豫西丘陵坡地弃耕农田植被演替对土壤碳、氮库的影响
2018-11-14李亚鸽
赵 威,李亚鸽,亓 琳,2,李 琳,王 馨
1 河南科技大学农学院/牡丹学院,洛阳 471023 2 中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085
耕作环境较差、农民进城务工等因素促使河南丘陵坡地农田弃耕现象严重[1],因而这类土地资源的动态变化与驱动因素问题备受关注[2- 3]。对弃耕农田进行土地管理,究竟是复垦,还是自然恢复?回答这个问题首先要明确其植被与土壤变化规律以及二者的响应关系。
无干扰条件下弃耕农田次生演替过程通常为分阶段循环往复式发展[4],物种组成存在延续性与递进性[5];演替进程可划分为一年生草本—一年生草本+多年生草本—草本+灌木—乔灌草4个阶段[6- 8];群落结构趋向复杂化,与临近原生植被的相似性逐渐增加[9- 11]。但弃耕农田的演替存在区域性差异。我国西北盐渍化弃耕地群落结构趋向于单一化[12- 13],而秦岭太白山地区群落复杂性则逐渐增加,分层现象明显[14]。弃耕地土壤性质的变化与地上群落演替密不可分[15- 16],例如:紫色土弃耕坡地的植被物种多样性指数与表层土壤碳、氮含量呈正相关关系[17];黄土丘陵区表层土壤有机质也随植被密度增加而逐渐积累[18- 23]。土壤理化性质对群落演替有直接影响[24- 26],有机碳、有效氮是制约弃耕地植被恢复的关键因素[27]。土地利用方式的转变可打破土壤碳、氮的库源平衡[28],黄土高原丘陵区和缙云山坡地弃耕农田表层土壤碳、氮储量均显著高于原有耕地[29- 30]。
目前有关弃耕农田研究多专注于群落演替影响因素[1,31]、土壤理化性质变化[32- 35]、植被演替方向[10,36]、复垦潜力评价[37- 38]等,而针对丘陵坡地弃耕农田演替过程中群落植物功能多样性以及植被与土壤碳、氮库变化相互关系的研究十分有限。本研究采用时空替代法[25],以豫西地区丘陵坡地弃耕农田为研究对象,分析自然演替过程中群落植物功能多样性与物种多样性、群落生物量以及碳、氮储量变化规律,探讨土壤碳、氮库对植物群落变化的响应,为明确丘陵坡地弃耕农田演替方向和再利用途径提供理论参考。
1 材料与方法
1.1 研究区概况
豫西通常是指三门峡市、洛阳市所在的河南省西部地区,总面积约2.6×103km2,西依秦岭、南望伏牛、北靠太行,地形、地貌复杂多变,是黄土高原的边缘地带,生态环境比较脆弱。土壤类型以黄绵土、棕壤土、褐土为主。豫西属温带大陆性季风气候,四季分明;也是典型的半干旱、半湿润地区,年平均温度13.5—14.5℃,年平均降水量550—650 mm。该地区的农业活动主要通过在丘陵山地开垦梯田,种植玉米(Zeamays)、花生(Arachishypogaca)、小麦(Triticumaestivum)、红薯(Ipomoeabatatas)、芝麻(Sesamumindicum)等农作物。由于受自然与水肥条件限制,农作物产量始终受到制约。
试验开展前期通过大量走访调查,选取了5个海拔高度(400—500 m)、土壤类型(黄绵土)相对一致,且无人为干扰和放牧的典型弃耕农田,其弃耕年限分别是1、3、8、15 a和25 a,构建为一个演替系列。所选样地中以多年生草本植物与灌木居多,自然植被是以酸枣+多年生草本群落为主的暖温性灌草丛。研究样地的植被状况如表1所示。

表1 调查样地植物群落物种组成
