线粒体代谢与干细胞的命运决定
2018-10-22马明慧马悦佼孙婕张海燕
马明慧 马悦佼 孙婕 张海燕
作者单位:100069 北京,首都医科大学基础医学院2014级基础医学班专业1,细胞生物学系2
由于干细胞在再生医学和细胞治疗领域具有广泛的应用前景,越来越多的研究者致力于解析它们自我更新、多能性和可塑性的调控方式[1]。最近的研究证实,线粒体及有氧代谢直接参与干细胞多能性的调控。细胞通过生物发生、循环、融合和分裂等方式调节线粒体数量和活性以适应环境和细胞内部的因素。除了通过氧化磷酸化(oxidative phosphorylation,OXPHOS) 在产能过程起重要作用之外,线粒体在氨基酸、脂肪酸和类固醇代谢以及活性氧产生的细胞信号传导、钙平衡和细胞凋亡中均起着重要作用。本文综述近几年有关干细胞干性维持及定向分化过程中,线粒体代谢改变与线粒体形态、结构和功能变化的研究进展。
一、干细胞中线粒体的形态、结构与功能
干细胞是一类具有较强自我更新和可分化成多种细胞类型的细胞。依据其来源,干细胞分为三类:胚胎干细胞(embryonic stem cells,ESCs)、成体干细胞、诱导多能性干细胞(induced pluripotent stem cells,iPSCs)。依据干细胞的潜能,干细胞又分为全能干细胞、多能性干细胞和单能性干细胞。其中多能性干细胞是目前研究最为广泛的干细胞类型。
研究表明[2],相较于成体细胞能量产生主要依赖于OXPHOS,多能性干细胞主要基于低氧状态下的糖酵解。多能性干细胞中的线粒体形态结构也与之相适应,呈现出独特的变化。
(一)线粒体的形态及数量
与成体细胞中线粒体中所具有发达的嵴、密集的基粒数量、呈细长或有分支的复杂结构相比,利用透射电子显微镜观察人和小鼠的ESCs时惊奇地发现,ESCs中的线粒体呈不成熟的球形、数量较少且分布在细胞核周围[3-6],线粒体内具有不发达的嵴和低电子密度的基质[7]。而在ESCs的体外分化过程中,观察到线粒体形态的延伸和嵴超微结构的成熟[4-5]。线粒体DNA的含量和质量均明显增加。
(二)线粒体的结构
尽管多能性干细胞能量的产生主要依赖于糖酵解,但是多能性干细胞的线粒体中仍具有完整的呼吸链复合体结构[8]。
(三)线粒体的功能
多能干细胞中能量的来源主要通过糖酵解产生,这一产生方式导致细胞内乳酸不断累积,OXPHOS受到限制,从而使耗氧量和活性氧(reactive oxygen species,ROS)产量减少,底物进入三羧酸循环受限,糖酵解中间产物进入磷酸戊糖途径,并作为底物维持核苷酸的自我更新、合成,如图1所示。
多能干细胞中呈现出球形、不成熟线粒体,定位在核周,以电子密度较低的基质以及不发达的嵴为特征。在分化细胞中,其线粒体网则以更高电子密度和成熟嵴为特征的[10],通过OXPHOS活动确保ATP产量。这导致了氧消耗的升高和ROS产量,并减少了经糖酵解产生的乳酸的量。有趣的是,抑制OXPHOS或刺激糖酵解阻碍了干细胞分化,同时支持其重新编程为iPSCs。相反,刺激线粒体生物合成促进细胞分化,而抑制糖酵解削弱了重新编程。
