Huangqinflavonoid extraction for spinal cord injury in a rat model
2018-10-22QianZhangLiXinZhangJingAnLiangYanCuiCuiLiuJingJingZhaoHaoYang
Qian Zhang , Li-Xin Zhang, Jing An Liang Yan, Cui-Cui Liu Jing-Jing Zhao Hao Yang
1 Translational Medicine Center, Hong Hui Hospital, Xi’an Jiaotong University, Xi’an, Shaanxi Province, China
2 Department of Clinical Pharmacy, Hong Hui Hospital, Xi’an Jiaotong University, Xi’an, Shaanxi Province, China
3 Department of Spine Surgery, Hong Hui Hospital, Xi’an Jiaotong University, Xi’an, Shaanxi Province, China
Abstract Flavonoids from Huangqin (dried roots of Scutellaria baicalensis Georgi) have anti-inflammatory effects, and are considered useful for treatment of spinal cord injury. To verify this hypothesis, the T9–10 spinal cord segments of rats were damaged using Allen’s method to establish a rat spinal cord injury model. Before model establishment, Huangqinflavonoid extraction (12.5 g/kg) was administered intragastrically for 1 week until 28 days after model establishment. Methylprednisolone (30 mg/kg) was injected into the tail vein at 30 minutes after model establishment as a positive control. Basso, Beattie, and Bresnahan locomotor scale scores were used to assess hind limb motor function. Hematoxylin-eosin staining was used to detect pathological changes in the injured spinal cord. Immunofluorescence and western blot assays were performed to measure immunoreactivity and expression levels of brain-derived neurotrophic factor, neuronal marker neurofilament protein, microglial marker CD11b and astrocyte marker glialfibrillary acidic protein in the injured spinal cord. Huangqinflavonoid extraction markedly reduced spinal cord hematoma, inflammatory cell infiltration and cavities and scars, and increased the Basso, Beattie, and Bresnahan locomotor scale scores; these effects were identical to those of methylprednisolone. Huangqinflavonoid extraction also increased immunoreactivity and expression levels of brain-derived neurotrophic factor and neurofilament protein, and reduced immunoreactivity and expression levels of CD11b and glialfibrillary acidic protein, in the injured spinal cord. Overall, these data suggest that Huangqinflavonoid extraction can promote recovery of spinal cord injury by inducing brain-derived neurotrophic factor and neurofilament protein expression, reducing microglia activation and regulating reactive astrocytes.
Key Words: nerve regeneration; spinal cord injury;flavonoids; Scutellaria baicalensis; neurofilament; brain-derived neurotrophic factor;astrocytes; microglia; neural regeneration
Introduction
Spinal cord injury (SCI) is caused by a severe insult or progressive neurodegeneration, and frequently leads to neurological deficits and disabilities. The prevalence of SCI ranges from 250 to 906 cases per million, with an annual incidence of acute SCI of 8–49.1 per million worldwide,while the global prevalence is expected to increase over the next few years (Singh et al., 2014). Although there have been significant advances in the treatment of SCI, many patients still suffer substantial neurological disabilities (Michailidou et al., 2014). The major obstacle of SCI therapy is neuronal survival and axonal regeneration, which are usually inhibited by multiple chemical and physical factors in the damaged microenvironment (Colón and Miranda, 2016; Hachem et al., 2017). Thus, treatments are required to eliminate inhibitory factors, but enhance neuronal survival, after SCI.
Scutellaria baicalensis (Chinese skullcap) is the dry root of Scutellaria baicalensis Georgi (Lamiaceae) (see http://www.theplantlist.org). Scutellaria baicalensis is one of the most commonly used traditional Chinese herbs, and it is typically referred to as Huangqin. Huangqin is widely distributed in China, Russia, Mongolia, Korea, and Japan, and is officially listed in the Chinese Pharmacopoeia. The major phytochemicals of Huangqin have been confirmed asflavonoids, which possess anti-inflammatory, anti-tumor, anti-apoptosis, free radical scavenging, and anti-human immunodeficiency virus activities (Li et al., 2011; Gaire et al., 2014). Recently, the neuroprotective effects of Huangqin extraction and its isolatedflavonoids have also been examined in various in vivo and in vitro models, suggesting a protective action against neurodegenerative diseases (Gasiorowski et al., 2011; Cheng et al., 2012; Xu et al., 2015).
