黑麦草的地上部分和根系C含量在T和PP、W和PP、T+W和PP的交互作用下均无显著差异(表2)。

图1 土壤厚度、水分及土壤厚度+水分对不同种植方式下苇状羊茅和黑麦草C含量的影响(平均值±标准误)Fig.1 Effect of soil thickness, water and soil thickness plus water on C content of F. arundinacea and L. perenne under different planting patterns(M±SE)柱上的标注表示同一物种在相同资源水平下单种和混种间的差异;**(P﹤0. 01)和***(P﹤0.001)极显著水平;*显著水平(P﹤0.05);+(0.05﹤P﹤0.1);ns未达到显著水平(P﹥0.1)
2.2.2 N含量
无论单种还是混种,与对照相比,在浅土处理下苇状羊茅和黑麦草的地上部分和根系N含量显著降低,在干旱和浅土+干旱处理下显著增加,且在各资源水平下,单种和混种之间均无显著差异(图2)。两因素方差分析表明,苇状羊茅和黑麦草的地上部分和根系N含量在T和PP、W和PP、T+W和PP的交互作用下均无显著差异(表2)。
2.2.3 P含量
无论是单种还是混种,在3种低资源水平下,苇状羊茅的地上部分和根系P含量相比于对照组显著降低。混种后苇状羊茅的地上部分P含量在各资源水平下与单种均无显著差异。苇状羊茅的根系P含量在对照资源水平下比单种显著增加30.29%,而在浅土、干旱和浅土+干旱3种处理中分别比单种显著增加70.02%、72.04%和24.81%(图3)。
表2苇状羊茅和黑麦草营养元素含量和分配的双因素方差分析
Table2ResultsofTwo-wayANOVAtestfortheeffectsofsoilthicknessandplantingpattern,waterandplantingpattern,soilthicknesspluswaterandplantingpatternonthenutrientcontentanddistributionratioofF.arundinaceaandL.perenne

物种 Species方差来源 Source of variation分子自由度ndfF营养元素含量 Nutrient content地上部分Aboveground part根系 Root根系营养元素的分配比 Nutrient distribution ratio of root/%CNPCNPCNP苇状羊茅F. arundinacea土壤厚度 Soil thickness (T)18.95**29.52***20.21***0.67ns21.21***4.78*14.03**26.02***16.69***种植方式 Planting pattern (PP)17.78*0.08ns0.27ns0.00ns0.64ns21.31***0.08ns0.05ns2.78ns土壤厚度×种植方式 T×PP15.46*4.24+0.02ns2.81ns2.10ns0.01ns8.63**14.86**1.26ns水分 Water (W)133.65***14.37**12.77**3.88+3.67+25.10***0.03ns0.07ns0.03ns种植方式 PP115.75***0.04ns0.19ns1.63ns0.36ns21.09***0.00ns0.00ns2.81ns水分×种植方式 