强度石漠化区不同植被修复模式下土壤螨类群落差异
2018-11-14金道超王鹏举林丹丹
陈 浒,金道超,陈 航,王鹏举,周 政,林丹丹
1 贵州大学昆虫研究所,贵州大学昆虫资源开发利用省级特色重点实验室,贵阳 550025 2 贵州师范大学喀斯特研究院/国家喀斯特石漠化防治工程技术研究中心,贵州省喀斯特山地生态环境国家重点实验室培育基地,贵阳 550001
为有效遏制石漠化的进一步加剧,促进区域生态的快速修复和农林经济的快速发展,石漠化生态修复一直是石漠化治理面临的关键问题[1]。自21世纪初以来,在以中-强度石漠化为主的贵州花江峡谷喀斯特区石漠化综合治理中,实施了“花椒(顶坛花椒,Zanthoxylumplanispinumvar.dingtanensis)[2]”、“金银花(Lonicerajaponica)”、“花椒+金银花”、“花椒+砂仁(Amomumvillosum)”、“石榴(Punicagranatum)”、“任豆(Zeniainsignis)”等不同植被修复模式,这些模式在植被生态适宜性、生态效益、经济效益和社会效益上有较好效果[3-6]。2010—2012年的监测显示,该区以土壤动物反映的生态效果也十分良好,同时显示土壤螨类是土壤动物的优势类群[7-8]。
国内外的研究表明,森林火烧、植被砍伐、土壤污染和土地利用变化会改变土壤螨类的原有结构,降低物种丰富度和多样性[9-13],而受损生态系统的恢复又使土壤螨类呈不同程度地增加,且不同地貌区域不同植被修复方式下的土壤螨类科属组成有所差异[14-18]。本研究选择贵州花江峡谷区顶坛小流域强度石漠化治理区作为研究区,对研究区内“花椒林”、“金银花林”、“花椒+金银花林”3种植被修复模式下的土壤螨类群落变化进行了对比研究,以揭示不同植被修复模式下的土壤螨类群落差异与生态恢复进展,为区域石漠化生态治理效益的后续评估提供土壤生物对比参数。
1 材料与方法
1.1 研究区概况
顶坛小流域面积1641.01 hm2,地理位置为105°38′48″—105°39′7.98″E,25°38′22.08″—25°39′50.88″N,海拔500—1200 m,夏季多持续高温,年均温18.4℃,年降水量1100 mm,无霜期300 d以上,土壤为黄壤和石灰土,土层厚度一般为5—20 cm。生态治理前石漠化面积达726.63 hm2,其中中强度石漠化面积占石漠化总面积的80.68%[5]。在强度石漠化区,主要野生植物有石岩枫(Mallotusrepandus)、构树(Broussonetiapapyrifera)、蒿草(Artemisiasp.)、荩草(Arthraxonhispidus)等,植被类型为稀疏灌草。
金银花、花椒是该区域最为适生的经济植物,是实施经济植物修复中使用的主要物种。在该小流域内实施的多种经济植物修复模式中,“花椒”、“金银花”、“花椒+金银花”模式维持时间长:花椒10年,金银花5年。
1.2 样地设置与样品采集
以强度石漠化生态修复区作为研究样区,在样区内分别选择花椒林、金银花林和花椒+金银花林作为样地,在各样地内地势相对平缓区域分别设置(1—2 m)×(1—2 m)样方,每样地设置样方6个,相同样地的样方比邻以确保样方土壤质地的相对一致性,每个样方每期设1个不重复样点。