The aim of the present study was to investigate the neuroprotective effect of the Huangqin flavonoid extraction in terms of recovery of motor function and histomorphological changes following SCI in rats. The therapeutic effects of Huangqin flavonoid extraction were compared with the positive control drug methylprednisolone (MP), a routine clinical therapy for early stage SCI. We also used immunofl uorescence and western blot to evaluate changes in axonal regeneration, brain-derived neurotrophic factor (BDNF), activation of microglia and proliferation of reactive astrocytes.
Materials and Methods
Animals
Animals used in this study were maintained in accordance with the Guide for Care and Use of Laboratory Animals published by the U.S. National Institutes of Health (NIH publication No. 85-23, revised 1996). The experiments were approved by the Animal Ethics Committee of College of Medicine, Xi’an Jiaotong University, China. The 48 female specific-pathogen-free Sprague-Dawley rats aged 12 weeks and weighing 200± 20 g were provided by College of Medicine, Xi’an Jiaotong University, China (license No. SCXK [Shaanxi] 2007-001).The rats were housed at 22 ± 2°C in a 12-hour dark/light cycle,and allowed free access to food and water. Rats were randomly divided into four groups (n = 12 per group): normal, model(SCI), MP (positive control; SCI + MP) and Huangqin (SCI +Huangqinflavonoid extraction) groups.
Preparation and separation of Huangqinflavonoid extract
Huangqin was purchased from Sichuan Xinhehua Medicinal Materials Electuary Co., Ltd. (Sichuan, China), and the plant material was identified by Dr. Li-Xin Zhang. A voucher specimen was deposited at the Herbarium of Translational Medicine Center, Hong Hui Hospital, Xi’an Jiaotong University, China.
Huangqin flavonoid extraction was prepared from the dried roots of Scutellaria baicalensis Georgi by three rounds of reflux with 75% ethanol, with reflux times of 3 hours, 2 hours and 1 hour, respectively. All the extracted liquid was filtered, combined and concentrated under vacuum, and then stored at 4°C. Using this extraction preparation, 1 g extraction contains 1 g of dry root of Scutellaria baicalensis Georgi (Yune et al., 2009). Results of chemicalfingerprint of Huangqinflavonoid extraction are shown in Additionalfile 1. Identification and measurement of the major constituents in Huangqin (Scutellaria baicalensis Georgi) are shown in Additionalfile 2.
Drug dose determination offlavonoid extraction
In preliminary experiments, different doses (2.5, 7.5 and 12.5 g/kg) of Huangqinflavonoid extraction were administered by intragastric gavage to SCI rats. According to behavioral scores and hematoxylin and eosin staining (data not shown), there was a dose-dependent treatment effect. However, considering the viscosity of extraction and stomach volume of rats, we selected 12.5 g/kg as the optimal dosage for the present study.
SCI model establishment
Rats in model and Huangqin groups were anesthetized with pentobarbital sodium (40 mg/kg) intraperitoneally. A laminectomy was performed at the T9–10 levels, exposing the underlying spinal cord without destroying the spinal dura mater. The spine was clamped to stabilization at the T8–11 levels. A moderate contusion was induced using a weightdrop device (RWD G124-138; RWD Life Science Co., Ltd.,Shenzhen, China) by dropping a 10 g rod (2.5 mm in diameter) from a height of 50 mm onto the exposed cord. The skin and musculature were then sutured (Allen, 1911). Spastic swinging of the tail and flaccid paralysis of the posterior limb are considered signs of a successful SCI model. All SCI rats received bladder evacuation twice daily until sphincter control had recovered, and gentamicin (80,000 U in each rat, via intramuscular injection) was administered to prevent urinary tract infection. The cages were cleaned daily to prevent wound infection, and rats were provided free access to water and food.
Drug administration
Rats in the Huangqin group received intragastric gavage of the flavonoid extraction (12.5 g/kg, once daily) for 1 week before surgery, and then after SCI until sacrifice. Rats in the normal, MP, and model groups received free access to food and water. Rats in MP group were administered MP injection via the caudal vein at 30 minutes after surgery (30 mg/kg).
