脂联素对骨形成的调节及其机制的研究新进展
2018-10-24万丽娟龚燕平李春霖
万丽娟 龚燕平 祖 源 李春霖
解放军总医院南楼内分泌科,北京 100853
近年来脂肪组织分泌的各种细胞因子越来越受到研究者的关注,如瘦素、脂联素、肿瘤坏死因子等,这些脂源性细胞因子对糖代谢、动脉粥样硬化、炎症、胰岛素抵抗等方面产生关键性作用。脂联素是一种具有保护作用的脂肪源性的细胞因子,是体内唯一与体质含量呈负相关的脂肪因子,近年来的研究发现其与骨代谢关系密切。本文就近年来有关于脂联素骨调节的作用及其机制的研究综述如下。
1 脂联素概述
在1995年,Scherer等[1]率先发现了由脂肪细胞分泌的脂联素,并因其分子量为30kDa将其命名为Acrp30。之后发现脂联素由定位于3q27的APM1基因编码,其基因长度为16kb,由2个内含子及3个外显子构成[2]。247个氨基酸组成了脂联素的一级结构,包括N端的信号序列、高度可变区、胶原蛋白区及C末端的球状结构域。脂联素与肿瘤坏死因子-α(TNF-α)、补体C1q及胶原蛋白在结构上有同源性,提示脂联素的起源与TNF-α密切相关。
人血浆中的脂联素水平高达0.5~30mg/L其总量占蛋白总量的0.01%,其中血浆中的脂联素有三种存在形式,分别为三聚体、六聚体及多聚体,在血液循环中脂联素绝大部分以多聚体形式存在(大约占85%),是具有生物活性脂联素的主要存在方式[3-4]。脂联素在结构上可分为小球状片段型脂联素及全长型脂联素,这两种不同结构的脂联素通过不同的脂联素受体、作用于机体不同的组织来发挥其的生物学作用。Waki H等[5]研究发现小球状片段型脂联素是由中性粒细胞和(或)单核细胞激活后分泌产生的白细胞弹性蛋白酶裂解全长型脂联素而得来的。
2003年,Yamauchi等[6]采用分子克隆技术克隆出了两种脂联素受体的异构体,分别为脂联素受体1(AdipoR1)与脂联素受体2(AdipoR2),它们均属于七次跨膜的受体家族,但它们与经典的七次跨膜的G蛋白偶联受体不同,脂联素受体的C端位于细胞膜外,且AMP蛋白激酶(AMPK)通路是它们主要的信号转导通路。早期的研究发现为AdipoR1在骨骼肌细胞存在表达,而AdipoR2在肝细胞存在表达,胰岛素β细胞及内皮细胞等均表达AdipoR1与AdipoR2这两种受体[6]。近年来的研究发现为脂联素这两种受体在全身多种组织细胞广泛表达,如脂肪组织、肝、肾、脑、肺、睾丸、骨骼肌、成骨细胞及破骨细胞等,并且其表达存在组织差异。钙粘蛋白-T(T-cadherin)是最近发现的一种脂联素新型受体,其与脂联素结合后激活细胞内的信号转导通路从而发挥生物学作用。
2 脂联素促进成骨细胞增殖分化的作用及可能机制
2.1 脂联素有促进成骨细胞增殖的作用
2004年,Berner等首次[7]应用免疫组化的技术在人离体的成骨细胞以及小鼠的体颚骨中检测到了脂联素,并验证了人体胫骨及股骨培养的原代成骨细胞和小鼠的成骨细胞系MC3T3-E1中均存在脂联素mRNA的表达,脂联素mRNA的表达和分泌与成骨细胞的分化阶段有关。同时从人骨中提取的原代成骨细胞培养观察到了AdipoR1和AdipoR2的表达,AdipoR1在小鼠成骨细胞系MC3T3-E1中也存在表达。Oshima K等[8]发现脂联素可以提高成骨细胞系MC3T3-E1碱性磷酸酶(alkaline phosphatase,ALP)的mRNA表达及其矿化的活性;将具有活性的脂联素注射到实验组小鼠体内,发现松质骨的体积明显高于对照组,骨小梁骨矿物质含量(Bone Mineral Content,BMC)增加,提示了脂联素有促进成骨细胞增殖,并提高其活性的作用。