2014年1月、4月、8月和10月,在样方内按期(季)采集土样样品,每期每样点采用10 cm×6.4 cm的圆筒形不锈钢土壤环刀连续上(0—6.4 cm)、下(6.4—12.8 cm)层采集土壤样品2个,共采集样品136个。所有样品分别用棉布袋分装并编号后带回室内分离土壤动物。
1.3 螨类标本的收集与鉴定
螨类标本的收集:供试土壤采用Tullgren法分离土壤动物,分离时温度保持在35℃以下,连续烘烤48 h。然后将盛有土壤动物的培养皿置于显微镜下,用小毛笔和软镊再将土壤螨类从土壤动物中分离,并对分离出来的标本用70%酒精固定和清洗,清洗后的螨类标本保存于盛有乳酸溶液的小塑料管中进行透明。
螨类标本的鉴定:将透明后的螨类标本制作装片后置于显微镜下,主要参考《A Manual of Acarology》(Third Edition)[19]、《蜱螨学》[20]、《中国土壤动物检索图鉴》[21]、《中国东北土壤革螨》[22]和《中国粉螨概论》[23]等进行鉴定,所有标本鉴定到属级单元(若螨和体型不完整标本除外)。文中螨类分类阶元主要采用《A Manual of Acarology》(Third Edition)[19]的分类系统。
1.4 数据处理
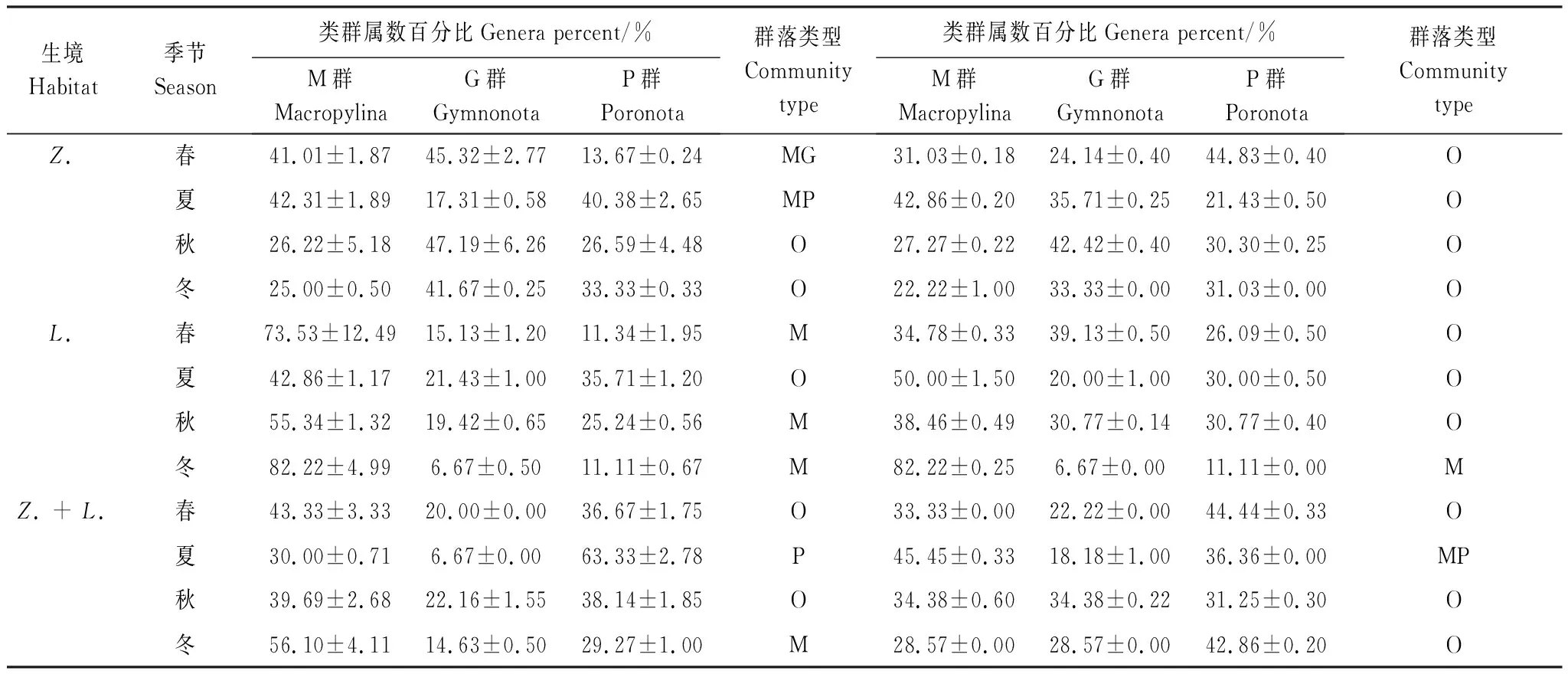
数量优势度:按郑祥等[24]的方法进行划分,即个体数占总捕获量的百分比,>10%为优势类群(+++),1%—10%为常见类群(++), Shannon-Winner多样性指数(H′)、Margalef丰富度指数(SR)和Pielou均匀度指数(J)[25]: H′ =-∑(Pi× lnPi) 式中,Pi为属i的个体数占全部个体数的比例; SR= (S-1)/lnN 式中,N为土壤螨类群落全部类群的个体总数; J=H′/lnS 式中,H′为Shannon-Winner多样性指数,S为属数。 Bray-Curtis相似性指数(CN)[26]: CN=2jN/ (Na+Nb) 式中,Na、Nb分别为样地Aj、Bj的总个体数,2jN为每个种在两个样地都存在的较低多度值之和。相似性划分见表1。 表1 群落相似性划分 捕食性螨类成熟度指数(MI)[13,27]: 式中,s为土壤螨群落革螨类属数,Ki为第i属所属科的K值,ri为第i属所属科的r值。 甲螨MGP分析[28-29]:M,大孔低等甲螨(Macropylina);G,无翅坚背甲螨(Gymnonota);P,有翅孔背甲螨(Poronota)。分别计算各类群种类数百分比进行MGPⅠ分析和各类群个体数百分比进行MGPⅡ分析。群落类型划分见表2。 数据分析在Excel 2007和SPSS 19中完成,群落差异通过方差分析(ANOV),差异显著性水平取P<0.05。 对3种植被模式下的土壤螨类调查,共捕获土壤螨类1372头,隶属3目55科89属。其中,中气门目(Mesostigmata)13科21属,绒螨目(Trombidiformes)6科6属,疥螨目(Sarcoptiformes)36科62属。