Basso, Beattie, and Bresnahan locomotor scale
Two researchers blinded to the groupings independently assessed the functional recovery of all 48 rats (12 rats per group) at each time point using the Basso, Beattie, and Bresnahan (BBB) locomotor scale. After SCI, the average BBB scores were recorded on days 0, 3 and 7, and then every week until the fourth week. The score was assigned from a range of 0 (flat paralysis) to 21 (normal gait) based on limb movement, gait, coordination and paw placement (Basso et al., 1995).
Tissue collecting and processing
At 3, 14 and 28 days after surgery, 12 rats in each group (6 rats for immunofluorescence staining, 6 rats for western blot assay) were sacrificed by pentobarbital sodium (40 mg/kg intraperitoneal), and the spinal cords collected.
For hematoxylin and eosin staining and immunofluorescence, rats were transcardially perfused with normal saline(100 mL) then 4% paraformaldehyde (100 mL). The spinal cords were removed and fixed in 4% paraformaldehyde overnight, and then transferred to 30% sucrose solutions for cryoprotection. Each spinal cord was cut into 20 mm lengths with the lesion site at the center. The tissues were then frozen, embedded in optimal cutting temperature compound and cut into 14-μm thick longitudinal sections with a freezing microtome (Leica, Germany).
For western blot assay, the spinal cords were freshly collected and frozen at −20°C without perfusion.
Hematoxylin and eosin staining
To examine the cavities and damage to the spinal cord, the longitudinal sections were stained with hematoxylin and eosin. The sections were stained with hematoxylin and rinsed,and placed in acid-alcohol (1% hydrochloric acid). The sections were then counterstained with eosin and rinsed, and treated with distilled water, alcohol and xylene. The sections were air-dried, cover slipped with neutral balsam and examined under a light microscope (DM1000; Leica, Wetzlar,Germany). Areas of cavities on the photographs were calculated using ImageJ software (V1.8.0; National Institutes of Health, Bethesda, MD, USA).
Immunofluorescence assay
For the immunofluorescence assay, sections were washed with phosphate buffered saline (PBS), and then exposed to bovine serum albumin solution (0.5%) for 1 hour at room temperature. Next, the sections were incubated overnight at 4°C with solution A or solution B. Solution A contained the following primary antibodies: goat anti-BDNF polyclonal antibody (1:400; Santa Cruz Biotechnology, Dallas,TX, USA) and mouse anti-neurofilament-H (NF-H; axonal marker (He and Nan, 2016)) polyclonal antibody (1:400; Cell Signaling Technology Inc., Danvers, MA, USA). Solution B contained the following primary antibodies: mouse anti-glialfibrillary acidic protein (GFAP; marker for astroglia (Yuan and He, 2013)) polyclonal antibody (1:400; Cell Signaling Technology Inc.) and rabbit anti-CD11b (microglia marker(Norden et al., 2015)) polyclonal antibody (1:400; Novus,New York, NY, USA). After washing with PBS, the sections were incubated for 1 hour with solution C or solution D at room temperature. Solution C contained the following secondary antibodies: rabbit anti-goat antibody (1:200; FITC;Boster, Wuhan, China) against BDNF, and goat anti-mouse antibody (1:200; TRITC; Boster) against NF-H. Solution D contained the following secondary antibodies: goat anti-mouse antibody (1:200; DyLight 594; Boster) against GFAP, and donkey anti-rabbit antibody (1:200; DyLight 488; Boster) against CD11b. Excess secondary antibody was washed off. Next, sections were incubated in 4′,6-diamidino-2-phenylindole for 5 minutes at room temperature, and then washed three times with PBS. Sections were coverslipped and immediately observed under afluorescence microscope (DM1000; Leica, Wetzlar, Germany).
The intensity of BDNF, NF-H, GFAP and CD11b expression was quantified by the sum of integrated optical density values obtained from the threshold pixels for all signals measured in each image. ImageJ (National Institutes of Health)was used to analyze optical densities. From each section,eight representative visualfields were chosen to calculate the mean optical density value. The integrated optical density was equal to neutral area in the immunofluorescence image× optical density.