W×PP10.97ns4.14+0.02ns9.57**0.00ns0.48ns5.97*7.22*3.56+土壤厚度+水分 T+W118.59***23.81***17.22***0.18ns7.93*1.62ns0.43ns1.26ns0.31ns种植方式 PP18.55**0.65ns0.00ns1.63ns1.24ns12.17**0.37ns0.58ns3.18+(土壤厚度+水分)×种植方式 (T+W) × PP14.02+1.68ns0.13ns0.59ns0.20ns0.91ns2.61ns4.33+4.45+黑麦草土壤厚度 T15.23*14.35**7.10*1.61ns10.15**12.50**29.44***28.64***20.97*** L. perenne种植方式PP10.42ns0.37ns0.00ns11.96**0.12ns0.04ns4.40+5.52*8.55**土壤厚度×种植方式 T × PP11.00ns2.82ns1.17ns0.75ns3.59+1.25ns9.10**7.99*10.77**水分 W110.38**23.28***4.42+1.01ns5.90*0.78ns6.42*10.16**0.64ns种植方式 PP10.03ns0.14ns0.00ns7.42*2.77ns0.20ns0.01ns0.03ns0.24ns水分×种植方式 W× PP10.00ns1.48ns0.75ns1.15ns0.02ns0.35ns0.78ns1.00ns0.85ns土壤厚度+水分T+W10.86ns23.16***1.53ns0.62ns13.39**51.83***3.27+2.18ns0.01ns种植方式 PP11.39ns0.36ns1.26ns11.70**0.48ns1.16ns1.88ns1.65ns0.11ns(土壤厚度+水分)×种植方式 (T+W) × PP11.04ns0.00ns0.02ns1.13ns0.26ns0.15ns0.56ns0.63ns0.00ns分母自由度 ddf181818181818181818
ns未达到显著水平(P﹥0.1);+(0.05﹤P﹤0.1);* 显著水平(P﹤0.05);**(P﹤0. 01)和***(P﹤0.001)极显著水平

图2 土壤厚度、水分及土壤厚度+水分对不同种植方式下苇状羊茅和黑麦草N含量的影响(平均值±标准误)Fig.2 Effect of soil thickness, water and soil thickness plus water on N content of F. arundinacea and L. perenne under different planting patterns (M±SE)
对于黑麦草而言,在单种和混种条件下,其地上部分和根系P含量在3种低资源水平下与对照相比均显著的降低,且在各资源水平下,混种后苇状羊茅的地上部分和根系P含量与单种无显著差异(图3)。
两因素方差分析表明,苇状羊茅和黑麦草的地上部分和根系P含量在T和PP、W和PP、T+W和PP的交互作用下均无显著差异(表2)。
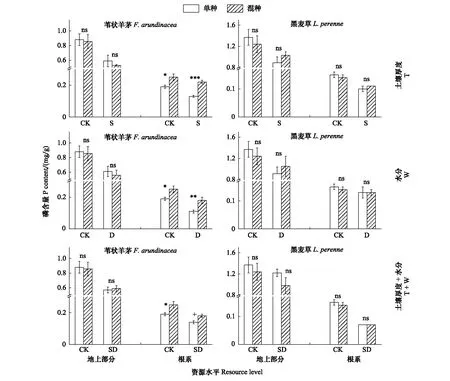