土壤螨类在各生境的分布为花椒林3目44科66属,金银花林2目31科50属,花椒+金银花林3目41科56属(表3)。螨类属数由多到少的排序为花椒林、花椒+金银花林、金银花林。 表2 甲螨群落类型划分 M:大孔低等甲螨,Macropylina;G:无翅坚背甲螨,Gymnonota;P:有翅孔背甲螨,Poronota;O:全部,Overall;MG:大孔低等-无翅坚背甲螨,Macropylina-Gymnonota;GP:无翅坚背-有翅孔背甲螨,Gymnonota-Poronota;MP:大孔低等-有翅孔背甲螨,Macropylina-Poronota 数量优势度方面,花椒林有3个优势属:懒甲螨属(Nothrus)、盖头甲螨属(Tectocepheus)和合若甲螨属(Zygoribatula) (表3),优势属、常见属和稀见属的百分组成为4.55%、24.24%和71.21%;金银花林有2个优势属:罗甲螨属(Lohmannia)和懒甲螨属,优势属、常见属和稀见属的百分组成为4.00%、36.00%和60.00%;花椒+金银花林的优势属是懒甲螨属,优势属、常见属和稀见属的百分组成为1.79%、41.07%和57.14%。显见,懒甲螨属是3种模式下共同的优势属,不同模式下的优势属类群组成存在差异,优势属、稀见属的百分组成按花椒林、金银花林、花椒+金银花林顺序逐渐减少,而常见属的百分组成则按此顺序增加。 不同模式下除科数上的差异外,从5个多属科即厉螨科(Laelapidae)、罗甲螨科(Lohmanniidae)、奥甲螨科(Oppiidae)、单翼甲螨科(Haplozetidae)和大翼甲螨科(Galumnidae)组成属数上看,花椒林分布有21属,金银花林分布有19属,花椒+金银花林分布有15属,显示了不同模式下多属科在属数组成上的差异(表3)。在科的个体数量上,盖头甲螨科(Tectocepheidae)、懒甲螨科(Nothridae)和若甲螨科(Oribatulidae)是花椒林的优势科,罗甲螨科和懒甲螨科是金银花林的优势科,单翼甲螨科和懒甲螨科是花椒+金银花林的优势科,显示了不同模式下优势科的差异。 从表3还看出,不同模式下的螨类属,除了优势属的差异外,3种模式下的共有属有29属;花椒林和金银花林有共有属35属,花椒林和花椒+金银花林有共有属40属,金银花林和花椒+金银花林有共有属35属,无论是3种模式还是两两模式对比,共有属的比例均<50%,显示不同模式间螨类属数组成的差异较大。分布于各模式单一生境的不同螨类有35属,包括花椒林的18属、金银花林的8属和花椒+金银花林的9属,其中花椒林螨类属数和个体数量均以无翅坚背甲螨、有翅孔背甲螨和革螨(Gamasina)占优势;金银花林的大孔低等甲螨、无翅坚背甲螨和革螨各属数虽占优势但各属数相差不大,个体数量则以大孔低等甲螨占优势;花椒+金银花林的螨类属数以革螨和尾足螨(Uropodina)略占优势,个体数量上以革螨占优势。 表3 土壤螨类群落组成 续表属GenusZ.L.Z. + L.合计Total个体数(优势度)个体数(优势度)个体数(优势度)个体数(优势度)Ind.(Dom.)Ind.(Dom.)Ind.(Dom.)Ind.