Western blot assay
Protein was extracted from spinal cord tissue and heated with loading buffer at 95°C for 5 minutes for denaturation.A 50 mg of each protein sample was then separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis with 8% (NF-H and CD11b) or 15% (BDNF and GFAP)gels. Protein bands were electroblotted onto polyvinylidene difluoride membranes and blocked with non-fat dried milk for 1 hour. The blots were then probed with mouse anti-NF-H polyclonal antibody (1:1000; Cell Signaling Technology Inc.), goat anti-BDNF polyclonal antibody (1:1000;Santa Cruz Biotechnology), mouse anti-GFAP polyclonal antibody (1:1000; Cell Signaling Technology Inc.), rabbit anti-CD11b polyclonal antibody (1:1000; Novus) and goat anti-β-actin monoclonal antibody (1:1000; Cell Signaling Technology Inc.) overnight at room temperature. After washing with Tris buffered saline tween, sections were incubated with peroxidase anti-mouse, anti-goat and anti-rabbit IgG (1:5000) secondary antibodies for 1 hour at room temperature. Immunoreactivity was detected by enhanced chemiluminescence assay. Autoradiograms were exposed for 5–15 minutes. The micrographs were analyzed by Image Pro Plus 6 (Media Cybernetics, Sliver Spring, MD, USA). After defining a threshold for background correction, relative optical density of NF-H, BDNF, GFAP and CD11b in the grey matter was calculated.
Statistical analysis
All data are expressed as the mean ± SD. Statistical significances between groups were evaluated using statistical software (IBM SPSS Statistics version 22; IBM Co., Armonk,NY, USA) by one-way analysis of variance followed by Student-Newman-Keuls post hoc test. A P-value < 0.05 was considered statistically significant.
Results
Flavonoid extraction improves the morphology of the injured spinal cord in SCI model rats
In the normal group without SCI, the spinal cord tissue was smooth and intact (Figure 1B). During SCI model establishment, a visible edema and hemorrhage occurred within a few seconds (Figure 1A), and a visible spinal cord hematoma was observed on the spinal cord at 3 days after surgery.The hematoma then disappeared and was replaced by a clear wound at 14 and 28 days after SCI. In the MP and Huangqin groups, the hematoma severity at 3 days was similar to that of the model group, but was reduced at 14 and 28 days compared with the model group.
Flavonoid extraction improves motor function of SCI rats
The BBB scores of all rats in the model and Huangqin groups were 0 on the day of surgery (Figure 1C), which indicated complete loss of motor ability in the posterior limbs.From 14 to 28 days after SCI, the BBB scores indicated that flavonoid extraction intervention significantly ameliorated the hind limb motor disturbances (P < 0.01).
Flavonoid extraction improves the morphological features of the lesion site in the injured spinal cord
Hematoxylin and eosin staining was used to examine the morphological changes at the lesion site in SCI rats. After SCI, there was evidence of disorganization of the lesion site,inflammatory cell infiltration and formation of a cavity and scar in the model, MP and Huangqin groups (Figure 2).At 3 days after SCI, hemorrhages scattered throughout the injured area and diffuse degeneration were observed in the model, MP and Huangqin groups.
However, at 14 days after SCI, there was a marked increase in cell infiltration and structural disturbances at the injury site in the model group, which was not observed in the MP and Huangqin groups. Further, at 28 days after SCI, there was clear evidence of extended areas of fibrinoid necrosis,and cavities and scar formation, at the lesion site in the model group, which was less severe in the MP and Huangqin groups. The cavity areas in the Huangqin group and MP groups at 28 days after SCI were also significantly smaller than that of model group (P < 0.05), while there were no differences between the groups at 14 days (P > 0.05; Figure 1D).
Overall, data from the BBB scores and hematoxylin and eosin staining suggest that Huangqin flavonoid extraction showed protective effects against SCI, which were similar to that for MP. Previous studies also reported that MP can upregulate NF-H and BDNF expression, while down-regulate GFAP and CD11b expression, after SCI (Donnelly and Popovich, 2008). Thus, immunofluorescence and western blot data of the MP group were not shown in the present study.
