5种水东哥属 (猕猴桃科) 植物的花粉形态及分类学意义*
2018-10-23杨晨璇向建英
李 璐 杨晨璇 罗 艳 向建英
(1. 西南林业大学云南生物多样性研究院,云南 昆明 650224;2. 中国科学院西双版纳热带植物园热带森林生态学重点实验室,云南 西双版纳 666303)
水东哥属 (SaurauiaWilld.) 是猕猴桃科 (Actinidiaceae) 3个属中最大的一个属,约有300种,主要分布在亚热带、热带亚洲和美洲,自锡金、不丹、尼泊尔、缅甸、印度、中国西南部和南部、越南、印度尼西亚至美洲均有分布[1-2]。其中,我国有13种,主产云南和广西,稀分布于四川、贵州、广东和台湾[3]。
水东哥属的系统位置不稳定,长期颇有争议[4]。它曾被放在第伦桃科 (Dilleniaceae)[5]、猕猴桃科[6-9],或自成水东哥科 (Saurauiaceae)[10-11],但近年来的分子系统学支持把它放在猕猴桃科[12-13]。南美[14-15]和越南[16]关于水东哥属的分类学研究主要集中在局部地区的水东哥属分类学修订,虽然水东哥属的研究资料不多,但现有研究表明该属为亚洲和美洲间断分布,物种数目较多,其物种多样性研究值得广泛开展。如细胞学研究认为水东哥属为染色体数目变异较大的古多倍体类群,表现出一定的细胞地理分化[17-19]胚胎学方面,尼泊尔水东哥 (S.napaulensis) 具有胚囊极性反转的现象[20],这在被子植物胚胎学发育中极为少见[21]。花粉形态学方面,研究报道的水东哥属种类较少,零星加起来不到20种[22-24]。本实验选择了5种水东哥属植物为研究对象 (4种产自中国,1种为来自南美),利用扫描电镜观察了它们的花粉形态特征,包括形状大小和外壁纹饰等,通过与近缘类群的花粉形态比较,阐明水东哥属花粉形态的分类学意义。
1 研究材料与方法
材料均取自于中国科学院昆明植物所标本馆 (KUN) 的馆藏腊叶标本 (表1)。用镊子轻轻破碎花粉囊后,用解剖针拨出花粉洒在贴好导电胶的样品台上,喷金镀膜后,利用日立S4800扫描电镜观察拍摄,所有花粉微观结构描述术语参考文献 [24-26]。