(Dom.)迭蜕甲螨属Plateremaeus3(+)1(+)4(+)裸珠足甲螨属Gymnodamaeus7(++)7(+)平壳甲螨属Palatyloides2(+)2(+)珠甲螨属Damaeus1(+)1(+)表珠甲螨属Epidamaeus2(+)1(+)3(+)贝氏棱甲螨属Berlesezetes1(+)1(+)2(+)沙甲螨属Eremulus7(++)6(++)5(++)18(++)窝甲螨属Fosseremus10(++)6(++)18(++)34(++)大龙骨足甲螨属Megeremaeus1(+)1(+)跳甲螨属Zetorchestes1(+)1(+)溯甲螨属Birnsteinius2(+)1(+)1(+)4(+)丽甲螨属Liacarus3(+)1(+)4(+)麦考瑞奥甲螨属Macquarioppia4(+)1(+)5(+)小盾珠甲螨属Suctobelbella1(+)1(+)2(+)弓奥甲螨属Arcoppia3(+)3(+)长矛奥甲螨属Lanceoppia9(++)3(+)7(++)19(++)多奥甲螨属Multioppia8(++)5(++)1(+)14(++)奥甲螨属Oppia8(++)1(+)9(+)枝奥甲螨属Ramusella1(+)1(+)梯奥甲螨属Trapezoppia25(++)23(++)11(++)59(++)自发甲螨属Autogneta2(+)2(+)盖头甲螨属Tectocepheus111(+++)9(++)8(++)128(++)垂盾甲螨属Scutovertex3(+)1(+)4(+)吕克奥甲螨属Lucoppia1(+)2(+)3(+)若甲螨属Oribatula3(+)3(+)合若甲螨属Zygoribatula58(+++)8(++)19(++)85(++)原结甲螨属Prinodus1(+)1(+)树穴甲螨属Truncopes1(+)6(++)10(++)17(++)菌甲螨属Scheloribates1(+)1(+)足肋甲螨属Podoribates8(++)4(+)12(+)美奥甲螨属Caloppia2(+)3(+)7(++)12(+)单翼甲螨属Haplozetes3(+)1(+)1(+)5(+)圆单翼甲螨属Peloribates1(+)1(+)8(++)10(+)细单翼甲螨属Pilobatella1(+)1(+)针单翼甲螨属Setoxylobates15(++)13(++)35(++)63(++)威单翼甲螨属Vilhenabates5(++)9(++)14(++)木单翼甲螨属Xylobates3(+)14(++)8(++)25(++)点肋甲螨属Punctoribates4(+)4(+)顶翼甲螨属Acrogaluma1(+)1(+)大翼甲螨属Galumna8(++)3(+)1(+)12(+)全大翼甲螨属Pergalumna1(+)1(+)原大翼甲螨属Protokagalumna1(+)2(+)3(+)毛大翼甲螨属Trichogalumna1(+)9(++)11(++)21(++) Z.:花椒林,Zanthoxylumplanispinumvar.dingtanensisforests;L.:金银花林,Lonicerajaponicaforests;Z.