图3 土壤厚度、水分及土壤厚度+水分对不同种植方式下苇状羊茅和黑麦草P含量的影响(平均值±标准误)Fig.3 Effect of soil thickness, water and soil thickness plus water on P content of F. arundinacea and L. perenne under different planting patterns (M±SE)
2.3 营养元素积累量
无论是单种还是混种,总体上来说,苇状羊茅的地上、根和总C、N和P积累量在3种低资源水平下相比于对照显著降低或不变。在对照资源水平下,混种后苇状羊茅的根系C、N和P积累量分别比单种显著增加117.02%,100.88%和174.01%,其余情况均无显著差异(图4,仅以P积累量为例)。
对于黑麦草而言,在3种低资源水平下,其地上、根和总C、N和P积累量在单种和混种下与对照相比显著降低或不变,且在各资源水平下,单种和混种间均无显著差异(图4,仅以P积累量为例)。
两因素方差分析表明,苇状羊茅仅根系C、N和P积累量在T和PP、W和PP、T+W和PP的交互作用下有显著差异,黑麦草仅地上部分和总N和P积累量在T和PP的交互作用下有或接近有显著差异(表3)。
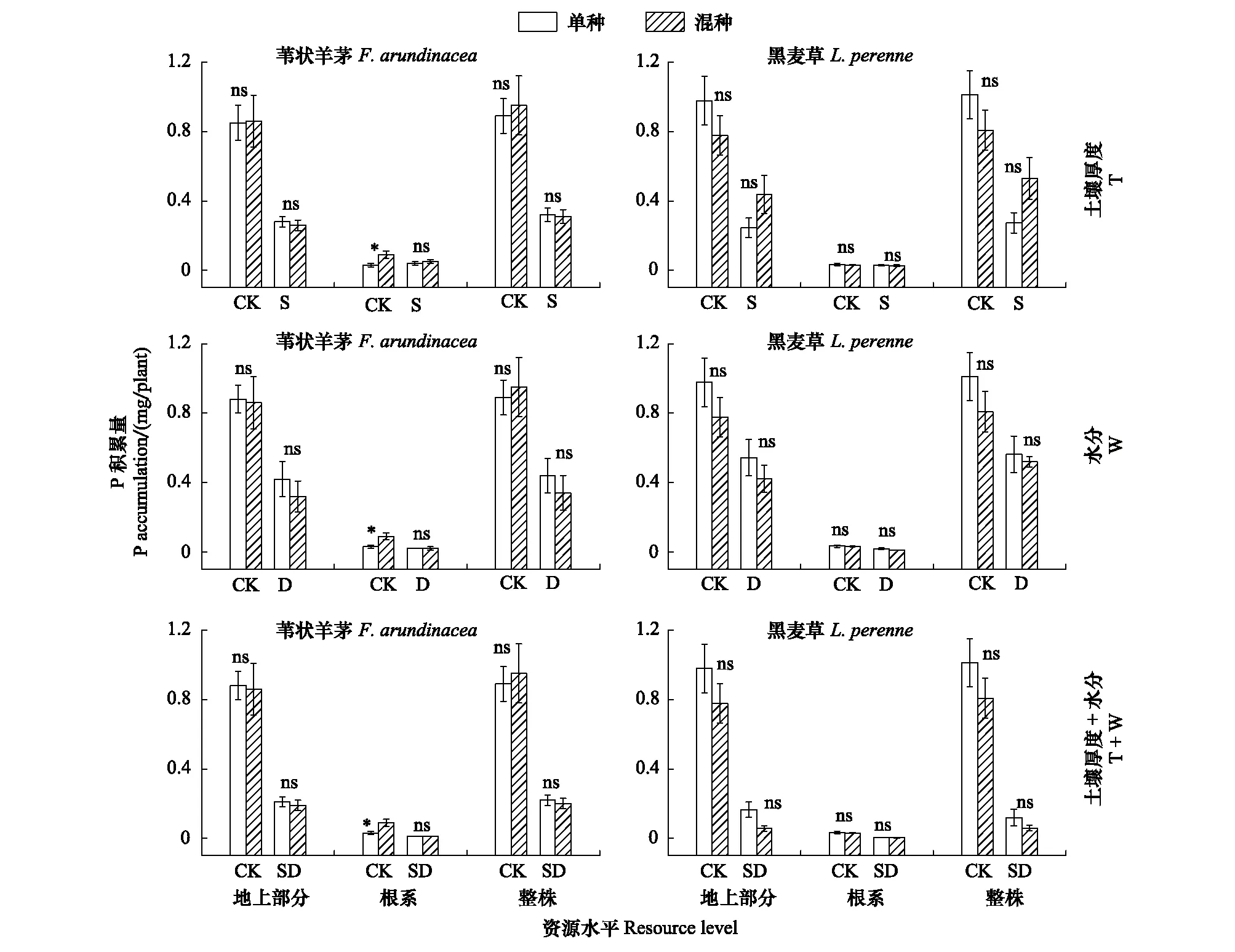
图4 土壤厚度、水分及土壤厚度+水分对对不同种植方式下苇状羊茅和黑麦草P积累量的影响(平均值±标准误)Fig.4 Effect of soil thickness, water and soil thickness plus water on P accumulation of F. arundinacea and L. perenne under different planting patterns (M±SE)
2.4 营养元素分配-根系营养元素分配比
与对照相比,在浅土组,苇状羊茅的根系C、N和P元素分配比在单种和混种下呈增加的趋势;而在干旱组和浅土+干旱组,苇状羊茅的根系C、N和P元素分配比在单种时增加,而混种时转为降低。在对照资源水平下,混种后苇状羊茅的根系C、N和P元素分配比在混种时分别比单种显著增加53.39%,98.21%和147.55%。而在3种低资源水平下,仅在浅土处理下,混种后苇状羊茅的根系C和N元素分配比显著或接近显著低于单种,降低的幅度分别为27.56%和31.04%(图5)。
无论是单种还是混种,与对照组相比,黑麦草的根系C、N和P元素分配比在浅土组显著降低,在干旱组显著增加,而在浅土+干旱组无显著变化。混种后黑麦草的根系C、N和P元素分配比在浅土处理下分别比单种显著降低31.54%,40.21%和51.93%,其余情况下单混种间均没有显著差异(图5)。
两因素方差分析结果表明,苇状羊茅仅根系C、N和P积累量在T和PP、W和PP、T+W和PP的交互作用下有显著差异,黑麦草仅地上部分和总N和P积累量在T和PP的交互作用下有或接近有显著差异(表2)。
3 讨论
3.1 两物种的营养元素对土壤厚度和水分减少的响应
