甘露聚糖酶促进纤维素酶水解转化马尾松聚糖至可发酵单糖1
2018-10-17张雨情张玲玲吴珊珊张月美吴淑芳
张雨情, 张玲玲, 吴珊珊, 张月美, 吴淑芳*
(1. 南京林业大学 江苏省林业资源高效加工利用协同创新中心,江苏 南京 210037;2 . 南京林业大学 江苏省制浆造纸重点实验室,江苏 南京 210037)
木质纤维素原料是地球上最大量的可再生资源。过去的几十年,利用木质纤维原料制备生物乙醇或其它化学品的研究持续成为热点[1-5]。木质纤维原料不能直接转化为乙醇或化学品,需要几个单元联合处理才能实现。典型的转化工艺过程包括原料预处理、酶解和发酵三个基本单元。其中酶解是通过纤维素酶为主的糖苷水解酶作用,将经过预处理的木质纤维原料中的聚糖转化为可发酵单糖的过程,是木质纤维素转化的关键,其成本对生物乙醇等工业化生产的经济可行性有着重要影响。
与阔叶材和非木材原料相比,针叶材原料受结构形态和化学组成等影响,更加抗生物降解,酶法转化聚糖至可发酵单糖更加困难。过去的几十年,科研工作者对针叶材原料的预处理和酶解工艺已经做了大量研究。预处理方法主要包括酸水解、湿氧化、蒸汽爆破、氧脱木素、亚硫酸盐处理、绿液蒸煮、硫酸盐预处理等[6-13]。Novozymes和Genencor等酶制剂公司也对木质纤维素转化所需要纤维素酶以及酶解条件作了更加深入研究和优化,纤维素酶的价格和用量已经逐渐降低[14]。尽管这些研究在改善针叶材原料的酶水解效率和降低酶解成本方面取得了实质性进展,但仍然存在糖化速率缓慢且不完全、酶解成本过高等问题,阻碍了针叶材原料制备生物乙醇和其它化学品的工业化进程。本研究基于针叶材原料中含有一定量的甘露聚糖的化学组成特点,探讨在纤维素酶水解过程中添加甘露聚糖酶对酶解效果的影响,目的在于进一步提高针叶材原料纤维素酶水解的总糖转化率。
1 实验
1.1 材料与试剂
原料马尾松由福建青山纸业有限公司提供。经过筛选,取 15~20 mm木片用于蒸煮。复合纤维素酶CTec2由诺维信北京公司提供。甘露聚糖酶产自青霉Penicillium kloeckeri,产酶条件和过程参见文献[15]。实验所用槐豆胶粉(LBG),购于Sigma公司,其它试剂均为国产分析纯。
1.2 硫酸盐预处理
马尾松木片硫酸盐预处理在1.2 L×10的甘油浴蒸煮罐中进行。蒸煮结束后用水洗净木片后,离心脱水,平衡水分后测定水分含量,计算蒸煮得率。预处理后的木片用盘磨机(KRK,转速3 000 r/min,磨盘直径300 mm)疏解成浆,经离心脱水后装于塑料自封袋中备用。
1.3 氧脱木素