+L.:花椒+金银花林,Zanthoxylumplanispinumvar.dingtanensis+Lonicerajaponicaforests;Ind.:个体数量Individual numbers;Dom.:优势度,dominance;+++:优势类群,Dominant groups;++:常见类群,Common groups;+:稀有类群,Rare groups 由前述2.1,螨类属数排序为花椒林>花椒+金银花>金银花林。在土壤层变化上,秋季的花椒林下层和花椒+金银花林上层具有丰富的属;季节变化上,金银花林的春季、花椒林和花椒+金银花林的秋季具有丰富的属(图1)。不同模式下的螨类属数在土壤上、下层之间以及相同层位上差异均不显著(P>0.05),而秋季分别与夏季和冬季之间则差异显著(P<0.05)。 个体数量的排序为金银花林(529头)>花椒林(526头)>花椒+金银花林(418头)。土壤层变化上,个体数量较高分别出现于春季金银花林的上层、秋季花椒林和花椒+金银花林的上层;季节变化上,金银花林的春季、花椒林和花椒+金银花林的秋季拥有较高的个体数量(图1)。不同模式下螨类个体数量的差异性变化与螨类属数的差异性变化是相同的情形。 个体密度的排序为金银花林(6072.19头/m2)>花椒+金银花林(5324.84头/m2)>花椒林(4467.09头/m2)。土壤层变化上,最高出现在春季金银花林的上层;季节变化上,花椒林的春季和秋季、金银花林的春季和冬季、花椒+金银花林的秋季和冬季拥有较高的个体密度,其中金银花林的春季拥有最高的个体密度(15668.79头/m2)(图1)。各模式下的个体密度在土壤层和季节差异上均不显著(P>0.05)。 垂直变化上(图1),除秋季和冬季的花椒林螨类属数表现出微弱的逆分布外,其他均表现为表聚性(主要聚集分布于土壤上(表)层时即表现为表聚性,反之则为逆分布);个体数量上,除冬季的金银花林为逆分布和花椒林上、下层相等外,其余均表现为表聚性;个体密度上,除冬季的金银花林为逆分布外,其他均表现为表聚性。 图1 土壤层螨类属数、个体数量与个体密度的水平和垂直分布Fig.1 Horizontal and vertical distribution of genera numbers, individual numbers and individual density on soil mites under different vegetation modesZ.:花椒林,Zanthoxylum planispinum var. dingtanensis forests;L.:金银花林,Lonicera japonica forests;Z.+L.:花椒+金银花林,Zanthoxylum planispinum var. dingtanensis + Lonicera japonica forests 不同植被修复模式下,金银花林拥有较高的多样性指数,花椒+金银花林拥有较高的丰富度指数,均匀性指数的变化不明显(图2)。 在季节变化上,春季的花椒林拥有较高的多样性指数和丰富度指数;各季节均匀性指数的变化不大(图2)。