在多能性干细胞低氧的糖酵解过程中,细胞产能相对较为低效,推测多能性干细胞选择糖酵解作为主要能量产生方式的原因可能有以下几点:(1)糖酵解与较低水平OXPHOS的共同作用可能为多能性干细胞生长增殖、生物合成提供反应所必需的辅助因子和酶作用物。此观点在Pereira等学者对癌细胞的实验中得以验证。Pereira等[11]发现,多能性干细胞和癌细胞获取能量主要依赖于糖酵解,即使在有氧环境下,它们仍偏好糖酵解的方式。这种方式与细胞增殖相关,通过提供反应所需因子,满足细胞分裂时生物合成的需要。设法干预代谢途径,迫使细胞改变其偏好,可以增加干细胞的多潜能性或影响其分化程度,也改变癌细胞的侵袭性。(2)糖酵解过程可降低ROS产生。ROS能破坏遗传物质和细胞组分,维持低水平的ROS可保护细胞不受上述伤害,对生物体的发育和维持内环境稳态有重要意义[12]。有报道表明,过表达氧化还原因子Ref-1可以通过抑制细胞内ROS的产生来减少人骨髓间充质干细胞的衰老[13]。(3)由于多能性干细胞存在于缺氧的微环境中,糖酵解代谢可能是它们对生活环境的适应性表现。尽管糖酵解的能量产生效率较低,但其产能速度快,且伴有较低水平的活性氧形成,这对缺氧条件下维持细胞多潜能性很重要[14]。
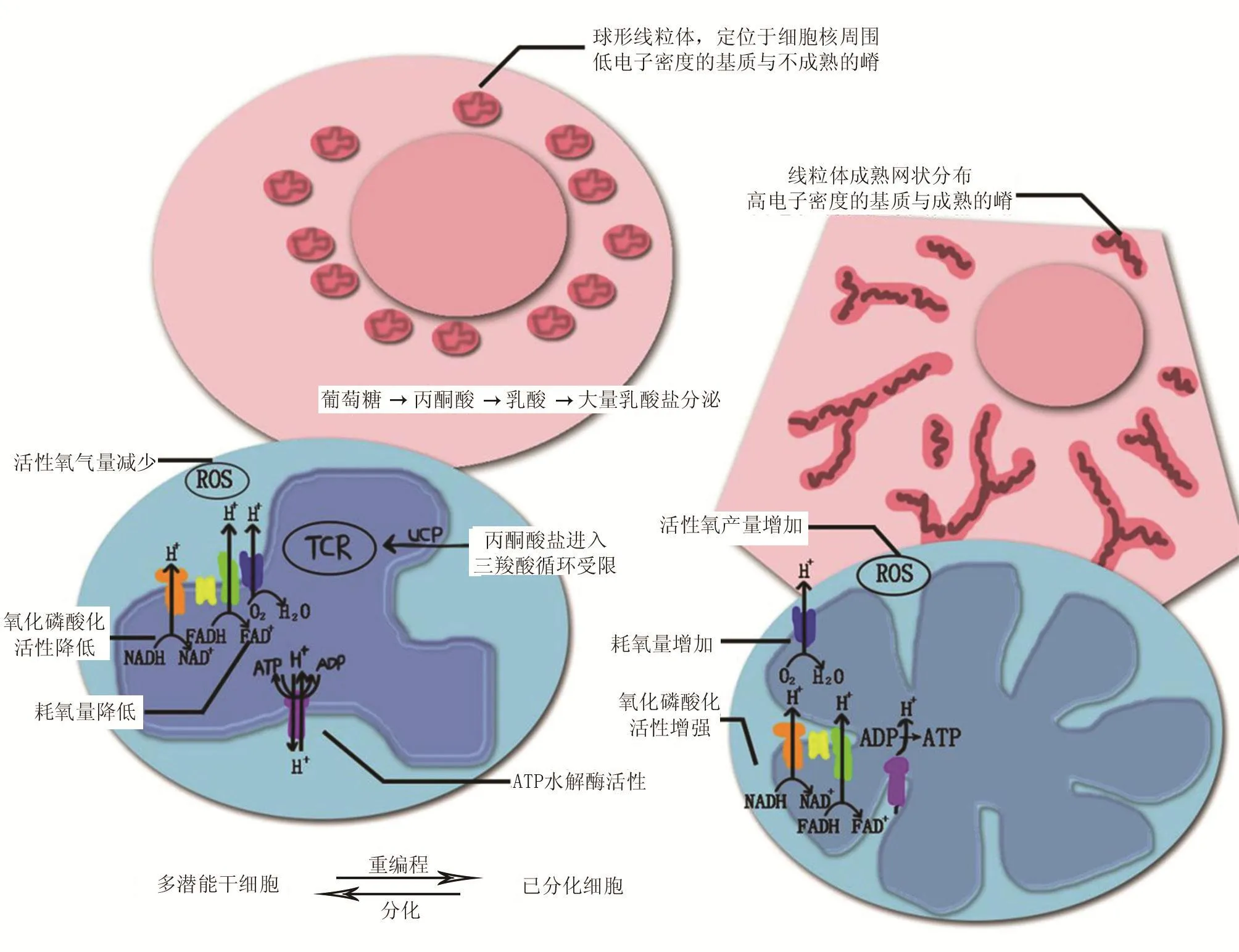
图1 多能干细胞与成体细胞线粒体能量产生过程比较[8]

表1 细胞类型与线粒体形态和代谢方式差异
由此可见,线粒体的形态、结构、功能都与细胞维持多潜能性密切相关。因此,线粒体特征鉴定也已经作为判定干细胞的干性指标。表1中列举了不同类型细胞中线粒体形态和代谢方式差异。
二、干性维持
当线粒体维持在多潜能性阶段时,它具有与成体细胞不同的成熟水平和代谢偏好,所以,有可能通过作用线粒体影响细胞的干性维持。比如,抑制线粒体功能增强干性、抑制分化。有实验表明,用一定浓度的T-2毒素作用于小鼠胚胎干细胞(mouse embryonic stem cells,mESCs),可见线粒体形态、结构和功能均受到抑制,与此同时,mESCs的分化也受到抑制,维持在干性状态[23]。近期,Shen等人发现,用一定浓度的甲基苯丙胺作用于间充质干细胞(mesenchymal stem cells,MSCs),会导致线粒体功能紊乱,线粒体生物发生及动态调节功能降低,抑制MSCs的成骨分化过程[10]。
除此之外,通过抑制过氧化物酶体增殖物活化受体γ共激活剂-1α(peroxisome proliferator-activated receptor gamma coactivator -1α,PGC-1α)[24-25]或线粒体转录因子 A,阻碍线粒体生物发生,减弱了MSCs向脂肪细胞分化[26],在用鱼藤酮抑制线粒体呼吸作用的实验中也出现了与上述类似的结果[24,27]。