预处理后的浆料氧脱木素在自制1 L压力罐中进行,氧脱木素罐放入15 L压力电热蒸煮锅中加热,水作加热介质。氧脱木素浆料洗涤后离心脱水,塑料自封袋中备用。
1.4 打浆
打浆在PFI磨(20S7-PEI)中进行,30克绝干浆料,浆浓10%,9 000转。
1.5 酶解
称取相当于绝干1 g的预处理样品于150 mL三角瓶中,酶解底物浓度5%,柠檬酸-柠檬酸钠缓冲液浓度0.05 mol/L(pH 4.8),按照工艺要求加入一定量的CTec2、甘露聚糖酶液。将三角瓶放入空气浴摇床中,在180 r/min,50℃条件下水解48 h。酶解结束后,5 000 r/min离心15 min分离酶解液和酶解残渣。收集酶解液用于单糖组分和含量测定;残渣用去离子水离心洗涤三次,烘干称重,用于计算酶解底物失重率。
1.6 分析方法
1.6.1 酶活测定
滤纸酶活测定:参照Ghose方法[16]。
甘露聚糖酶活测定:参照文献[15]。
1.6.2 原料及预处理样品化学组分分析
将马尾松原料粉碎,取40~60 目木粉进行苯-醇抽提后用于酸水解。
苯醇抽提后的木粉原料或预处理后浆料化学组分分析参照美国能源部标准[17]。0.3克绝干样品加入3 mL 72% 硫酸,于30℃润涨1 h,期间不断搅拌。然后转入100 mL 蓝盖瓶中,加84 mL 去离子水将硫酸稀释至浓度3%,于灭菌锅(DSX-280B)内121℃下水解1.5 h。G3砂芯坩埚经恒重后用于分离残渣和水解液,收集滤液用于测定酸溶木质素含量和单糖含量;残渣测定Klason木质素含量。
酸溶木素含量测定以3% H2SO4为参比,利用紫外-可见分光光度计在波长205 nm 处测定水解液的吸光值,然后根据经验公式计算酸溶木质素的含量;残渣用热蒸馏水洗涤至中性,于恒温干燥箱中 105℃下至恒重,称重后转入马弗炉中于 575℃下灼烧至恒重,根据烘干残渣重量和灼烧残渣重量计算Klason木质素含量。
1.6.3 单糖含量测定
酸水解液单糖测定:取1 mL马尾松原料或预处理样品的酸水解液,加入40 μL 50%的NaOH调节水解液pH,以满足色谱柱pH1~3的要求,混合均匀后用0.22 μm滤膜过滤,采用高效液相色谱测定单糖组成和含量。
酶解液单糖测定:将酶解液置于沸水浴中加热 5 min 使酶蛋白沉淀,然后离心分离取上清液。稀释10 倍后用0.22 μm滤膜过滤,采用高效液相色谱分析单糖组分和含量。