Flavonoid extraction increases neurofilament expression at the lesion site of SCI rats
In the normal group, there were low levels of NF-H detected in axons and cell bodies of neurons. By contrast, the clear funicular structure of axons disappeared after SCI, with evidence of severe diffuse axonal injury, which was particularly high at 3 and 14 days after SCI (Figure 3A). In model group, there was a progressive increase in NF-H staining up to 2 weeks, while NF-H staining was significantly lower at 4 weeks recovery (P < 0.01; Figure 3B). In the Huangqin group, there was less evidence of changes in the funicular structure of the axons, and NF-H expression was significantly higher than that in the model group at all time points (P <0.05, P < 0.01; Figure 3B).
Results of western blot assay showed a significant increase in NF-H (200 kDa) expression in the model group (P < 0.01)compared with controls, which was further increased in the Huangqin group at all time points (P < 0.01; Figure 3C and D).
Flavonoid extraction increases BDNF expression at the lesion site of SCI rats
BDNF is an important neurotrophic factor, and is co-expressed with NF-H in neurons and axons (Hodgetts and Harvey, 2017). As shown in Figure 3B, BDNF expression was significantly reduced in the model group compared with the normal group (P < 0.01). Flavonoid extraction was associated with a significant increase in BNDF content in the Huangqin group compared with the model group (P <0.01).
By western blot, there was a significant decrease in BDNF expression at all time points in the model group compared with controls (P < 0.01; Figure 3C). By contrast, BDNF expression was significantly increased in the Huangqin group compared with the model group at all time points (P < 0.01).
Flavonoid extraction increases microglial marker expression at the lesion site of SCI rats
Microglia labeled by CD11b are activated after injury and distributed closely around the injury site (Smith et al., 2012).There was a significant increase in CD11b staining in the model group compared with controls, which peaked at 3 days recovery, and then progressively decreased until 28 days (P < 0.01; Figure 4B). In the Huangqin group, CD11b staining was significantly less than that in the model group at 3 days after injury, but remained significantly higher than controls at 28 days (P < 0.01). Western blot data for CD11b expression showed a similar pattern (Figure 4D).
Flavonoid extraction exerts bi-directional regulation of GFAP expression at the lesion site of SCI rats
GFAP expression in astrocytes is indicative of glial scar formation (Oberheim et al., 2009). Immunofluorescence studies showed a significant increase in GFAP staining surrounding the injury epicenter at 28 days after SCI in the model group (P < 0.01; Figure 4B). Western blot assay results confirmed thesefindings at 28 days (P < 0.01; Figure 4D). Interestingly, in the Huangqin group there was a significant increase in GFAP expression at 3 days after SCI compared with the model group, but which was significantly lower at 28 days after SCI (P < 0.05, P < 0.01, respectively;Figure 4). Thus, flavonoid extraction initially upregulated GFAP expression, and then down-regulated GFAP expression a few weeks later. These data suggest that flavonoid extraction can exert bi-directional regulation of GFAP expression after SCI, thereby maintaining GFAP at a relatively stable level.
Discussion

Figure 1 Effect of Huangqinflavonoid extraction on motor function of rats after spinal cord injury (SCI).

Figure 2 Effect of Huangqinflavonoid extraction on the histomorphological changes of the injured spinal cord.