毛马唐(Digitariachrysoblephara),荩草(Arthraxonhispidus),委陵菜(Potentillachinensis),益母草(Leonurusjaponicus),夏至草(Lagopsissupina),荆条(Vitexnegundo),猪毛蒿(Artemisiascoparia),野艾蒿(Artemisialavandulifolia),白羊草(Bothriochloaischaemum),胡枝子(Lespedezabicolor),细叶苔草(Carextenuiflora),绣线菊(Spiraeasalicifolia),白酒草(Eschenbachiajaponica),博落回(Macleayacordata),罗勒(Ocimumbasilicum),紫花地丁(Violaphilippica),铁杆蒿(Artemisiasacrorum),阴地蒿(Artemisiasylvatica),黄背草(Themedajaponica),苔草(Carextristachya),狗娃花(Heteropappushispidus),莎草(Cyperusrotundus),酸枣(Ziziphusjujuba),长叶胡枝子(Lespedezacaraganae),截叶铁扫帚(Lespedezacuneata),抱茎小苦荬(Ixeridiumsonchifolium),马唐(Digitariasanguinalis)
1.2 样地调查与取样
于2016年植物生长季7—8月进行样地调查。采用5点取样法,在每个样地设置5个1 m×1 m的样方,调查植物物种组成与群落的高度、盖度、密度等数量特征。之后使用剪刀齐地面剪下所有植物的地上活体部分,同时收集样方内的凋落物,按照样方编号分别装入牛皮纸信封袋,带回实验室在65℃条件下24 h烘干至恒重,测定其干重,并留存备用做下一步的碳氮分析。
完成群落植物的调查和取样后,在每个样方中,使用直径7.5 cm根钻,按照0—5、5—10、10—20、20—30、30—50 cm的深度从上至下取样,每层3钻混合。按层分装在孔径为0.3—0.4 mm的纱袋中,并将写好取样信息的塑料标签置于纱袋内,带回室内漂洗,装进信封并做好标记,放进烘箱中65℃烘干至恒重;按照上述分层深度使用直径7.5 cm土钻每层再取3 钻土壤混合,装入自封袋中,带回室内自然阴干,然后使用0.145 mm筛网除去根系、动物残体等,留存备用做碳氮分析。
最后在5个样方中心位置挖一个体积为1.0 m×0.5 m×0.5 m(长×宽×深)的取样坑。在取样坑的边缘去除表面植物凋落物后,用环刀依次按照0—5、5—10、10—20、20—30、30—50 cm的深度从上至下取样,每层3个重复。环刀规格为高度5 cm,体积一般为100 cm3。按照不同土层做好标记带回室内105℃烘干至恒重,称重。土壤容重(g/cm3)=土壤烘干重(g)/环刀体积(cm3)。同时利用这些土壤样品进行土壤砾石比(砾石重/土壤烘干重)的测定。
1.3 样品分析与数据计算
(1)群落物种多样性采用α多样性指数:
Margalef丰富度指数:
Ma=(S-1)/Ln(N);
Pielou均匀度指数:
JP=H′/Ln(S);
Shannon-Wiener综合多样性指数:
H′=﹣∑Pi×Ln(Pi);
式中,S为样方中的物种数目,N为物种个体总数,Pi采用相对密度、相对频度、相对盖度等表示[10,39]。
(2)植物与土壤碳、氮含量测定与计算:
使用杯式粉碎机和MM400型冷冻混合球磨仪(德国 Retsch)进行样品的粗粉和细粉,过0.145 mm筛网,然后采用干烧法使用NA1500型元素分析仪(意大利 Carlo Erba)测定植物与土壤样品的全碳、全氮含量[40]。