各模式下,春季分别与夏季和冬季、夏季与秋季多样性差异显著(P<0.05),丰富度指数和均匀性指数在各季节差异均不显著(P>0.05)。 3种不同生境下土壤螨类的Bray-Curtis相似性指数(CN)值分别为:花椒林-金银花林:0.45;花椒林-花椒林+金银花林:0.47;金银花林-花椒林+金银花林:0.44。表明各生境间土壤螨类群落的相似性较低,且均表现为中等不相似。 图2 土壤螨类群落多样性、丰富度和均匀性的一致性与季节变化Fig.2 Consistency and seasons change of diversity (H′), richness (SR) and evenness (J) on soil mites under different vegetation modesH′:Shannon-Winner 多样性指数,Shannon-Winner diversity index;SR:Margalef丰富度指数,Margalef richness index;J:Pielou均匀度指数,Pielou evenness index 捕食性革螨(Gamasina)K和r的赋值参照Ruf[27]、吴东辉[13],其中蠊螨科根据Karg[30]植绥螨分类参比植绥螨进行赋值(表4),各生境中革螨的MI指数值见表5。结果显示,金银花林夏季的螨类以K选择型为主,花椒林与金银花林的秋季和冬季螨类为r选择型,其他均以r选择型为主。 表4 捕食性土壤革螨(Gamasina)的K-值和r-值 表5 捕食性土壤革螨(Gamasina)的成熟度指数(MI)值和主要生态类群 MI:捕食性螨类成熟度指数,Maturity index of predatory mites;K:有利于竞争能力增加的选择,The choice that is advantageous to enhancement of competition ability;r:有利于增大内禀增长率的选择,It is beneficial to increase the choice of intrinsic growth rate 不同模式下土壤甲螨群落结构-MGP分析见表6,结果显示,类群属数上3种模式均为O型;个体数百分比上花椒林为O型,金银花林为M型,花椒+金银花林为MP型,从数值上看,除花椒林以G群为主外其他两种模式均以M群为主。 不同季节甲螨群落结构显示,花椒林在类群属数上春、夏季分别为MG和MP型,秋、冬季为O型,在个体数量上均为O型;金银花林在属数上春、秋、冬季为M型,夏季为O型,个体数量上春、夏、秋季为O型,冬季为M型;花椒+金银花林春、秋季为O型,夏季和冬季分别为P型和M型,个体数量上春季、秋季和冬季为O型,夏季为MP型(表7)。整体看,研究区域的土壤甲螨主要为O型和M型。 表6 不同模式下甲螨的群落结构 表7 不同模式下不同季节甲螨的群落结构 在天然林,土壤螨类的类群丰富,个体数量多,生物多样性指数大,群落垂直分布表聚性明显[31-32]。