三、细胞分化过程中线粒体的动态变化与调节
线粒体的动态变化影响了大多数依赖线粒体的生物过程,比如细胞凋亡、钙平衡和ATP的合成。有研究表明,线粒体的动态变化与调节作用对细胞分化过程也有显著影响。刺激线粒体生物发生降低了多能性,且有利于细胞分化或者是形成特定的细胞谱系。
(一)不同种类细胞分化过程中线粒体的动态调节
1.多潜能干细胞分化:在mESCs中,敲除线粒体蛋白如肝再生扩展因子的基因,其多潜能性标志物,如NANOG,OCT4,SSEA的表达出现一定程度的降低。研究发现[28],抑制一种与线粒体分裂相关的GTP酶,即动力相关蛋白1(dynamin-related protein-1,Drp1)活性可修复肝再生扩展因子的基因表达缺失所引起的线粒体结构、功能失常,同时可恢复多潜能性基因的表达。这表明,除了多能性干细胞的糖酵解代谢,线粒体动态平衡与适合的网络结构,对维持多能性至关重要。
在多潜能干细胞的相关研究中,一些研究者选用呼吸链抑制剂,例如氯苯腙,抗霉素A,或鱼藤酮阻扰其分化,发现这不仅造成线粒体呼吸链的抑制,同时还引发细胞毒性或其它副作用[29]。研究发现,氯苯腙诱导线粒体解耦联导致了细胞内ATP水平的下降并且降低了细胞的增殖速率。在mESCs中使用复合体Ⅲ的抑制剂抗霉素A会引起AMP/ATP比值升高,这表明在mESCs中,至少会部分依赖OXPHOS进行ATP的合成[30-31]。
用S-亚硝基-N-乙酰-DL-青霉胺刺激ESCs中线粒体的生物发生则可降低多能性标记物的表达,且促进了mESCs向肝实质细胞样分化,以及mESCs和hESCs向心肌细胞样分化。在小鼠iPSCs中过表达线粒体发生主要的调控因子PGC-1α可促进了它们向脂肪细胞的分化[24]。
2.脂肪细胞分化:在人骨髓来源的间充质干细胞向脂肪细胞分化的过程中,研究发现PGC-1α mRNA,线粒体质量和线粒体DNA拷贝数的增加,以及OXPHOS复合物的形成和OXPHOS活性的增加。通过抑制PGC-1α或线粒体转录因子A阻止线粒体生物发生减弱了成脂分化,通过鱼藤酮诱导抑制线粒体呼吸,也出现类似的结果[24]。Sireewan等[32]在Interruptin B促脂肪干细胞成脂分化的实验中发现,一定量的Interruptin B,可通过诱导PPAR-α和PPAR-γ的mRNA水平,增强脂肪干细胞的成脂分化,同时还可以大幅增加线粒体数量,上调线粒体膜电位水平。除此之外,他们发现Interruptin B可上调棕色脂肪相关蛋白的mRNA水平,能够对棕色脂肪干细胞的分化起到促进作用。抗增殖蛋白(prohibitin,PHB)是维持线粒体功能所必需的结构蛋白,其存在于白色脂肪组织脉管丰富的部位。有实验表明,PHB可以抑制小鼠3T3-L1 前脂肪细胞的分化和增殖,调节脂肪细胞的凋亡。如果用siRNA沉默PHB,发现3T3-L1前脂肪细胞分化受到抑制,线粒体网状结构破坏,线粒体嵴消失,线粒体呼吸链复合体Ⅰ活性受损及活性氧的过度生成。在促进脂肪干细胞分化的条件下,同样发现细胞中PHB的mRNA及蛋白水平明显升高[33]。