酸水解液与酶水解液均采用高效液相色谱(HPLC,Agilent 1 200)分析单糖含量,分析柱为 Bio Rad Aminex HPX-87H,20 n,300 mm×7.8 mm(长×直径),保护柱为Cation-H Refill Cartridges,30 mm×4.6 mm(长×直径),检测器为RID示差检测器。进样量 10 μL,流动相为 5 mmol/L H2SO4,流速0.6 mL/min,柱温 55℃。
2 结果与讨论
2.1 甘露聚糖酶底物特异性及热稳定性
分别以槐豆胶粉(LBG,主要成分为半乳甘露聚糖)、羧甲基纤维素(CMC)、木聚糖(Xylan)和Whatman No.1滤纸(FP)为底物测定酶的活性,反应温度50℃,反应体系pH4.8,结果如表1所示。反应条件下,甘露聚糖酶对LBG酶活为7.00 U/mL,对CMC、滤纸和木聚糖也表现出少量的活性,但相对于LBG都很低,与LBG的活性比分别为0.01、0.01和0.02。
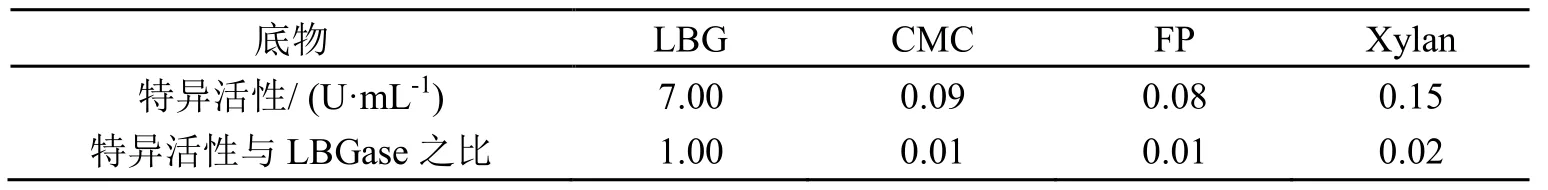
表1 P.kloeckeri甘露聚糖酶底物特异性
以LBG为底物,分别对酶的最适pH和温度进行测定,结果如图1和图2所示。由图1可见,当pH为5.5时,酶的活性最高,当pH高于或低于5.5时,酶活均随pH的增加或降低而降低,并且随pH增加降低的更加明显,pH 4.8时,活性由pH 5.5的7.6降至7.0,损失不到8%。由图2可见,当温度为50℃时 酶的活性最高,当温度45~55℃范围内,提高或降低温度使酶活稍有降低,继续升高或降低温度,则酶活降低幅度较大。图3显示的是酶在不同温度的水浴中放置不同时间后酶活的保留情况,可见,酶在温度为 25℃和 40℃时放置60分钟,保留了85%的活性;当放置的温度为55℃,则60分钟后酶活损失50%;当放置的温度升高至70℃,30分钟酶几乎完全失去活性。
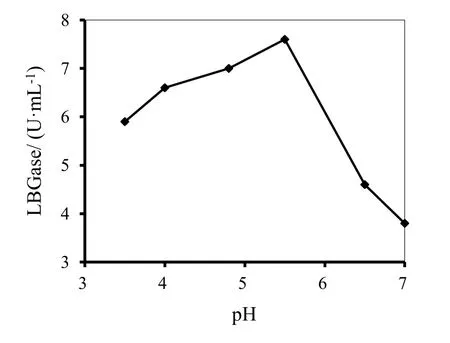
图1 pH对P.kloeckeri甘露聚糖酶活性的影响
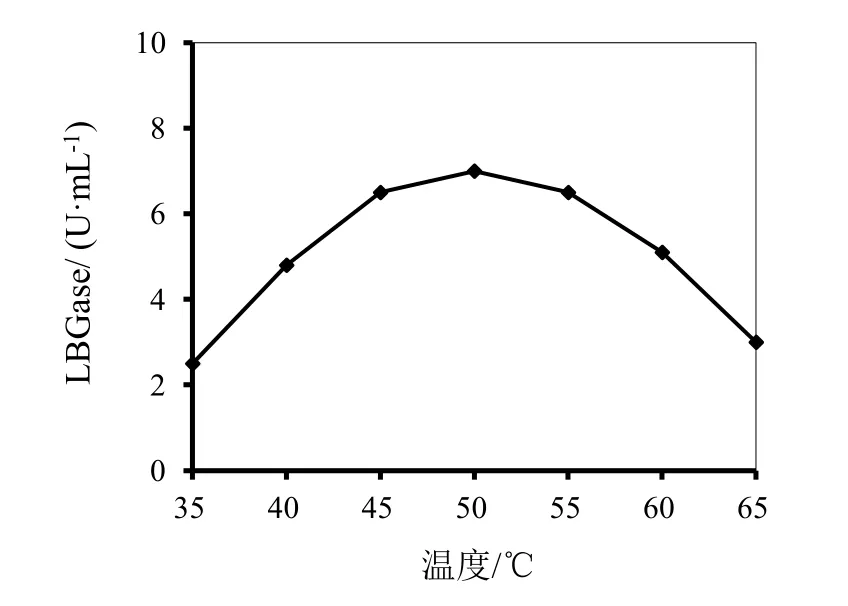
图2 温度对P.kloeckeri甘露聚糖酶活性的影响
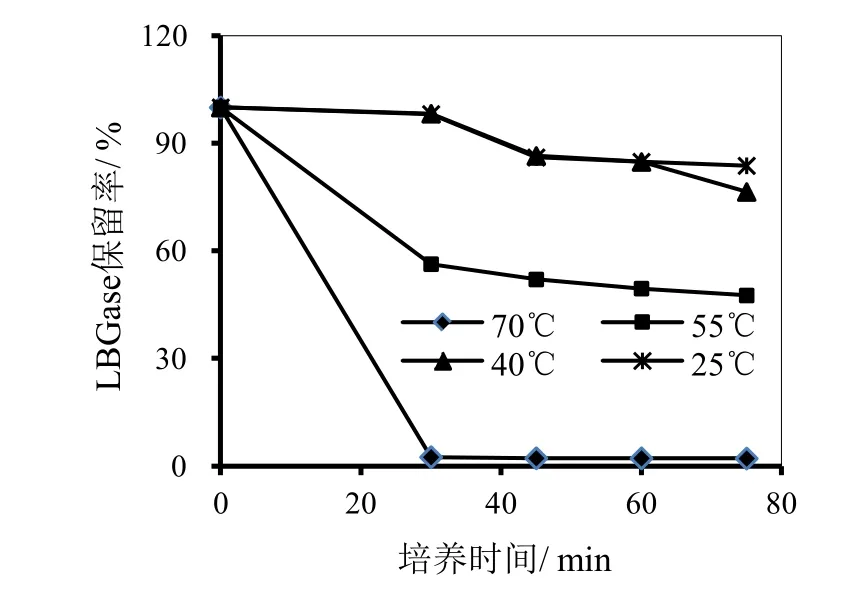
图3 P.kloeckeri 甘露聚糖酶热稳定性
根据上述测定分析结果,研究所用青霉甘露聚糖酶作用条件选择温度50℃、pH5.5为宜,pH4.8时活性略有损失,但仍保持较高活性。
2.2 马尾松木片预处理