表1 5种水东哥属花粉的材料来源Table 1 Materials for pollen observation in 5 species of Saurauia
2 5种水东哥属植物的花粉形态特征
5种水东哥属植物的花粉形态数据见表2,扫描电镜下花粉超微形态特征见图1。5种水东哥的花粉形态很相似,均为长球形,花粉大小属于小粒花粉范围 (10~25 μm)[24]。最小的花粉出现在红果水东哥,大小为12.06~13.36 μm × 8.25~9.47 μm;最大的花粉为水东哥,大小为20.20~21.09 μm × 14.29~14.36 μm,大小相差近一倍。孔沟为三孔沟或三拟孔沟型,内孔不明显或内孔明显呈矩形,沟膜明显被粗颗粒状突起;外壁纹饰具穴状、细穿孔、或粗网纹穿孔的皱波状。成熟花粉的极面观为三裂圆形,败育花粉的极面观为三裂三角形。具体描述如下。
2.1 绵毛水东哥
从图1-1~4可以看出:花粉形状为长球形,极轴与赤道轴比值为1.42;极面观为三裂圆形,等极,极面截平;小粒花粉,花粉大小为13.27~15.70 μm × 9.36~10.64 μm;三孔沟型,沟较宽,沟长占极轴的65%~72%,内孔明显,矩形;外壁在沟边光滑,无沟桥,沟膜表面具颗粒;外壁纹饰浅波状,具细穿孔。
2.2 墨西哥水东哥
从图1-5~8可以看出:花粉长球形或近长球形,极轴与赤道轴比值1.19;极面观为三裂圆形,等极;小粒花粉,花粉大小16.67~20.42 μm × 14.08~16.90 μm;三拟孔沟型,沟较宽,两端渐尖,沟长占极轴的85%~87%,内孔不明显,外壁在沟边光滑,稍隆起呈沟桥,沟膜表面具小颗粒状物,不甚明显;外壁纹饰皱波状,具细穿孔。
2.3 红果水东哥
从图1-9~12可以看出:花粉形状长球形,极轴与赤道轴比值1.43;花粉败育,极面观为三裂正三角形,亚等极;小粒花粉,花粉大小12.06~13.36 μm × 8.25~9.47 μm;三拟孔沟型,沟较宽,沟长占极轴的71%~75%,内孔不明显,外壁在沟边倾斜呈坡面,沟膜表面具密集大颗粒;外壁纹饰皱波状,具细穿孔。
2.4 尼泊尔水东哥