3.骨骼肌细胞分化:在骨骼肌细胞分化为肌小管的过程中,伴随着线粒体代谢的改变和自噬的调节。Fortini等[28]提出,成肌分化过程中,线粒体活动和功能的增强与自噬过程相伴随。在P53缺陷的小鼠成肌过程中,发现其新陈代谢过程糖酵解占优,线粒体生物发生受损,成肌细胞难以分化为肌小管,同时伴随自噬过程减少。由此证明,自体吞噬,线粒体稳态和细胞分化与肌肉生长发育过程相辅相成。
4.心肌细胞分化:线粒体成熟在心肌细胞分化过程中也是至关重要的。其中,线粒体通透性过渡孔(mitochondrial permeablity transition pore,mPTP)的关闭,是线粒体成熟的特征,这已被广泛认为是心肌细胞分化的关键标志。Hom等[34]发现,在早期胚胎心肌细胞中,线粒体结构和功能尚未成熟,表现为开放的mPTP及呈解偶联状态的线粒体呼吸链;而正常心肌细胞中mPTP则处于关闭状态。因此,可利用化学或基因方式关闭mPTP使得线粒体成熟,增加心肌细胞分化。人含Ⅲ型纤连蛋白域蛋白5在高耗能的心肌、脑和肌肉组织中高表达。敲除mESCs中的人含Ⅲ型纤连蛋白域蛋白5降低mESCs向心肌细胞分化的能力[25],降低分化心肌中线粒体的膜电位、ATP的产生。
线粒体的动态变化、融合分裂都与细胞分化有关;细胞多能性可影响线粒体动态变化、活性,由此可见,线粒体的动态变化与干细胞的分化相辅相成。
(二)线粒体动态调节的分子机制
有证据表明,线粒体的动态调节是通过特定的信号通路影响干细胞增殖和分化过程[35]。在线粒体分裂、融合过程中,其延伸需要线粒体内外膜的融合。动力蛋白相关GTP酶,线粒体融合蛋白1和2调节线粒体外膜的融合,OPA1蛋白和线粒体融合蛋白1调节内膜融合。一些其他蛋白也参与线粒体融合,如PHB[36]。胞浆内Drp1调节线粒体分裂,通过受体招募线粒体外膜因子,包括线粒体分裂因子、49KDa及51KDa线粒体动态蛋白,和线粒体分裂蛋白1,最终造成线粒体收缩、分裂[37]。以下将着重阐述其中部分分子在线粒体动态调节过程中的作用。
1.Drp 1:Drp1蛋白是调节线粒体分裂的关键蛋白,在线粒体分裂的过程中,首先受线粒体膜电位Fisl的招募,Drp1转位至线粒体外膜,并且富集于线粒体潜在的分裂位点。在此处,多个Drp1分子围绕线粒体形成指环结构,并通过水解GTP改变分子间的距离或角度,逐渐压缩至线粒体断裂,产生2个独立的线粒体。之后Drp1重新回到胞浆,如此循环往复[38-39]。