C、N和P在植物生长发育过程中发挥着重要的作用,结构性元素C与功能性元素N、P相互作用共同调控植物生长[20]。植物C、N和P含量能够反应植物对干旱胁迫等不利环境的适应和防御策略[21- 22]。C含量越高,植物对外界不利环境的防御能力越强,而N和P含量越高,植物的资源获取和利用效率越高[21,23- 24]。整体上来说,在浅土组、干旱组和浅土+干旱组,苇状羊茅和黑麦草的地上部分C含量相比于对照组显著增加。这表明在资源缺乏或干旱情况下,两物种地上部分通过光合作用同化空气中的CO2,合成大量富含C 的有机化合物(如木质素或纤维素)[6,25],使得两物种茎叶中的C含量增加,以增强自身防御能力来应对土壤厚度和水分资源的减少[21]。在浅土处理下,苇状羊茅和黑麦草的地上部分和根系N和P含量较对照显著降低,因为土壤厚度的减少导致土壤中所含的营养元素总量降低,因而植物根系对N和P的吸收减少,运输至地上部分元素也减少。在干旱和浅土+干旱处理下(均属于中旱生境[19]),两物种地上部分和根系的N含量显著增加。研究认为,植物的抗旱性与N元素有关,在干旱或半干旱地区选择更高效吸收N途径的植物可能是一种很好的策略[26]。Sánchezrodríguez等[27]也发现,在中度干旱胁迫下,抗旱型品种cv. Zarina的N含量提高,其他品种的N含量均降低。可见,苇状羊茅和黑麦草在中旱生境下对资源的获取和利用效率较高,抗旱性相对较强,可优先考虑用于喀斯特地区植被恢复的工作中。相反,干旱和浅土+干旱处理下,苇状羊茅和黑麦草P含量与对照相比显著降低。Du等[28]提出,在西南喀斯特生态系统中,N/P<10为N限制,N/P>18为P限制。本研究中,在干旱和浅土+干旱处理下,苇状羊茅和黑麦草的N/P均大于18,这表明两物种的生长确实受到P的限制,干旱胁迫又更加抑制土壤P的扩散以及植物对P的吸收和运输,使得两物种对P的吸收速率小于相对生长速率(RGR),导致自身体内的P元素降低[7,28- 29]。董顺德等[30]也发现干旱胁迫使得烤烟体内的N含量增加,而P含量降低。
植物营养元素的积累量主要是由植株各组分的生物量和养分含量决定,是植物生长潜力的重要指标[31]。本研究中,在浅土组和干旱组,苇状羊茅和黑麦草的地上部分、根系和整株的C、N和P积累量在单种和混种下较对照均显著降低,且在土壤厚度+水分双重资源减少下降低幅度比其单种资源减少高2倍左右,这表明土壤厚度和水分资源的减少抑制了两物种元素积累量,且双重胁迫比两者单独胁迫抑制作用更强。尹丽[8]和王晓君[32]也发现,干旱胁迫降低了植株各器官中营养元素的积累量。
根茎叶中营养元素百分比反应了某元素在植物体内各器官间的迁移规律[33]。与对照相比,在3种低资源水平下,苇状羊茅的根系C、N和P元素分配比在单种时呈增加的趋势,而混种时转为降低(浅土除外)。这表明在低资源水平下,苇状羊茅在单种时趋向于将更多营养元素分配到根系中,以促进根系的生长来获取更多的水分和养分来抵御干旱胁迫,而在混种时通过减少根系营养元素的分配来减少根系生长,以小根系来应对土壤资源匮乏和加剧的竞争。同时,小根系形态可塑性强、C资源投资少,在贫瘠的土壤中可以迅速占领水分和养分充足的斑块,具有较强的竞争力[34- 35]。对于黑麦草而言,与对照相比,在单种和混种下其根系C、N和P元素分配比在浅土处理下显著增加,而在干旱处理下显著降低。这表明单种和混种后的黑麦草在浅土处理下通过增加对根系营养元素的分配来促进根系生长以获取更多土壤资源,而在干旱处理下通过减少对根系营养元素的分配以较少的根系资源投入来使自身在干旱胁迫下生长和生存。
表3苇状羊茅和黑麦草营养元素积累的双因素方差分析
Table3ResultsofTwo-wayANOVAtestfortheeffectsofsoilthicknessandplantingpattern,waterandplantingpattern,soilthicknesspluswaterandplantingpatternonthenutrientaccumulationofF.arundinaceaandL.perenne