从图1-13~16可以看出:花粉形状长球形,极轴与赤道轴比值1.48;花粉败育,极面观三角形,亚等级;小粒花粉,花粉大小12.55~15.57 μm × 9.24~10.45 μm;三拟孔沟型,沟较宽,沟长占极轴的75%~85%,内孔不明显,外壁在沟边倾斜呈坡面,沟膜表面具密集大颗粒;外壁纹饰皱波状,具深穴状或粗网状穿孔。
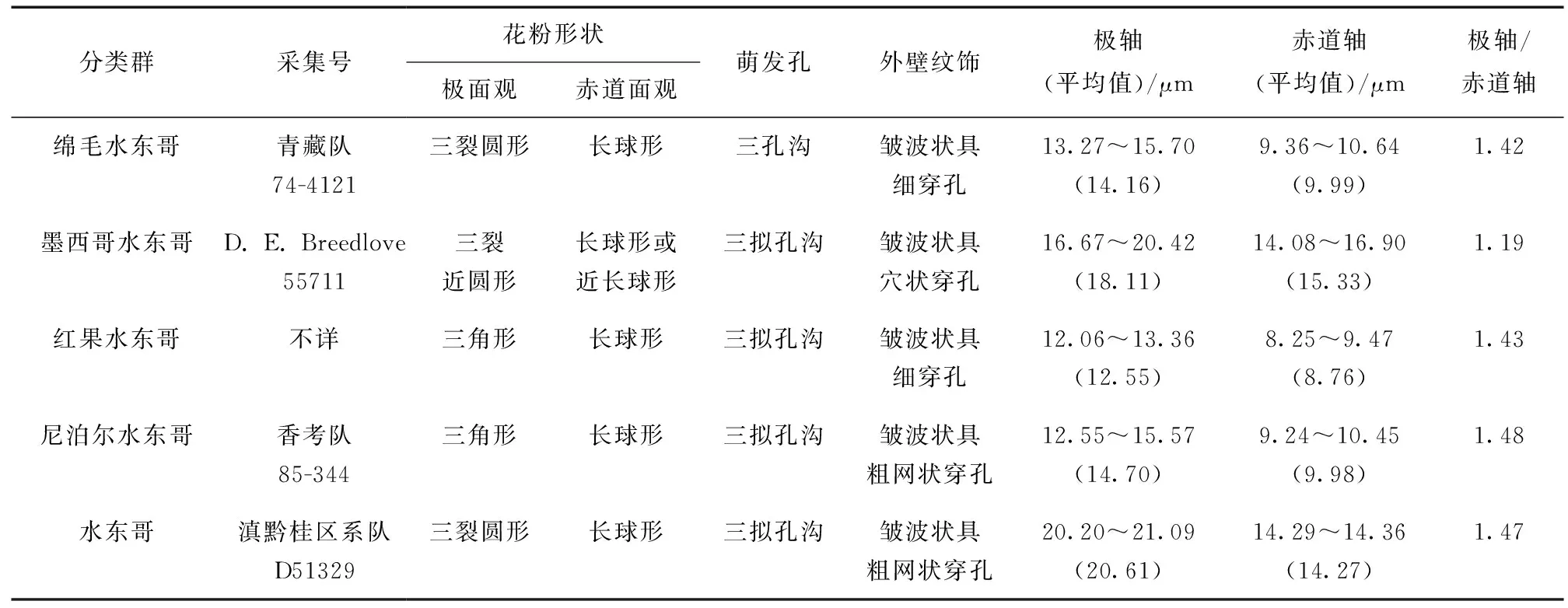
表2 5种水东哥属植物的花粉形态Table 2 Pollen morphology in 5 species of Saurauia
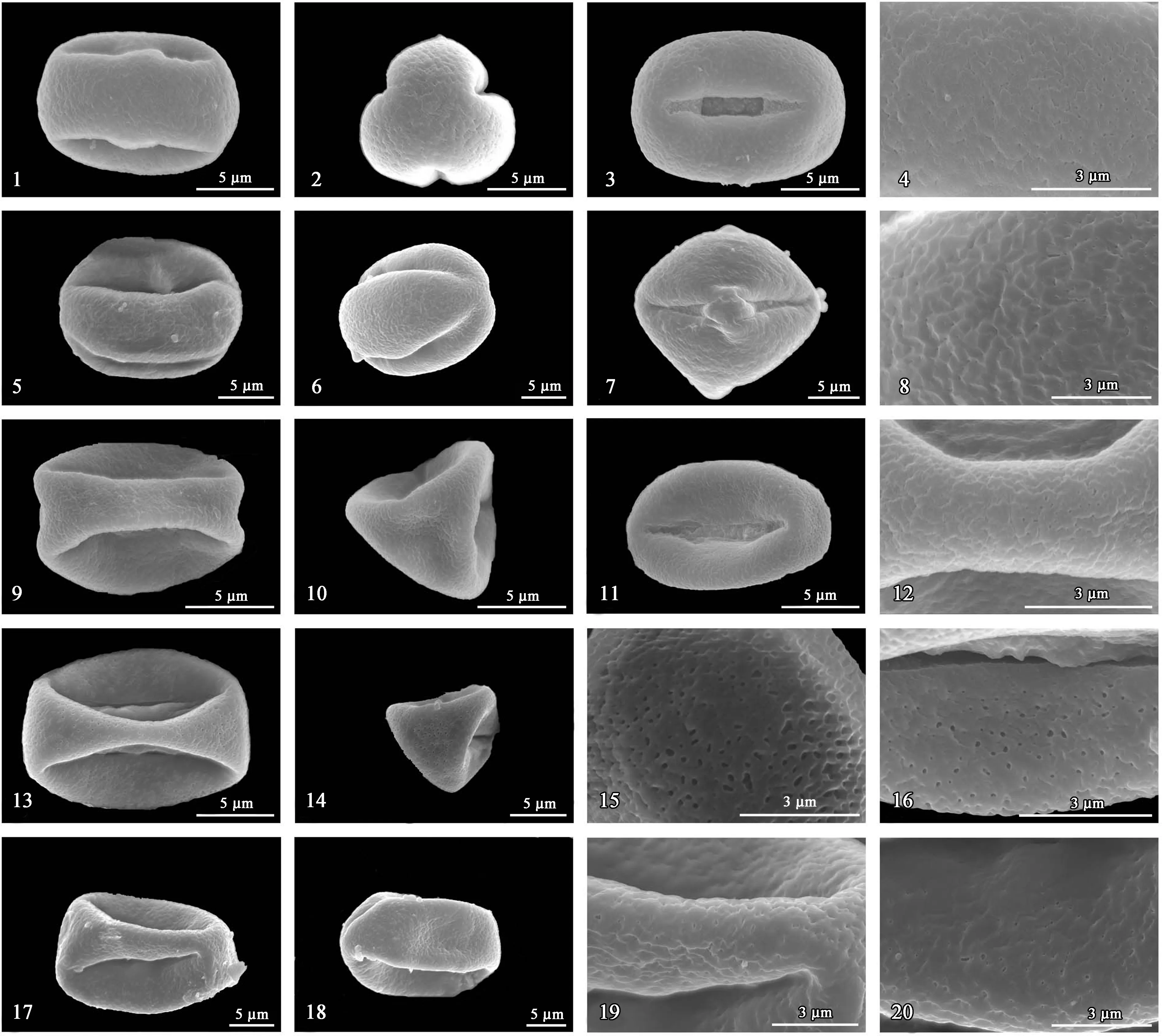
1~4为绵毛水东哥;5~8为墨西哥水东哥;9~12为红果水东哥;13~16为尼泊尔水东哥;17~20为水东哥。
示赤道面,极面观和外壁纹饰。
图1扫描电镜下5种水东哥属植物的花粉形态
Fig.1 Pollen morphology under SEM in 5 species ofSaurauia
2.5 水东哥
从图1-17~20可以看出:花粉形状长球形,极轴与赤道轴比值1.47,极面观三裂圆形,亚等级;小粒花粉,花粉大小20.20~21.09 μm × 14.29~14.36 μm;三拟孔沟形,沟较宽,沟长占极轴的70%~75%,内孔不明显,外壁在沟边光滑;外壁纹饰皱波状,具深穴状穿孔。
3 水东哥属花粉形态的系统分类学意义
3.1 花粉形态
本研究的观察结果中,4种水东哥的花粉萌发沟的内孔都不明显,为三拟孔沟花粉,只有绵毛水东哥的花粉萌发沟的内孔明显,呈矩形,属于三孔沟型花粉。5种水东哥的花粉外壁纹饰近光滑,但在纹饰类型方面差异明显。绵毛水东哥为浅波状纹饰具细孔状穿孔,其余4种为皱波状纹饰。其中,墨西哥水东哥和红果水东哥的具细穿孔,尼泊尔水东哥和水东哥具网状或穴状穿孔。因此,花粉孔沟类型和外壁纹饰特征可以作为水东哥属物种划分依据之一。
花粉外壁纹饰上[22-27],猕猴桃属和藤山柳属的皆为颗粒状或皱波状突起;而水东哥属的花粉外壁纹饰近光滑,为细孔状或粗网状穿孔的皱波状纹饰。换言之,花粉形态特征支持猕猴桃属和藤山柳属的亲缘关系较近,属于姐妹群;而水东哥属与这2个属的亲缘关系较远,这也许是水东哥属被处理为单型的水东哥科或水东哥亚科 (Saurauioideae) 的一个佐证[22-23]。