在用碱量分别为12%、16%和20%(均以Na2O计),硫化度25%,液比1∶4,最高温度170℃、保温时间90 min条件下预处理马尾松木片,相应的预处理样品分别标记为K100、K50和K25。再分别对预处理后的样品进行氧脱木素,对应的样品分别标记为K100-O、K50-O和K25-O,预处理条件和预处理得率见表2,预处理后样品中聚糖和木质素含量列于表3。

表2 硫酸盐蒸煮和氧脱木素预处理条件及得率

表3 马尾松原料及预处理后样品的化学组成(%,对原料)
由表3可见,木片经不同条件的KP蒸煮后,木素含量分别由原料中的28.7%降至10.9%、4.1%和1.2%,相应的葡聚糖含量分别由原料中的39.0%降至38.7%,36.6%和35.1%。KP预处理过程中降解最多的是半纤维素,木聚糖和甘露聚糖之和由原料中的22.5%分别降至7.0%~5.7%,阿拉伯聚糖也由原料中的2.9%降至0.8%~0.4%。氧脱木素段,三个样品的木素含量分别由10.9%、4.1%和1.2%降至7.1%、1.9%和0.5%,由于有镁盐保护,加之条件比较缓和,氧脱木素过程中聚糖损失不多,最高不到2个百分点。
2.3 预处理马尾松浆料的酶水解
首先对马尾松KP蒸煮样品进行PFI磨打浆处理,对应的样品记为K100-R、K50-R和K25-R,然后以10 FPU/g的酶用量,用复合纤维素酶CTec2对马尾松KP蒸煮及其打浆后的样品进行酶解,同时添加甘露聚糖酶,比较甘露聚糖酶对 CTec2酶解效果的影响。以酶解总糖得率和底物失重率评价酶解效果,其中总糖得率定义为酶解得到的单糖质量总和(以聚糖计)占酶解前底物质量的百分比;底物失重率定义为酶解底物损失的质量占酶解前底物质量的百分比。
KP蒸煮样品酶解结果如图4所示。由图4可见,未经打浆处理的样品,酶解总糖得率和底物失重率都比较低,除样品K25底物失重率稍高外,其它均低于35%,并且酶解时添加甘露聚糖酶均未明显提高总糖得率,甚至稍有降低。KP预处理后打浆的样品,相同酶解条件下,总糖得率和底物失重率比未打浆样品均明显提高,并且添加甘露聚糖酶均不同程度提高了总糖得率和底物失重率,其中底物失重率提高幅度更大。尤其是样品K100-R和K50-R,当甘露聚糖酶添加量分别为5 U/g和10 U/g时,底物失重率分别由对照的44.6%和49.9%提高至49.5%、54.0%和51.0%、56.4%;而样品K25-R,对照样品酶解失重率相对较高,已达54.7%,添加甘露聚糖后提高的幅度较小,不到2个百分点。相对于底物失重率,添加甘露聚糖酶虽然也能够提高总糖得率,但是提高的幅度较小,并且甘露聚糖酶用量由5 U/g增加至10 U/g时,总糖得率没有明显提高,样品K100-R甚至稍有降低。继续增加甘露聚糖酶用量,则失重率不再提高,而总糖得率随甘露聚糖酶用量的增加而下降(数据未列出)。