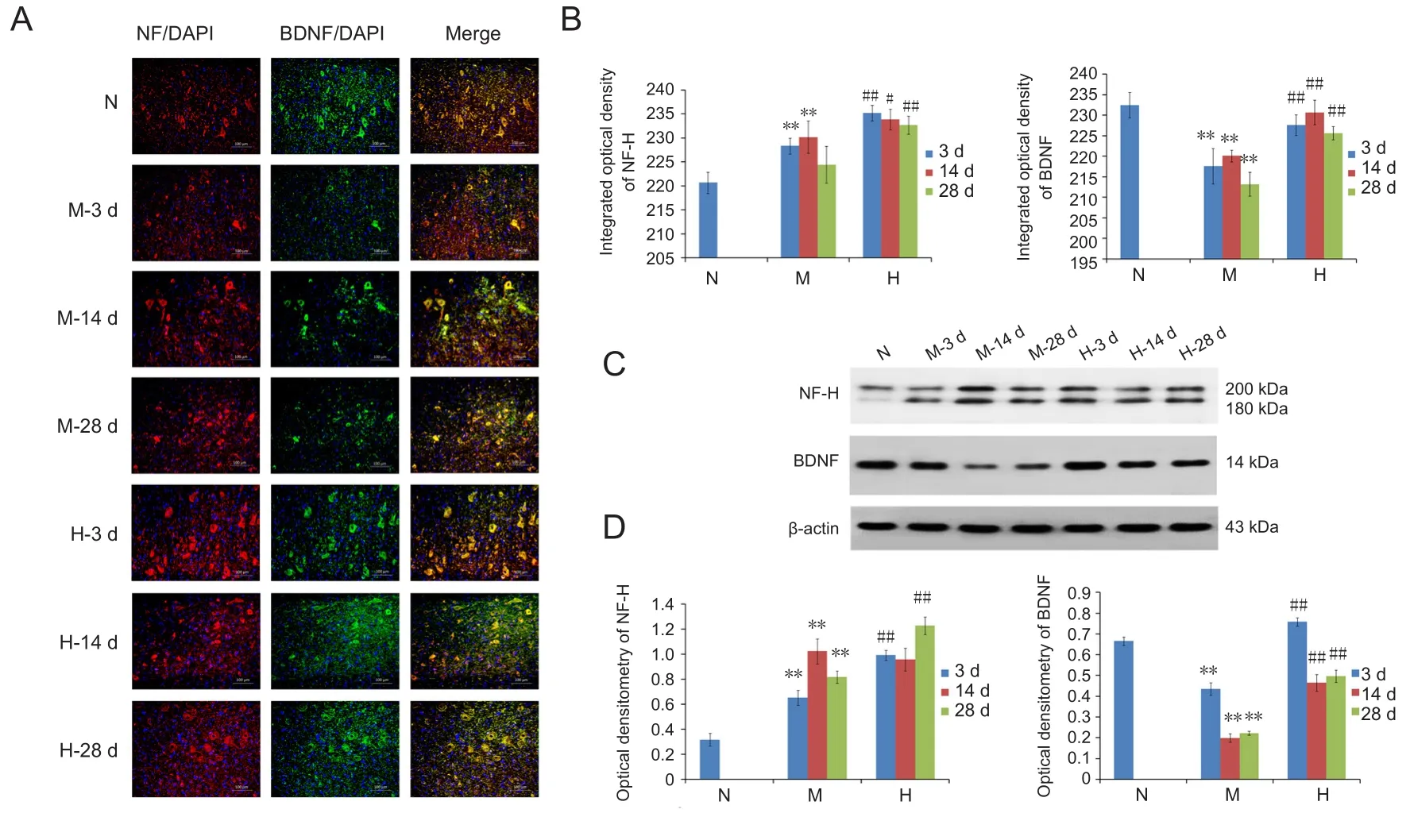
Figure 3 Effect of Huangqinflavonoid extraction on the expression of neurofilament-H (NF-H) and brain-derived neurotrophic factor (BDNF) in neurons in the injured spinal cord of SCI rats.

Figure 4 Effect of Huangqinflavonoid extraction on the expression of glialfibrillary acidic protein (GFAP) and CD11b in astrocytes and microglia in the injured spinal cord of SCI rats.
As the active ingredients and most studied flavonoids in Huangqin, baicalein, baicalin, wogonin and wogonoside have shown neuroprotective effects in multiple neurodegeneration diseases in vitro and in vivo (Chen et al., 2015a;Ding et al., 2015; Zhou et al., 2017), and via a range of mechanisms including scavenging peroxynitrite to reduce neurotoxicity (Xu et al., 2013b), ameliorating Aβ1–42protein-related pathology and cognitive dysfunction (Chen et al., 2015a), modulating stress-responsive genes (Xu et al.,2015) and helping dorsal root ganglion neurons prevent tunicamycin induced endoplasmic reticulum stress (Chen et al., 2015b). Importantly, baicalin can pass through the blood-brain barrier and is detected in cerebrospinal fluid(Huang et al., 2008), and as such, is considered a novel and promising therapeutic agent for SCI.