2.2 脂联素促进成骨细胞增殖的作用机制
Luo X等[9]研究发现脂联素通过诱导激活p38丝裂原的活化蛋白激酶(mitogen-activated protein kinase,MAPK)及c-Jun的氨基末端激酶(c-Jun N-terminal kinase,JNK),促进成骨细胞的分化与增殖,增强ALP的活性,提高血浆1型胶原氨基端前肽(Procollagen I N-terminal Propeptide,P1NP) 与骨钙素(osteocalcin,OCN)的生成量,增加骨髓基质的矿化量。P38 MAPK的阻断剂SB203580可以使脂联素诱导ALP的活性增加受阻;JNK的阻断剂SP600125预处理可以使脂联素诱导的成骨细胞分化增殖受阻;而AdipoR1的小干扰RNA则同时有以上两种阻断作用。因此脂联素受体/JNK信号途径介导了成骨细胞的增殖,脂联素受体/p38MAPK信号途径介导了成骨细胞的分化。此外,有研究[10]发现重组脂联素对人成骨细胞RANKL的诱导及骨保护素(osteoprotegerin,OPG)mRNA的抑制呈剂量与时间依赖性,AdipoR1的siRNA可以消除脂联素调控的RANKL及OPG mRNA的表达。MAPK信号通路在重组的脂联素诱导RANKL激活及抑制OPG的表达中也起着重要的作用。
骨形态发生蛋白(Bone morphogenetic proteins,BMPs)对成骨细胞分化及骨生成起重要作用。有研究者发现,脂联素在体外培养的成骨细胞中可通过AdipoR1促进BMP-2 mRNA的表达,但不能使BMPs的其他家族成员表达增加;5’-AMP激活性蛋白激酶 (5’-AMP-activated protein kinase,AMPK)的siRNA及AMPK抑制剂可抑制脂联素介导的成骨细胞BMP-2的表达;加入特异性p38/NF-κB抑制剂或使p38/NF-κB突变, 脂联素诱导的BMP-2表达可被抑制。因此脂联素通过AdipoR1激活AMPK,p38/NF-κB通路促进成骨细胞BMP-2生成,可能是脂联素促进成骨分化和生成的机制之一[11]。
2.3 脂联素在模型动物和临床研究中的作用存在争议
对于脂联素基因敲除/过表达模型动物的研究结果,目前并不完全一致。Mitsui[12]等对高表达脂联素的转基因小鼠的研究显示,12周时模型鼠的骨密度和骨形成明显增加。Wu Y等[13]的研究显示,脂联素基因敲除鼠的骨小梁结构和骨密度明显降低。脑室注射脂联素后,小鼠灰色脂肪组织的解耦联蛋白(uncoupling protein 1,UCP1)表达明显减少,血浆肾上腺素和去甲肾上腺素的水平明显下降,神经元亮氨酸拉链蛋白结构(leucine zipper motif -1,APPL-1)表达增加,骨小梁体积在脂联素基因敲除鼠和对照鼠中均增加。脂联素有促进骨髓基质细胞和间质细胞向成骨细胞的分化的作用。