植物碳、氮储量(g/m2)=植物碳、氮含量(%)×生物量(g/m2)×102。
(3)根据土壤碳或氮含量、容重、砾石比和土层厚度计算土壤碳或氮储量:
土壤碳或氮储量(g/m2)=∑Ci×θi×Di×(1-δi)×10
式中,Ci、θi、Di、δi分别为第i层土壤碳或氮含量(g/kg)、土壤容重(g/cm3)、土层厚度(cm)、土壤砾石比[41]。
1.4 统计分析
使用SPSS 17.0统计软件进行单因素方差分析(ANOVA),采用新复极差法(Duncan)进行平行数据间比较,采用字母标记法在P=0.05水平下进行差异显著性检验。使用Canoco 4.5软件进行群落各因子的冗余分析(RDA)。采用Excel 2016与Origin 9.0软件进行图表制作。
2 结果与分析
2.1 不同年限弃耕农田植物群落的生物多样性特征
植物的功能多样性是生物多样性的重要组成部分。如图1所示,在群落演替过程中,一、二年生草本植物与多年生草本植物在弃耕群落中所占比例较大,合计达到60%—80%,前者先降低后升高,后者变化与前者相反。在弃耕初期,就有灌木幼苗存在,群落演替到8 a时,灌木所占比例降低为10%。按照生态型将群落植物分为旱生植物与中生植物两种,前者比例维持在80%左右远大于后者。植物的光合途径类型可反映植物的初级生产能力。随着演替进行,弃耕农田植物群落C3植物的比例逐渐增加到80%,而C4植物的比例则减小。弃耕农田植物群落中物种繁殖采取有性繁殖、无性繁殖、有性兼无性的方式,整体上所占比例为有性繁殖<无性繁殖<有性兼无性。

图1 不同年限弃耕农田群落植物的功能多样性Fig.1 The plant functional diversity of farmland community in different abandoned years
物种多样性是生物多样性的数量特征。在弃耕3 a时,群落植被高度显著大于其他年限的群落植被(P<0.05),但其群落盖度最小,仅为50.20%(表2)。在弃耕25 a时,群落盖度显著大于演替前期(P<0.05),达到最大值(84.60%)。在弃耕初期(1、3 a),群落植被密度较小,随后逐渐增大;弃耕8 a时,植被密度达到163.67株/m2。随着演替进行,群落Margalef丰富度指数、Pielou均匀度指数与Shannon-Wiener综合多样性指数具有相对一致的变化规律,即先增大后减小,但三者达到峰值的时间不同,分别是第15年、第8年。
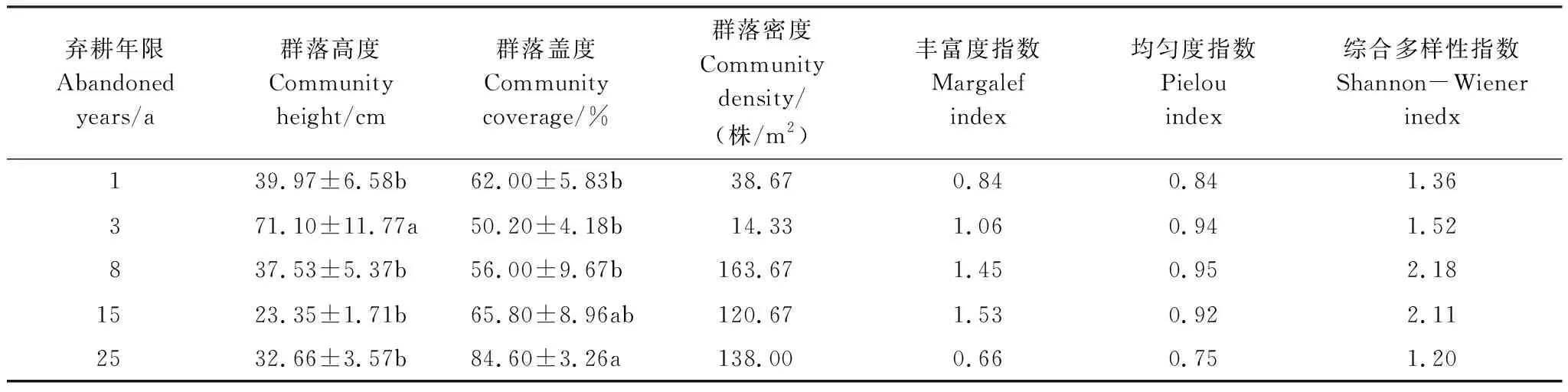
表2 不同年限弃耕农田植物群落的数量特征与物种多样性指数
同列不同小写字母表示不同年限间差异显著(P<0.05)
2.2 不同年限弃耕农田群落植被的生物量与碳、氮储量
将群落植被生物量分成3个部分:地上活体生物量、凋落物生物量与根系生物量。随着演替进行,不同年限弃耕农田植物群落地上活体生物量之间不具有显著差异(P>0.05),凋落物生物量在弃耕15 a时具有最大值(149.75 g/m2,P<0.05)(图2)。而根系生物量显著大于地上活体生物量和凋落物生物量,呈现不断增大趋势,其变化规律符合线性方程y=406.28x-37.38(R2=0.991,y、x分别表示根系生物量、弃耕年限)。在演替后期(15、25 a),根系生物量达到最大值(P<0.05),且植物总生物量与根系生物量的变化规律一致。受植物生物量影响,植物地上活体、凋落物、根系及植物总碳储量的变化规律与对应的生物量变化同步(图2)。但是,植物总碳储量在演替的15 a与25 a之间无显著差异(P>0.05)。