Interestingly, using in vivo and in vitro experiments,Yune et al. (2009) reported that totalflavonoid extraction of Huangqin produced better recovery of motor function after SCI compared with any specificflavonoid isolated from the herb. It should be noted that other flavonoids than baicalein, baicalin, wogonin and wogonoside, and other chemical components, may also provide synergistic neuroprotective actions (Wang et al., 2013; Gaire et al., 2014). As such, we utilized theflavonoid extraction for our study, rather than an individualflavonoid.
MP is a routine clinical therapy used in combination with surgery in the early stage of SCI (Bowers et al., 2016; Liu et al., 2017). Although its use remains controversial because of side effects, MP can inhibit lipid peroxidation, maintain the blood-spinal cord barrier, enhance spinal cord blowflow, inhibit endorphin release and limit the inflammatory response (Suberviola et al., 2008; Evaniew et al., 2015; Lu et al., 2016). In the present study, the Huangqinflavonoid extraction produced a similar in vivo effect on the morphology of spinal cord tissues, the BBB score and hematoxylin and eosin pathology to that observed with MP. However, further studies are required to determine the specific mechanisms of action.
Regulation of axonal regeneration
NF-H is an accepted marker of mature neuronal cells in the central nervous system. Under normal conditions, NF-H content in the cell body is low (Yang et al., 2018). After SCI,nerve cells in the lesion area are directly destroyed by the trauma, and show degeneration necrosis, structural damage,cell disruption and loss of responsiveness. However, neurons adjacent to the lesion area begin to synthesize NF-H,which is deposited in cell bodies (He and Nan, 2016; Yang et al., 2018).
Interestingly, the number of NF-H-positive neurons and the degree of NF-H staining on neuron cell bodies was strongly associated with functional recovery in the posterior limb(Wang et al., 2014; He and Nan, 2016). In the present study,we found a marked increase in NF-H levels in neuronal cell bodies after SCI, likely as a response to promote axonal regeneration. Further, administration of flavonoid extraction was associated with further upregulation the NF-H levels in the lesion area, and recovery of locomotor function, in SCI rats.
Regulation of BDNF content
BDNF is the most important neurotrophic factor in experimental SCI, and can promote neurogenesis, neuroprotection, axonal sprouting, myelination, adaptive synaptic plasticity and synaptic transmission (Kovalchuk et al., 2004;Hodgetts and Harvey, 2017). As such, a number of studies have shown that addition of exogenous BDNF, such as with gene delivery and transplant combined with cellular grafts,can promote motor recovery after SCI (Brock et al., 2010;Zhao et al., 2013; Ollivier-Lanvin et al., 2015).
In the present study, BDNF expression was decreased after SCI, as reported (Ying et al., 2008; Jung et al., 2016). Further,administration offlavonoid extraction was associated with a significant increase in BDNF expression, and an improvement in the BBB score and hematoxylin and eosin staining pathology, in the injured spinal cord.
Regulation of microglial activation
Uncontrolled microglial activation is thought to play an important pathogenic role in a range of neurodegenerative diseases. Activated microglia undergo hypertrophy and rapid proliferation, then migrate to inflammatory sites and produce neurotoxic and proinflammatory mediators that can injure neurons (Smith et al., 2012; Dursun et al., 2015; Li et al., 2016). Several studies have reported that baicalin and wogonin can reduce microglia activation in the central nervous system, via reduced expression of CD11b, suppressed phagocytosis (Xu et al., 2013a) and decreased production of proinflammatory cytokines (interleukin-6, tumor necrosis factor-α, inducible nitric oxide synthase and nitric oxide)and chemokines (Lee et al., 2003; Li et al., 2005; Xu et al.,2013a; Yeh et al., 2015). This was confirmed in vitro in primary microglia and the microglial BV2 cell line (Lee et al.,2003; Xu et al., 2013a; Yeh et al., 2015). In the present study,administration offlavonoid extraction also suppressed microglial activation, as indicated by reduced CD11b expression. However, the specific signaling pathways remain to be determined.