而Ealey[14]等的研究认为,脂联素过表达小鼠的股骨BMC和股骨颈峰值骨含量明显低于对照小鼠。Williams GA的研究显示,与对照鼠比较,雄性脂联素基因敲除鼠的骨小梁容积增加约30%,骨小梁数量增加了38%。Wang F等[15]对脂联素基因敲除鼠的研究发现,其成骨细胞的RunX2和Osterix的表达明显增加;骨髓间质细胞向成骨细胞的分化和细胞钙化明显增加。对于这一矛盾结果的产生的原因,目前尚不明确。但有学者认为[16],基因敲除/过表达鼠存在除骨骼外多个器官组织功能的改变,尤其是脂肪组织,而脂肪组织调节的脂肪代谢对骨代谢的影响是不容忽视的;此外基因改变后小鼠的破骨细胞的功能也相应发生改变,成骨/破骨的平衡被打破。因此对脂联素基因敲除/过表达模型骨骼的改变及机制有待更多的研究证据证实。
数个临床研究的结果也并不理想。Tenta R等[17]对81位绝经后女性(其中43位为低骨量)的研究并未发现脂联素与BMD之间的关联。MohitiArdekani J 等[18]对81位非糖尿病的骨质疏松患者的研究显示,血浆脂联素水平与股骨颈和脊柱BMD呈负相关关系。Wu 等[19]对336位绝经后中国女性的血浆脂联素水平、BMD和骨转化指标的研究显示,脂联素水平与BMD成负相关。设计更为合理的临床研究,尤其是前瞻性研究,更有效的避免糖尿病、甲状腺疾病、性别、年龄、药物等因素对骨质疏松评估的影响,对于进一步研究脂联素在人体对骨骼的影响就显得尤为重要。
本研究也对脂联素与小鼠骨折修复的关系进行了研究。我们的研究发现,骨折后局部应用脂联素可以促进骨痂的生长,骨折的愈合。Western blot显示局部应用脂联素后,骨折局部BMP2水平的增高是其可能机制之一,但小鼠血浆的脂联素和BMP2的水平并没有明显差异。因此我们认为,脂联素确实有促进骨生成的作用,但作用的时机、疗程和部位都有可能对其作用产生很大的影响。这也可以解释为何脂联素基因模型鼠和人体临床观察试验的结果并不理想。我们已经在进行小鼠全身应用和局部应用脂联素对骨折修复作用的比较(相关的文章整理投稿中),希望为脂联素对成骨的作用机制提供更多理论依据。
3 脂联素对破骨细胞的双向调节作用及可能机制
3.1 脂联素抑制破骨细胞的作用及其机制
2005年Oshima K等[8]发现,将具有活性的脂联素注射到实验组小鼠体内,除了有促进成骨增殖的作用,还同时伴有破骨细胞数量的减少,脂联素可以抑制破骨细胞的增殖与活化。Wang QP等[20]对脂联素基因敲除鼠的研究显示,与对照鼠比较,模型鼠的破骨细胞数目较少,骨密度、骨小梁数目和骨容量更高,RANKL水平较低,而骨保护素(OPG)的水平更高。因此认为脂联素可通过OPG/RANKL途径抑制破骨细胞作用。Luo E等[21]的研究显示,脂联素抑制破骨细胞分化的作用与作用时间关系密切。持续给予脂联素16天以上,破骨细胞的增殖和重吸收功能被明显抑制。对破骨细胞增殖基因的抑制,以及对凋亡基因的上调,是脂联素抑制破骨细胞的可能机制。YG等[22-24]应用APPL1的特异靶位小干扰RNA发现,脂联素通过APPL1下调Akt1的活性,对破骨细胞的骨吸收产生抑制作用。Yamaguchi N等[25]的研究显示,脑室注射脂联素后,脂联素基因敲除鼠和对照鼠的破骨细胞数量、破骨细胞生成标志降低,而骨小梁体积增加。中枢应用脂联素不仅有促进骨髓基质细胞和间质细胞向成骨细胞分化的作用,而且有抑制破骨细胞分化的作用。