物种 Species方差来源 Source of variation分子自由度ndfF营养元素积累量 Nutrient accumulation地上部分 Aboveground part根系 Root整株植株 Whole plantCNPCNPCNP苇状羊茅 土壤厚度 Soil thickness (T)119.38***20.72***38.72***0.09ns3.16+2.60ns10.85**29.99***37.57***F. arundinacea种植方式 Planting pattern (PP)10.02ns0.36ns0.00ns0.26ns1.02ns6.01*0.01ns0.97ns0.00ns土壤厚度×种植方式 T×PP10.08ns0.92ns0.01ns6.23*4.73*4.55*0.62ns1.43ns0.01ns水分 Water (W)17.98*1.23ns18.53***5.50*3.53+16.22***9.07**2.83ns20.21***种植方式 PP10.37ns0.64ns0.16ns0.60ns0.14ns6.47*0.05ns1.27ns0.13ns水分×种植方式 W×PP10.06ns0.16ns0.22ns6.11*7.25*6.32*1.07ns0.23ns0.19ns土壤厚度+水分 T+W129.75***10.49**49.08***31.74***28.73***57.00***31.49***18.97***52.39***种植方式 PP10.17ns0.86ns0.00ns0.31ns0.09ns2.80ns0.05ns1.65ns0.00ns(土壤厚度+水分)×种植方式 (T+W) × PP10.00ns0.28ns0.02ns4.20+5.12*4.96*0.51ns0.57ns0.02ns黑麦草土壤厚度 T125.75***46.34***24.05***0.87ns0.02ns1.03ns8.25*29.87***20.41***L. perenne种植方式PP10.22ns0.10ns0.00ns0.07ns0.01ns0.10ns0.28ns0.28ns0.05ns土壤厚度×种植方式 T × PP10.53ns4.83*3.26+0.37ns0.02ns0.02ns1.35ns4.71*1.19+水分 W11.48ns0.31ns11.67**9.98**6.72*11.35**4.15+0.50ns10.77**种植方式 PP10.72ns0.23ns1.97ns0.05ns0.58ns0.61ns0.21ns0.01ns1.19ns水分×种植方式 W× PP10.00ns0.26ns0.12ns0.40ns0.22ns0.09ns0.04ns0.87ns0.55ns土壤厚度+水分T+W1129.11***99.15***51.19***29.71***26.23***33.75***132.86ns99.02***45.68***种植方式 PP12.15ns4.29+2.11ns0.01ns0.07ns0.09ns1.04ns3.55+1.19ns(土壤厚度+水分)×种植方式 (T+W) × PP10.00ns0.01ns0.19ns0.08ns0.00ns0.02ns0.04ns0.01ns0.35ns分母自由度 ddf181818181818181818