Proliferation of reactive astrocytes
There is increasing evidence for a role of astrocytes in blood-spinal cord barrier formation and maintenance of neuronal homeostasis (Oberheim et al., 2006; Oberheim et al., 2009). Astrocytes were also suggested to constitute an extra neuronal signaling system in the central nervous system,and play important roles in information processing during brain development, adulthood, aging and injury (Bazargani and Attwell, 2016).
In the present study, administration of flavonoid extraction increased GFAP expression at 3 days after SCI. In the primary lesion stage of SCI, the upregulation of GFAP provides beneficial support to the injured area, including maintenance of the blood-spinal cord barrier, secretion of cytokines such as interleukin-6, interleukin-10, interferon-b,tumor necrosis factor-a and monocyte chemotactic protein 1, prevention of excitotoxicity and modulation of synaptic transmission. Together with microglia, their immune responses play a ‘housekeeper’ role in the central nervous system (Radford et al., 2015; Sun et al., 2016).
In the secondary lesion stage, astrocytes become hyper-
trophic. The synthesis of a number of intermediatefilaments(e.g., GFAP and vimentin) and proteoglycans are increased,and the glial scar is formed weeks after injury. This molecular and physical barrier isolates the injured area from adjacent normal tissue, which prevents re-establishing the integrity of the central nervous system (Silver and Miller,2004; Gao et al., 2013; Yuan and He, 2013). Thus, reactive astrogliosis is considered an important therapeutic target for axonal regeneration (Zhang et al., 2007; Fitch and Silver,2008). Importantly, in the present study, administration of flavonoid extraction significantly decreased GFAP expression at 28 days after SCI, suggestive of decreased glial scar formation and removal of the physical and molecular barrier for axonal regeneration.
Conclusion
The present study provides new evidence that Huangqin flavonoid extraction can promote recovery of locomotor function following SCI in rats. Further, this effect was associated with promotion of axonal regeneration, up-regulation of BDNF, suppression of microglia and bi-directional regulation of reactive astrocytes. Future studies are required to examine the specific signaling pathways underlying these protective effects.
Author contributions:Study design, experiment implement, data analysis and paper writing: QZ; experiment implement: LXZ, JA, LY, CCL and JJZ; study supervising and paper reviewing: HY. All authors approved thefinal version of the paper.
Conflicts of interest:The authors declare no competingfinancial interests.Financial support:This study was supported by the National Natural Science Foundation of China, No. 81403278 (to QZ); the Natural Science Foundation of Shaanxi Province of China, No. 2017JM8058 (to QZ); the Shaanxi Administration of Traditional Chinese Medicine in China, No.15-ZY038 (to QZ). The funding bodies played no role in the study design,collection, analysis and interpretation of data, in the writing of the report, or in the decision to submit the paper for publication.
Institutional review board statement:The experiments were approved by the Animal Ethics Committee of College of Medicine, Xi’an Jiaotong University, China.
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement:Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-Non-Commercial-ShareAlike 4.0 License, which allows others to remix,tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewers:Mengliang Zhang, Lunds Universitet, Sweden;Syoichi Tashiro, Keio University School of Medicine, Japan.
Additionalfiles:
Additional file 1:Chemical fingerprint of Huangqin flavonoids extraction.
Additionalfile 2:Identification and measurement of major constituents in Huangqin (Scutellaria baicalensis Georgi).
Additionalfile 3:Open peer review reports 1 and 2.
杂志排行
中国神经再生研究(英文版)的其它文章
- Lithium promotes recovery of neurological function after spinal cord injury by inducing autophagy
- Analysis of transcriptome sequencing of sciatic nerves in Sprague-Dawley rats of different ages
- Exogenous brain-derived neurotrophic factor attenuates cognitive impairment induced by okadaic acid in a rat model of Alzheimer’s disease
- Partial improvement in performance of patients with severe Alzheimer’s disease at an early stage of fornix deep brain stimulation
- Epigenetic marks are modulated by gender and time of the day in the hippocampi of adolescent rats:a preliminary study
- Dysphagia in patients with isolated pontine infarction