图2 不同年限弃耕农田群落植被生物量构成及其碳储量Fig.2 The plant biomass composition and C storage of farmland community in different abandoned years不同小写字母表示不同年限间差异显著(P<0.05)
弃耕农田群落植被总氮储量随着演替时间不断增加(图3),在演替顶级阶段(25 a)达到最大值11.80 g/m2,显著大于其他的演替阶段(P<0.01)。随着演替进行,地上活体氮储量没有显著性变化(P>0.05)。弃耕15 a时凋落物氮储量达到最大值(0.178 g/m2),且与其他演替阶段差异显著(P<0.05)。群落演替至25 a时,根系氮储量为11.35 g/m2,显著大于其他的演替阶段(P<0.05)。

图3 不同年限弃耕农田群落植被氮储量 Fig.3 The N storage of farmland community in different abandoned years
2.3 不同年限弃耕农田土壤碳储量与氮储量变化
随着群落演替的进行,土壤碳储量与氮储量的变化趋势相反,碳储量逐渐增大;而氮储量整体上逐渐减小,其中在弃耕15 a时存在一个不显著的增加变化(图4)。演替顶级阶段(25 a)的土壤碳储量显著大于其他演替阶段的碳储量(P<0.05),数值可达3.6—6.4倍。在演替的前期与中期(1—15 a),土壤碳储量比较稳定,且增量较小,变化不显著(P>0.05)。对于土壤氮储量来说,弃耕25 a时的土壤氮储量显著低于弃耕1 a时的氮储量(P<0.05),降低幅度约为1倍。

图4 不同年限弃耕农田土壤碳储量与氮储量变化Fig.4 The changes of soil C and N storage of farmland in different abandoned years
2.4 不同年限弃耕农田植物群落特征对土壤碳库与氮库的影响
土壤碳储量、氮储量与植物群落各指标间的RDA分析如图5所示。群落盖度、密度等生态因子对土壤碳储量具有正效应,影响程度由大到小依次为:群落盖度、群落密度、根系氮储量、植物氮储量、地上活体碳储量、植物总生物量、植物总碳储量、根系碳储量;而与均匀度指数、群落高度、地上活体生物量、地上活体氮储量存在较大的负相关。对土壤氮储量而言,其对均匀度指数、群落高度、地上活体生物量、地上活体氮储量之间表现为正响应,且影响程度依次递减,而与群落密度、盖度之间存在显著的负效应关系。其他生态因子如丰富度指数、综合多样性指数、凋落物碳氮储量等对土壤碳储量与氮储量的影响很小。土壤碳储量与氮储量之间具有显著的负效应,这与图4中所呈现的变化结果相一致。

图5 不同年限弃耕农田土壤碳氮储量与群落特征、植物碳氮储量关系的RDA分析Fig.5 The RDA analysis of the relationship between C, N storage of soil and community characteristics, plant C, N storage of farmland in different abandoned yearsH:群落高度 Community height;C:群落盖度 Community coverage;D:密度 Density;MA:丰富度指数 Margalef index;JP:均匀度指数 Pielou index;SW:综合多样性指数 Shannon-Wiener index;ALB:地上活体生物量 Aboveground living biomass;LB:凋落物生物量 