3.2 脂联素对破骨细胞分化的抑制作用及其机制
此外,脂联素还可以抑制单核细胞集落刺激因子(macrophage colony stimulating factor,M-CSF)及RANKL诱导的小鼠骨髓巨噬细胞及人CD14阳性单核细胞向破骨细胞的分化,并抑制破骨细胞骨吸收的活性。有研究证实脂联素可以抑制RANKL诱导的RAW264.7细胞向破骨细胞的转化,并下调RANKL的上游调控基因,包括激活性T细胞的细胞核因子2(nuclear factor of activated T cells 2,NFAT2),肿瘤坏死因子受体相关因子6(tumor necrosis factor receptor-associated factor 6,TRAF6),组织蛋白酶K(cathepsin K),及酸性磷酸酶(tartrate-resistant acid phosphatase,ACP)[24]。Wu Y等[13]发现脂联素使RAW264细胞的Toll样受体所介导的NF-κB活化受阻,从而抑制了破骨细胞的形成。
3.3 脂联素对破骨细胞的促进作用及其机制
Luo X等[10]发现脂联素诱导人成骨细胞RANKL表达,阻断OPG表达的作用,对破骨细胞有间接的促进作用。进一步在成骨细胞及外周血单核细胞的共培养体系中加入MAPK抑制剂SB203580进行预处理,结果SB203580阻断了脂联素调节的RANKL及OPG mRNA的作用,抑制了破骨细胞的形成。提示脂联素对破骨细胞的作用可能是双向的。重组脂联素有可能通过AdipoR1/ p38-MAPK蛋白通路诱导破骨细胞的增殖和分化[9]。Wang F等[15]对脂联素基因敲除鼠的研究发现,其成骨分化和钙化明显增加,但并未发现模型鼠有明显骨密度的改变,而模型鼠的碱性磷酸酶活性和破骨细胞数量也明显增加。卵巢切除也不影响脂联素基因敲除鼠的骨密度、碱性磷酸酶活性和破骨细胞数量。因此认为脂联素有同时促进成骨细胞和破骨细胞分化增殖的作用,对调节骨形成和骨吸收平衡起着重要作用,对绝经后的骨骼保护有重要作用。Lubkowska A等[26]对脂联素与骨密度关系的相关动物和临床研究进行了总结,认为脂联素对破骨细胞的作用及其对骨密度的影响,存在很多的争议,有待进一步的研究证实。
4 脂联素间接影响骨形成分化的内分泌代谢机制
研究表明,脂联素对骨生成的作用不仅仅只通过通过旁分泌和自分泌途径,而且还通过内分泌代谢途径来影响骨生成:骨组织局部生成的脂联素通过旁分泌或自分泌的代谢途径来促进骨生成,而血液循环中的脂联素通过内分泌的代谢途径抑制骨生成。
脂联素是调节代谢和胰岛素敏感性的重要脂肪因子。脂联素通过促进胰岛素的磷酸化信号这一特定途径可对骨生成起积极作用。有研究[27]显示,脂联素基因缺陷型的纯合子小鼠的骨髓干细胞的促进骨生成的作用明显低于野生型的小鼠;在培养骨髓干细胞的培养液中加入胰岛素以后,可部分纠正脂联素基因缺陷组骨髓干细胞的生长抑制。研究认为脂联素促进了胰岛素诱导的胰岛素受体底物-1及Akt下游区分子的磷酸化作用,脂联素通过影响胰岛素信号转导通路间接的促进了骨生成。