不同类型和不同程度的干扰则对土壤螨类群落结构有不同程度影响:森林火灾后,螨类的物种数减少,丰富度和多样性降低,原有群落的平衡关系丧失,不同过火程度环境的土壤螨类存在差异[9-10];森林植被遭砍伐后,甲螨的密度降低,中气门和前气门螨类的物种数也减少,但多样性指数未发生显著变化[11];在不同的土地利用下,土壤螨类的类群数、个体密度、群落多样性和丰富度会降低,中气门亚目、甲螨亚目螨类比前气门亚目和无气门亚目螨类对土壤耕作的影响更为敏感[13,33],少耕和免耕更有利于土壤螨类群落结构的稳定性及多样性的保持[34];重金属的污染在个体水平上还会导致甲螨(Chamobatescuspidatus)附肢(腿)的残缺[12]。这些变化与不同影响下凋落物的破坏与移除、环境条件的改变等有关[11,13,33]。 在植被修复环境,如俄罗斯哈巴罗夫斯克市从耕地、禾本科草地、豆科草地、高山植物到苔藓云杉林35年的植被恢复,江西余江退化红壤区旱生草坡、稀疏针叶林、针叶林和针阔混交林 4种植被修复,纽约市中部石灰岩、砂岩、页岩区种植冬黑麦草进行侵蚀控制以及松嫩草原中度和重度退化草地种植苜蓿、围栏封育和种植碱茅的植被恢复中,土壤螨类的物种数、个体数量和个体密度呈不同程度的增加;同时,俄罗斯哈巴罗夫斯克市35年植被恢复后,土壤螨类群落结构变得复杂,表层物种和非特有物种比例增加,深层物种减少,群落组成趋于稳定并随植被和气候表现带性特征。相关研究表明,土壤螨类群落结构的差异与植被类型、植被年龄、郁闭度、凋落物重量、土壤特征等综合因素有关[13,35-37],植被的恢复和干扰的减少有利于有机物的积累、土壤湿度增加和微环境多样性的形成,有利于土壤螨类群落结构的恢复[14-18]。本研究中,以恢复期较长的花椒林和花椒+金银花林土壤拥有丰富的螨类科数和属数,恢复期较短的金银花林土壤则拥有较高的个体数量和个体密度,但由于研究区土壤螨类群落多样性指数值相对较低,且花椒林的螨类属数和金银花林的螨类个体数量与个体密度还存在一定的逆分布,表明研究区的土壤螨类仍处于恢复变化中。 优势类群是生物适应环境和指示环境变化程度的重要指标,强度石漠化生态修复环境与其他生态修复环境的优势类群存在较大差异。松嫩草原中度退化草地的围栏封育生境以吸螨属(Bdella)占优势,种植苜蓿以下盾螨属(Hypoaspis)占优势;重度退化草地的围栏封育生境以吸螨属、下盾螨属占优势,种植碱茅以原大翼甲螨属(Protokalumna)、吸螨属、肉食螨属(Cheylteus)占优势[17-18]。在江西余江退化红壤区的不同植被修复环境,旱生性植被以单翼甲螨科(Haplozeidae)、若甲螨科(Oribatulidae)和矮汉甲螨科(Nanhermanniidae)为优势,稀疏针叶林以单翼甲螨科和矮汉甲螨科为优势,针叶林以盖头甲螨科(Tectocepheidae)、奥甲螨科(Oppiidae)和矮汉甲螨科为优势,针阔混交林以喜螨科(Terpnacaridae)、珠足甲螨科(Belbidae)和矮汉甲螨科为优势[16]。而在本研究区,花椒林以懒甲螨属、盖头甲螨属和合若甲螨属为优势属,金银花林以罗甲螨属和懒甲螨属为优势属,花椒+金银花林以懒甲螨属为优势属。