图5 土壤厚度、水分及土壤厚度+水分对不同种植方式下苇状羊茅和黑麦草根系营养元素分配比的影响(平均值±标准误)Fig.5 Effect of soil thickness, water and soil thickness plus water on root nutrient distribution ratio of F. arundinacea and L. perenne under different planting patterns (M±SE)
3.2 两物种的营养元素对不同种植方式的响应
在对照资源水平下,混种后苇状羊茅的地上部分C含量显著高于单种。这表明在对照资源水平下由于生态位分化[12,16- 17],混种后深根植物苇状羊茅能够吸收和利用更多的水分和养分,进而提高植物光合作用C固定,增加体内有机化合物含量,来提高自身防御环境胁迫的能力[21]。对于黑麦草而言,其根系C含量在混种时显著高于单种,这表明浅根植物黑麦草在与深根植物竞争时,为了吸收更多的水分和养分资源,将叶片固定的C更多分配给根系,来促进根系生长,以吸收更多的水分和养分。而在3种低资源水平下,混种后苇状羊茅和黑麦草的地上部分和根系C含量与单种无显著差异。这可能是因为在低资源水平下,土壤水分和养分缺乏,无论是单种还是混种,植物吸收的水分和养分有限,导致单、混种差异小。在各资源水平下,仅混种后苇状羊茅根系P含量显著高于单种,而N含量与单种无显著差异。根系作为接受土壤水分和养分的先锋器官,其生产和更新最先受到土壤资源的影响。由于生态位互补,在土壤P缺乏的环境下,混种后深根植物苇状羊茅受到的P亏缺的抑制作用要低于单种,根系通过吸收更多P来增加其生物量的积累,延长根系生长以吸收更多的资源,进而提高自身对资源获取和竞争能力[21]。
在对照资源水平下,混种后苇状羊茅的根系C、N和P积累量和根系元素分配比显著高于单种。这表明由于生态位分化,混种促进深根植物苇状羊茅的根系营养元素的积累。同时,混种后苇状羊茅将更多的营养元素分配到根系中,以促进根系对外界环境的防御能力和对土壤水分和养分的获取和利用效率,进而提高自身的竞争力。而在3种低资源水平下,混种后苇状羊茅的各部分的营养元素积累量和根系营养元素分配比显著低于或与单种无显著差异。这表明在低资源水平下,由于两物种间的竞争加剧[11],混种后苇状羊茅通过减少根系养分元素的积累和分配来转换策略,采取低根系养分资源的投入(小根系)来避免加剧的竞争以更好共存。对于黑麦草而言,仅在浅土处理下,其根系C、N和P元素分配比在混种时显著低于单种,其他情况下其元素积累和分配在单混种间均无显著差异。这表明混种后黑麦草在某些低资源水平下还是通过减少其根系元素的分配以小根系来避免竞争。另一方面也表明,混种后黑麦草采取相对恒定的营养元素积累和分配来响应竞争。
4 结论
苇状羊茅和黑麦草能够通过增加部分营养元素的含量,降低营养元素的积累量,减少根系营养元素的分配以小根系应对土壤厚度和水分资源的减少,并在低资源水平下表现较强的资源获取和防御能力。在对照资源水平下,混种后苇状羊茅通过提高营养元素的含量、积累和分配来提高自身对资源获取和竞争能力;而在低资源水平下,面对加剧的竞争,混种后苇状羊茅通过减少其营养元素在单混种间的差异以求更好共存。在各资源水平下,黑麦草采取相对恒定的营养元素的积累和分配来响应竞争。