同时,水东哥属一些花粉形态特征在猕猴桃科内及其近缘类群 (第伦桃科、山茶科、厚皮香科等) 中也很少见。如萌发孔类型 (矩形或圆形,或无)、萌发沟数目 (3-孔沟或多沟)、花粉败育 (是否来自功能性的雌株) 以及花粉为单粒或四合花粉等。
萌发孔是花粉萌发生长的器官,孔的数目、位置和形状在不同的植物类群中都各不相同,具有重要的分类学意义[23,26]。本研究观察到绵毛水东哥具有明显的矩形内孔,为三孔沟花粉;其他4种水东哥的内孔不明显,为三拟孔沟花粉。矩形内孔在张芝玉研究的3种水东哥中都有描述[23],但Dickenson等研究过的9种水东哥都没有观察到矩形内孔,而是描述为具圆形或卵圆形内孔的三孔沟花粉[23];其中,包括本文观察的尼泊尔水东哥和水东哥。然而,本实验观察到的这2种水东哥的花粉内孔不明显,为三拟孔沟。这也许是不同研究中观察的材料处在不同发育时期或来自不同居群所致,需进一步澄清。在广义猕猴桃科中,矩形内孔目前仅出现在水东哥属[23],其他3个属及其近缘类群如山茶科、第伦桃科、山柳科等尚未有报道[22-28]。其他被子植物中,矩形内孔在蔷薇科 (Rosaceae) 的3个亚科中也很普遍,可以作为该科的鉴定特征之一[29-30]。
3.2 四合花粉
水东哥属的花粉多为单粒[22-24],有研究表明菲律宾产的S.elegans具有四合花粉的现象[22]。本实验观察的5种水东哥的花粉也为单粒,没有观察到四合花粉。四合花粉是指花粉成熟散粉时,二细胞型花粉粒没有从胼脂质中释放出来,而是呈四分体的形式散粉[21]。在猕猴桃科中,四合花粉仅出现在水东哥属[22,25],其他2个属 (藤山柳属和猕猴桃属) 皆为单粒[22,27]。在被子植物中,四合花粉出现在番荔枝科 (Annonaceae)、罗摩科 (Asclepiadaceae)、夹竹桃科 (Apocynaceae)、杜鹃花科 (Ericaceae) 以及兰科 (Orchidaceae),花粉聚合成块是植物与传粉生物协同进化的一种适应性机制[21,28]。有意思的是,现在分子系统学把猕猴桃科归为广义杜鹃花目的核心类群[12-13]。因此,猕猴桃科的水东哥属出现了四合花粉也许证实了它和杜鹃花科在系统进化上的某种联系。随着资料的不断积累,水东哥属四合花粉的系统演化意义将会得到更好的理解。