其中,罗甲螨属、懒甲螨属属大孔低等甲螨,宜于人为影响较小的环境栖息[38],合若甲螨属可能与生态修复中凋落物的积累有关[39],盖头甲螨属常栖息于受干扰或早期演替环境[19],表明研究区的土壤环境虽已得到初步恢复,但花椒林的土壤环境恢复较慢。 3种生境中只在各单一生境分布的35属不同土壤螨类,其优势类群包括革螨股、尾足螨股、大孔低等甲螨、无翅坚背甲螨和有翅孔背甲螨的一些属。有研究显示,不同土壤螨类具有一定的生态学特性差异,其中革螨喜潮湿环境[22,40];尾足螨中糙尾足螨属、内特螨属和二爪螨属主要栖息于腐烂有机质、凋落物和好的土壤环境[19,41-42];大孔低等甲螨丰富可能与土质疏松和腐殖质含量增加有关[33];无翅坚背甲螨中裸珠足甲螨属、大龙骨足甲螨属主要分布于干旱环境,自发甲螨属在潮湿和干旱环境中均有分布,珠足甲螨属主要分布于腐殖质和凋落物环境[19,43];有翅孔背甲螨中若甲螨属、顶翼甲螨属和全大翼甲螨属栖息于多种环境[19],细单翼甲螨属、点肋甲螨属主要分布于森林土壤和凋落物环境[44-45]。因此从植被恢复使土壤湿度增加[14-15]和研究区尚存一些喜干旱环境分布的螨类类群推断,该区土壤环境的恢复速度较慢,以花椒林的更为明显。 土壤捕食性革螨生态对策类型和甲螨生态类群可以反映土壤受扰动的状况,在干扰较小的环境,捕食性革螨以K选择型为主,在顶级常绿阔叶林、自然林和防护林,甲螨群落为O型[16,27,46]。从不同植被恢复下的捕食性革螨和甲螨生态类群看,江西余江退化红壤区的植被修复中,除针阔混交林外其他生境均有K选择类群的存在,甲螨群落为P型、O型和M型[16],松嫩草原中度退化草地的恢复生境,捕食性革螨以K选择型所占的比例较大[17],表明它们的生态恢复已取得较好效果。本研究区的捕食性革螨主要为r选择型,其中金银花林的夏季主要为K选择型,甲螨生态类群主要为O型和M型,表明研究区的土壤螨类和土壤环境的恢复已取得一定效果。 在强度石漠化生态修复区,不同植被修复模式下土壤螨类的群落组成和结构均表现出一定的差异,可利用优势属和具有典型生物学与生态学特性差异的土壤螨类对石漠化生态恢复的进程和效果进行初步指示。由于喀斯特地区土壤斑块的大小、厚度、质地、肥力等与降雨、植被、土壤微生物、基岩特性、地上地下连通性、人类活动程度等密切相关,具有强烈的异质性[47-48],喀斯特地区尤其是喀斯特石漠化地区土壤螨类的研究相对匮乏,因此,要深入认识石漠化地区不同植被修复环境土壤螨类群落结构的差异及其反映环境修复的程度,还有待对环境因子差异与螨类群落结构关系、基本分类单元(物种)及其参与的生态过程等开展深入研究。 致谢:感谢安明态教授对植物标本的鉴定和朱佳运、付超凡、吕小溪等对样品采集的帮助。
2 结果与分析
2.1 螨类组成差异

2.2 多属科、优势科的差异
2.3 属的差异
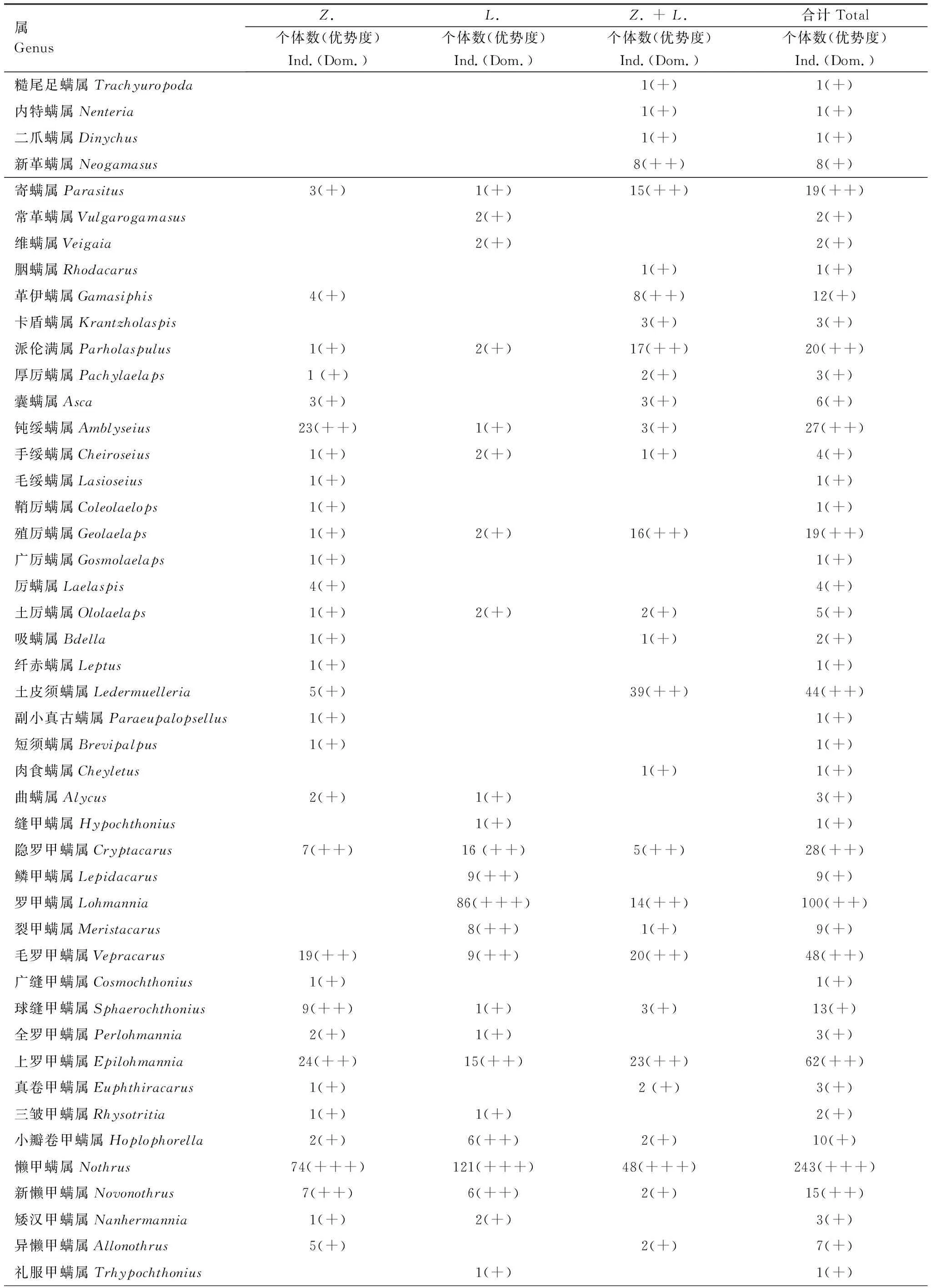

2.4 螨类水平和垂直结构差异

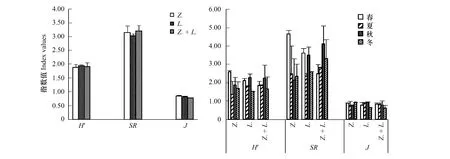
2.5 群落多样性差异与相似性
2.6 捕食性革螨群落结构的差异


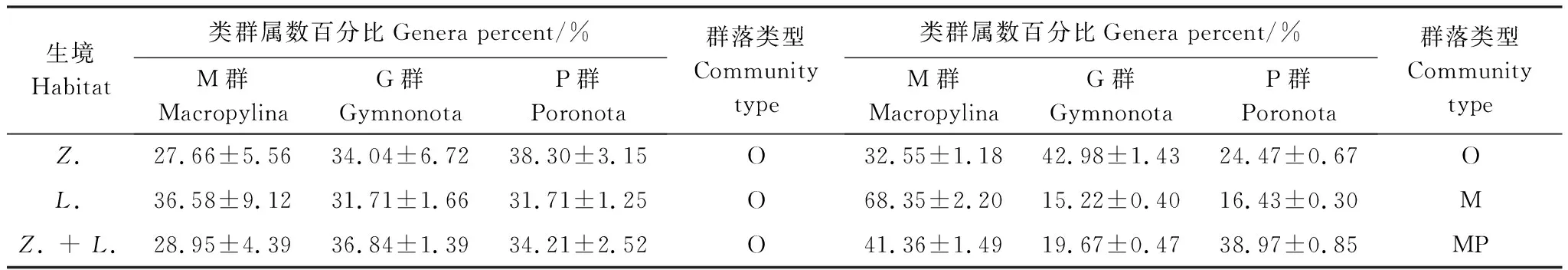
2.7 甲螨群落结构的差异
3 讨论
4 结论