KP蒸煮导致大部分甘露聚糖降解并溶出,尤其是暴露在纤维表面的甘露聚糖更少。而酶的水解是由外及内的,添加甘露糖酶不仅没有促进水解,反而因为影响纤维素酶在底物上的吸附而导致酶解效果下降。打浆后,由于纤维分丝帚化,相对较多的葡甘聚糖暴露于酶液中,更利于甘露聚糖酶的作用。因此,添加甘露聚糖酶对于打浆后的样品纤维素酶水解的促进作用更大。
对KP预处理样品K100、K50和K25分别进行氧脱木素,对应的氧脱木素样品标记为K100-O、K50-O和K25-O,对应的氧脱木素再打浆样品标记为K100-O-R、K50-O-R和K25-O-R。同样以10 FUP/g 的CTec2用量对氧脱木素及其打浆后的样品进行酶解,比较添加甘露聚糖酶对酶解效果的影响,结果如图5所示。
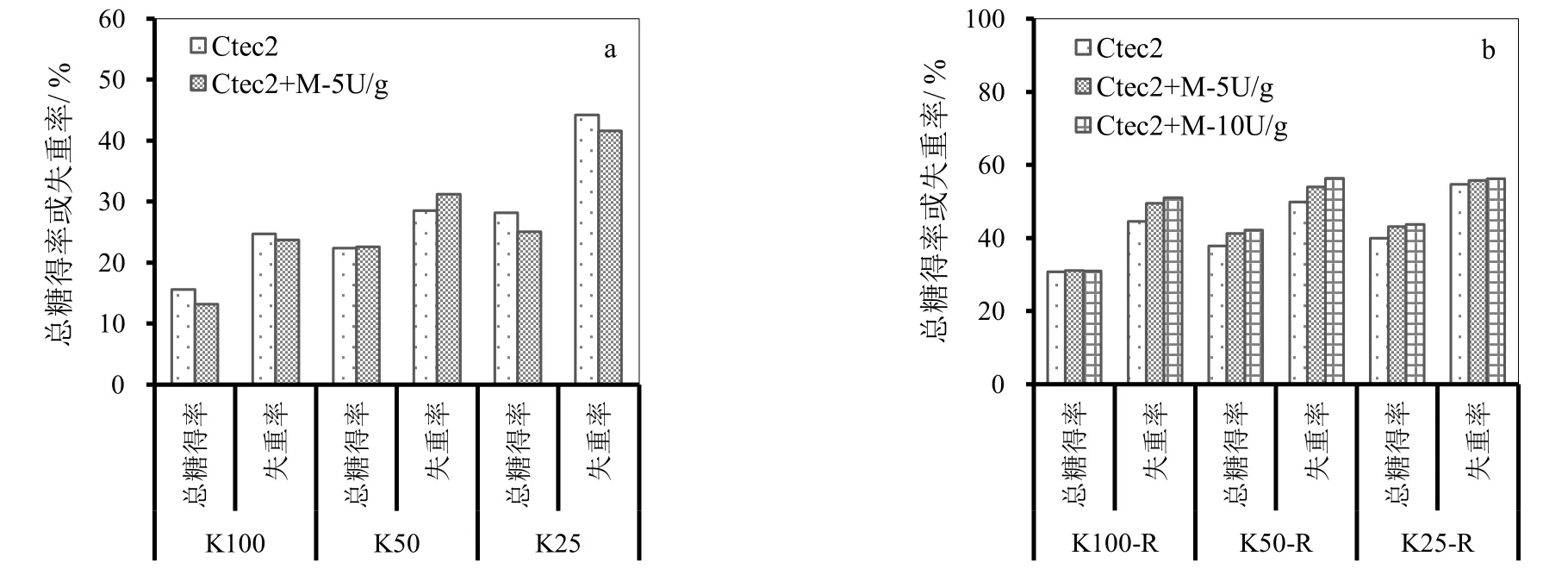
图4 KP预处理样品纤维素酶水解时添加甘露聚糖酶效果(a. KP预处理样品,b. KP-R样品)

图5 KP样品氧脱木素后添加甘露聚糖酶的酶解酶解效果(a. 未打浆,b. 打浆)
氧脱木素样品酶解总糖得率和底物失重率均比相应氧脱木素前的样品有明显提高。添加甘露聚糖酶后的酶解总糖得率,仅K25-O由对照的54.3%提高至58.1%,其它两个样品的总糖得率均没有提高甚至略有下降;但是,添加甘露聚糖酶后的底物失重率却有较大幅度提高,K50-O和 K25-O尤其明显,由对照的55.4%和48.8%提高到71.0%和68.8%,分别为15.6和20个百分点。氧脱木素后打浆的样品,相同条件下CTec2单独酶解总糖得率和底物失重率均明显高于未打浆样品。添加甘露聚糖酶后,无论总糖得率还是底物失重率也均明显高于对照样品。其中K100-O-R和K50-O-R提高幅度更大,总糖得率分别由对照的54.4%和66.0%提高至58.0%和70.6%,提高了近5个百分点。底物失重率则由对照的65.7%和76.6%提高至68.1%和82.0%,提高的幅度与总糖得率相当。
针叶材由于原料结构和化学组成等原因,原料中的聚糖通过纤维素酶水解转化至单糖非常困难,通常需要通过较强条件的预处理[18]。以K50-O-R 为例,原料在KP预处理和氧脱木素过程中,聚糖损失约22.4个百分点。预处理后的样品经10 FPU/g的纤维素酶水解,酶解底物的66%转化为可发酵单糖,而酶解失重率为76.6%,尚有约16%的聚糖在酶解条件下无法转化为单糖,可能以低聚糖或者纳米微晶的形式存在。若进一步延长酶解时间或者增加酶用量,可以得到更高的总糖得率。