3.3 败育花粉
本研究观察到2种水东哥的花粉颗粒干瘪,极面观为三角形,呈败育状,这与美洲具有雄花-两性花异株的S.veraguensis产生的败育花粉很相似[[27]。花粉败育是指植物的有性繁殖过程中,出现了雄配子体发育不正常的现象,即没有产生正常形态和功能的二细胞型花粉,影响了双受精的繁殖过程[21]。然而,水东哥属的败育花粉并不影响植物的有性繁殖,而是为吸引传粉昆虫的一种策略[27]。在南美洲分部的一些水东哥属植物中,存在着雄花-两性花异株现象[15]。生长在哥斯达黎加的水东哥表现为雄花-两性花异株的形态特征,但实际功能却分化为雌雄异株[27]。雄株产生的花粉颗粒大而饱满,具有萌发沟,落在两性花的柱头上可正常萌发生长;相反,两性花花药中产生的花粉颗粒小且干瘪,没有萌发沟,落在柱头上不能萌发。类似的雌雄异株的花粉功能分化现象在同科的猕猴桃属也存在,中华猕猴桃 (Actinidiachinensis) 的雄花植株产生正常的花粉粒,而两性花植株的花粉生产败育花粉,功能分化为雌株[29]。研究表明,被子植物中的雌雄异株的功能分化[28,31-32]和进化途径大约有5种[31],水东哥属雌雄异株现象也许是通过雄性-两性花异株演化而来[27]。目前,水东哥属中,雄花-两性花异株现象仅见于南美洲的部分种类,在热带亚洲分布的种类尚未有类似报道。因此,本实验观察到的产败育花粉的2种水东哥是否也存在着相似的雌雄异株功能分化现象,需要继续开展野外调查和繁殖生物学研究进一步澄清。
综上所述,水东哥属的花粉形态特征具有重要的分类学价值,可作为划分属内种间和科内属级水平的划分依据。其次,水东哥属表现出来的矩形内孔、四合花粉等在近缘类群中较为少见,具有重要的系统演化意义。此外,花粉败育现象在水东哥属的性别分化和适应性进化方面的科学意义值得进一步研究。
致谢:本研究依托西南林业大学云南生物多样性研究院科研实验平台完成,中国科学院昆明植物研究所标本馆支持花粉材料,中国科学院昆明植物研究所任宗昕博士协助扫描电镜观察。