Wang等[40]的研究表明,敲除Drp1会影响ESCs的晚期分化,这一点在神经发生过程中尤为明显,同时,也会在ESCs中期分化过程中延迟降低多潜能性的标志物(NANOG,OCT4)水平。Wang等发现Drp1在干细胞增殖过程中,对于线粒体生物形成影响不大,但在干细胞分化、神经发生时会下调多潜能相关基因。
Hoque等[10]在研究多潜能干细胞定向诱导为心脏中胚层细胞时,同样发现Drp1会影响线粒体的有氧呼吸及糖酵解过程。研究表明,抑制Drp1,线粒体形态趋向融合,促进多潜能干细胞向心脏中胚层细胞分化,同时促进线粒体从糖酵解到OXPHOS的转化。
2.REX1:Reduced expression 1(REX1)是一种广泛应用的多能性标志。最近发现,它可通过影响线粒体动态变化来调节细胞命运。抑制REX1在hPSCs中的表达导致多能性标志的下调,所有三胚层分化标志物的上调,以及线粒体形态和活性的发育。进一步研究证实,REX1对周期素B1/ B2 具有正向调节作用,这导致了周期素B-周期素依赖激酶1对 Drp1Ser616位点磷酸化的增加,这是一种翻译后的修饰触发线粒体分裂。因此,过表达REX1或者DRP1-S616D突变蛋白更易于重编程成iPSCs,而抑制REX1或DRP1的表达阻止了重编程过程[41]。
3.PHB2:PHB是一种由PHB基因编码、高度保守的蛋白,广泛分布于多种生物细胞之中,因其具有明显的抗细胞增殖作用而得名[42]。在正常细胞中PHB可以抑制细胞周期转换和DNA的合成,它的突变可能导致细胞无限增殖。最近发现它也干细胞命运控制相关。
正常情况下,PHB2在mESCs中含量较高。当细胞过表达PHB2时,有利于mESCs的增殖,但同时抑制了它们向神经和内胚层细胞系的分化。在mESCs分化过程中,Kowno等[43]还观察到PHB2的过表达也会抑制线粒体成熟。PHB2对线粒体成熟的影响是由增强OPA1加工介导的,这导致了线粒体分裂增加和功能紊乱。
4.非编码RNA:非编码RNA是指各种不翻译成蛋白质的 RNA 分子,包括rRNA,tRNA,snRNA,snoRNA 和microRNA 等多种已知功能的RNA,还包括其他未知功能的RNA。其中,部分非编码RNA在干细胞分化、代谢调节等过程中起到重要作用。有文献报道[44],一种在肌源性分化过程中诱导产生的microRNA,miR-1,可通过共同调节线粒体生物发生和肌源性有关的转录影响肌源性程序的进行。
由于线粒体在诱导和维持干细胞的多能性、细胞分化等方面发挥了重要作用,目前已引起人们的广泛关注,但尚未全面了解线粒体调控其生理功能的作用及机制。例如线粒体在调控分化过程中的作用机制、线粒体的结构和功能在干细胞多潜能状态和分化期间变化的因果关系、与干细胞线粒体生物合成、结构及功能变化相关的表观修饰等。详细了解线粒体在以上过程中的作用及调控机制,不仅可以为揭示线粒体与干细胞干性维持、分化的关系提供重要理论基础,同时也能加深对疾病病理的理解,促进再生医学的发展。