Litter biomass;RB:根系生物量 Root biomass;PB:植物生物量 Plant biomass;ALCS:地上活体碳储量 Aboveground living carbon storage;LCS:凋落物碳储量 Litter carbon storage;RCS:根系碳储量 Root carbon storage;PCS:植物碳储量 Plant carbon storage;ALNS:地上活体氮储量 Aboveground living nitrogen storage;LNS:凋落物氮储量 Litter nitrogen storage;RNS:根系氮储量 Root nitrogen storage;PNS:植物氮储量 Plant nitrogen storage;SCS:土壤碳储量 Soil carbon storage;SNS:土壤氮储量 Soil nitrogen storage
3 讨论
豫西丘陵坡地弃耕农田群落演替的阶段性不明显,主要经历了一、二年生草本与多年生草本+灌木两个阶段,黄土高原丘陵坡耕地植被演替也遵循上述规律[42]。演替初期就有灌木幼苗存在,这可能与周围环境原生植被中灌木的种子扩散有关[43]。豫西丘陵原生植被以暖温性灌草丛为主,荆条和酸枣等灌木类物种较为常见。但退耕40多年的黄土高原植被恢复区并没有演替为灌丛或小乔木组成的群落[44],其原因主要是草本植物在群落中占绝对优势,在没有外界干扰条件下其凋落物不断堆积,从而抑制了木本植物种子萌发和幼苗定植[45]。从群落植物功能多样性角度分析,豫西丘陵坡地弃耕农田群落中多年生草本、有性兼无性繁殖物种比例较大,多年生草本植物庞大的根系及其繁殖方式有利于群落结构的稳定。如图1所示,随着演替的发展,群落中C4植物数量减少而C3植物增加,这是由于在演替早期植物群落盖度小,光照充足,资源竞争压力小,有助于C4植物占据群落空隙;相对于C4植物来说,C3植物的适应能力和竞争力更强,所以在演替后期比例增加。弃耕地群落植物整体耐旱性较强,与该地区处于半干旱、半湿润地带有关。随着群落演替进行,群落高度降低、群落盖度和密度增加,物种多样性指数先增大后减小,这与薛超玉等[19]的研究一致,表明撂荒演替过程中上述的群落变化使群落结构不断趋于稳定。
群落植被的碳、氮储量在很大程度上依赖于群落生物量,整体上均表现出逐渐增加的趋势。随着弃耕时间的延长,植被总生物量呈递增变化,这种差异主要是由根系生物量积累引起的[46]。地上活体生物量具有一定的减小趋势,一方面是植物活体凋落回归土壤,另一方面可能是多年生草本积累的光合产物更多地转移至根系。杜峰等[26]认为撂荒地群落地上生物量先减小后增加,主要受弃耕年限和土壤养分影响。在弃耕25 a时,凋落物生物量减小为71.27 g/m2,而且地上活体生物量也小于前一阶段,原因可能是群落内部环境改变使分解作用加强所导致。
按照演替系列发展,土壤生态系统表现出“固碳放氮”现象,即发挥着“碳汇氮源”的生态功能。如图5所示,群落盖度、密度、植被生物量、植被碳氮储量等与土壤碳储量具有显著的正相关关系,但它们与土壤氮储量存在负相关关系。由图2和图3可知,群落的生物量决定着植物体碳储量、氮储量的变化。通过对样品生物量、氮储量的分析发现,土壤的平均氮含量有微小幅度的下降变化,这个变化通过土壤氮储量公式被放大了约102倍。此外,凋落物生物量在弃耕15 a时较其他年份多,在该年份土壤的氮输入增多。正如图4所示,土壤氮储量在15 a时有不显著的增加变化。总的来说,土壤氮储量的变化一方面来自于土壤氮含量的下降,另一方面取决于凋落物的输入量,而凋落物的多少受植被群落密度、盖度、生物量等的影响。