Liu L等[28]的研究进一步发现,siRNA敲除成骨细胞中的PPARγ或AdipoR1后,可以阻止与3T3-L1细胞共培养的成骨细胞中的RunX2 mRNA的表达的减弱。而RunX2对成骨细胞的发育与骨形成不可或缺,是转录因子Runt结构域家族的成员[29]。共转染PPARγ会降低成骨细胞RunX2和碱性磷酸酶的活性。研究认为脂联素与其受体结合使MAPK及PPARγ配体的活性增加,从而使葡萄糖摄取及脂肪酸氧化增加,良性调节了骨的能量代谢,促进骨生成。因此,脂联素可能通过脂肪细胞PPARγ对成骨细胞的调节起重要作用。
此外,还有研究显示脂联素可通过MAPK信号通路转导途径使造血干细胞增加并使其功能受到影响,调节间充质细胞的分化成熟,抑制骨髓前脂肪细胞分化为脂肪细胞的过程[30-31]。脂联素可以通过中枢和周围神经的作用机制来抑制破骨细胞的分化,促进骨髓间充质干细胞(bone marrow derived stroma cells,BMSCs)向成骨细胞的转化,从而促进骨生成[32]。
综上所述,脂联素为脂肪细胞分泌的一种细胞因子,对骨形成的调节起着重要作用。脂联素及其受体在成骨细胞和破骨细胞均存在表达,并通过自分泌、内分泌或旁分泌的作用机制来影响成骨细胞与破骨细胞的活性及分化。脂联素作用的不同部位、疗程和时机,有可能导致了脂联素对骨代谢平衡调节的不同,成骨和破骨的作用相辅相成。脂联素调节成骨和破骨细胞的可能信号通路总结见图1。脂联素对调节骨代谢平衡有着重要作用。深入地研究脂联素促进骨形成的作用及调节机制将有助于更进一步了解脂联素与骨生成的关系,并可能为临床上与骨生成相关的疾病,如骨折,骨质酥松症等的治疗提供新的临床思路及临床途径,以指导临床上的治疗。
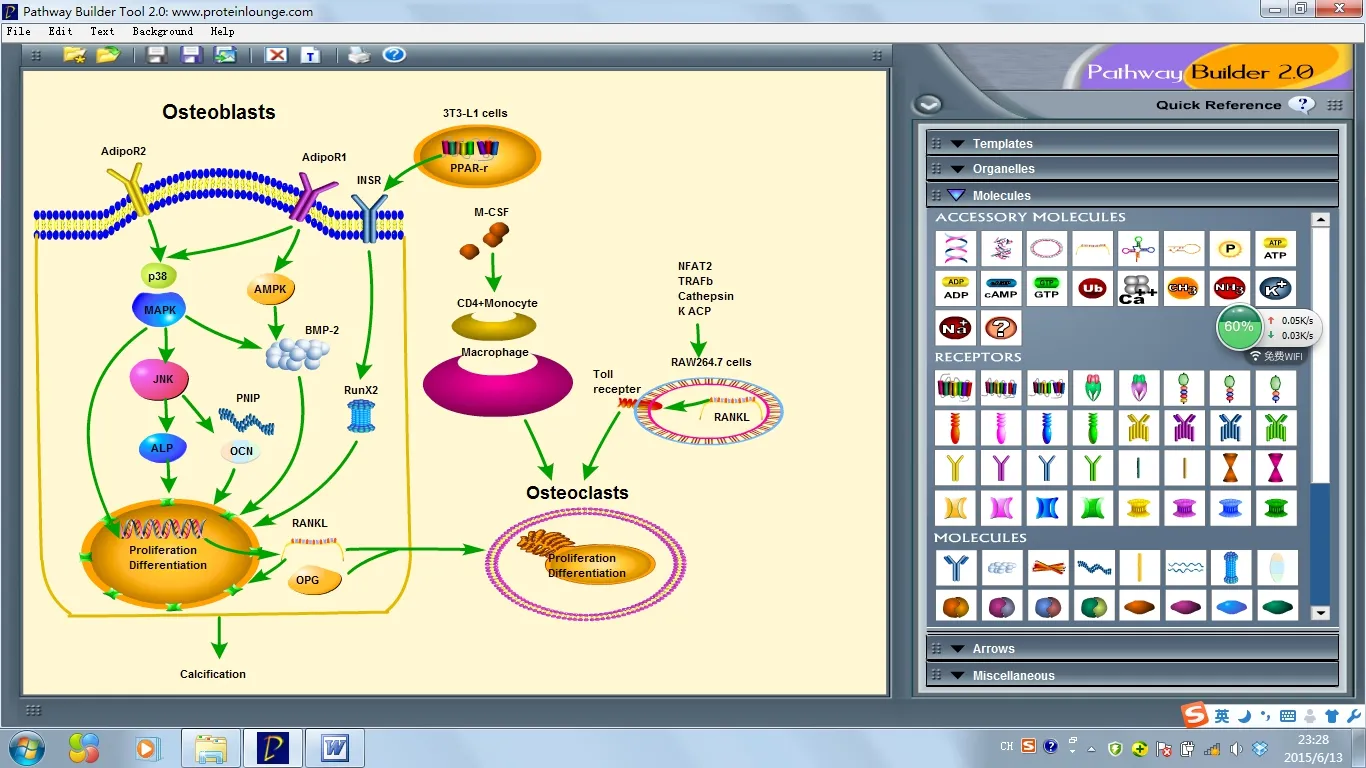
图1 脂联素调节成骨和破骨细胞的可能信号通路
ACP:酸性磷酸酶 (tartrate-resistant acid phosphatase);ALP: 碱 性 磷 酸 酶(alkaline phosphatase);AMPK:5’-AMP激活性蛋白激酶(5’-AMP-activated protein kinase);BMP-2:骨 形态 发 生 蛋 白 -2(Bone morphogenetic protein-2);Cathepsin: 组织蛋白酶; JNK: c-Jun的氨基末端激酶(c-Jun N-terminal kinase);M-CSF:单核细胞集落刺激因子(macrophage colony stimulating factor);MAPK:p38丝裂原的活化蛋白激酶(mitogenactivated protein kinase);NFAT2:激活性 T细胞的细胞核因子2(nuclear factor of activated T cells 2);OCN:骨 钙 素(osteocalcin);OPG:骨 保 护素(osteoprotegerin);P1NP:1型胶原氨基端前肽(Procollagen I N-terminal Propeptide);TRAF6:肿瘤坏死因子受体相关因子6(tumor necrosis factor receptor-associated factor 6)。