RDA分析也说明,弃耕农田群落植被盖度、密度及生物量是影响土壤碳氮储量的主要因子,而且土壤氮含量也是影响因子之一。土地利用方式的改变导致植被变化,植物通过光合作用固定大气中的碳,并从土壤中吸收氮素供给自身生长,促进群落盖度、密度增大和生物量的积累,形成更多的凋落物。而在群落演替后期,土壤微生物活动加快,凋落物加速分解,一部分氮素以气体形式释放到空气中,而其余氮素回归土壤[29,47]。Novara等[8]也认为农业活动停止后土壤发挥“碳汇”作用,且呈线性递增。缙云山坡耕地有机碳含量为2.58 g/kg,而在弃耕后其含量增加到14.90 g/kg,这充分说明农田弃耕演替过程使土壤“碳汇”的作用不断恢复和增强[30]。研究发现土壤有机质、全氮等随着弃耕年限的延长而积累[7,11],而且土壤碳与全氮之间具有显著的正相关关系[28]。这与本研究的结果不一致,可能与耕地撂荒之前的耕作方式有关,也可能是土壤中的氮通过一系列的化学与物理过程进入植物与大气中。另外一个不可忽略的因素是弃耕年限。李永强等[32]研究表明,土壤有机质与全氮呈现出先降低后增加的变化规律,其中大约在弃耕33 a后才表现出增加趋势。因此,在农田弃耕后土壤碳与氮的变化不同步,究竟是“汇”还是“源”,受何种因素影响,还有待于进一步深入研究。从生态系统物质循环角度分析,弃耕农田演替过程土壤对碳的固定不断增强,有助于缓解半干旱、半湿润地区丘陵坡地农田生态系统碳排放。在未来该地区的土地管理中,首先需要考虑选择的物种类型,建议适当增加群落中豆科植物的丰富度以增加土壤的氮储量,减少氮素流失。
4 结论
(1)豫西丘陵坡地弃耕农田群落演替缓慢,且阶段性不明显,大致可划分为一、二年生草本与多年生草本+灌木两个阶段,而没有遵从一年生草本—一年生草本+多年生草本—草本+灌木—乔灌草这样一个常规演替系列模式。
(2)在群落演替进程中,草本植物始终占据优势,合计达到60%—80%,且多属旱生植物;C3植物的比例逐渐增加,繁殖方式以有性兼无性繁殖为主。群落盖度在演替顶级时(25 a)达到最大值(84.60%),而群落高度与密度分别在弃耕3 a和8 a时出现最大值(71.10 cm,163.67株/m2)。群落丰富度指数、均匀度指数与综合多样性指数先增大后减小,峰值分别为1.53、0.95、2.18,但它们达到峰值的时间不同。
(3)随着演替进行,不同年限弃耕农田植物群落地上活体生物量之间不具有显著差异,凋落物生物量在弃耕15 a时达到最大值(149.75 g/kg),而根系生物量呈现不断增大趋势,且显著大于地上活体生物量和凋落物生物量。植物地上活体、凋落物、根系及植物总碳储量的变化规律与对应的生物量变化同步,总氮储量在演替顶级阶段达到最大值(11.80 g/kg),且显著大于其他的演替阶段。
(4)土壤碳储量与氮储量的变化趋势相反,碳储量逐渐增大而氮储量不断减小,但是在弃耕15 a时氮储量具有不显著的增加变化。演替顶级阶段(25 a)的土壤碳储量为960.98 g/kg显著大于其他演替阶段,而土壤氮储量(54.31 g/kg)则显著低于演替初期的氮储量(27.08 g/kg),减小了近1倍。
(5)土壤氮储量与碳储量表现出显著的负相关关系,分别受不同的生态因子影响。群落盖度、密度、根系氮储量、植物氮储量、地上活体碳储量、植物总生物量、植物总碳储量、根系碳储量等因子对土壤碳储量具有正效应;土壤氮储量与均匀度指数、群落高度、地上活体生物量、地上活体氮储量之间存在正响应关系,但是群落密度和盖度对土壤氮储量表现为显著的负作用。