为了探讨进一步提高酶解总糖得率的可能性,对于氧脱木素且打浆处理的样品,分别将CTec2用量由10 FPU/g增加至15 FPU/g,再比较添加甘露聚糖酶对酶解效果的影响,结果如图6所示。首先,对于三个不同木素含量的样品,CTec2用量增加均明显提高了酶解总糖得率和底物失重率。此外,除K25-O-R样品当CTec2用量为15 FPU时,添加甘露聚糖酶相对于未添加甘露聚糖酶降低了其酶解底物失重率和总糖得率外,其它样品无论CTec2用量为10 FPU/g还是15 FPU/g,添加甘露聚糖酶后酶解总糖得率和底物失重率均明显提高,并且CTec2用量15 FPU/g时提高的幅度更大。对于样品K50-O-R,当CTec2用量为15 FPU/g添加甘露聚糖酶后,其总糖得率和底物失重率分别由对照的83.6%和90.3%提高至89.2%和96.8%,分别提高了近7个百分点。
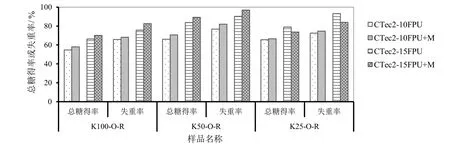
图6 KP-氧脱木素样品不同纤维素酶用量时添加甘露聚糖酶对酶解效果的影响
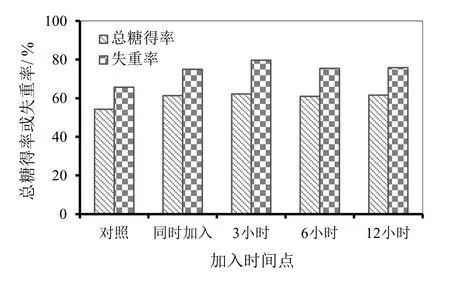
图7 甘露聚糖酶加入时间点对纤维素酶解效果的影
为了使甘露聚糖酶能够最大限度促进纤维素酶水解,以得到更高的酶解总糖得率,选取K100-O-R作为酶解底物,探讨在CTec2酶解过程中的不同时间点添加甘露聚糖酶,对其促进纤维素酶水解的影响,结果如图7所示。由图7可见,与CTec2和甘露聚糖酶同时加入相比,CTec2酶解 3小时后再添加甘露聚糖酶,其总糖得率仅提高1个百分点;CTec2酶解更长时间后添加甘露聚糖酶均没有明显提高总糖得率。但是,比较底物失重率发现,CTec2酶解3小时加入甘露聚糖酶,可使底物失重率比二者同时加入时提高约5个百分点;CTec2酶解更长时间后添加甘露聚糖酶,底物失重率则没有明显高于同时添加的情况。分析认为,CTec2与甘露聚糖酶同时添加,则甘露聚糖酶与纤维素酶竞争吸附于纤维表面时处于劣势,并随着水解时间的延长而失活。纤维素酶水解3小时左右,底物在纤维素酶的作用下部分碎片化,但仍处于水解的初始阶段,此时添加甘露聚糖酶更加利于其促进作用。
2.4 马尾松预处理样品酶解后各单糖得率及总糖转化率
为了探讨添加甘露聚糖酶促进纤维素酶水解效果的原因,对样品K50-O-R酶解液中单糖组分进行了分析,并且与纤维素酶水解时添加木聚糖酶的效果进行了比较,结果如表4所示。比较意外的是,尽管添加甘露聚糖酶后,酶解总糖含量增加,但是酶解液中木糖和甘露糖总量增加的幅度并不大,葡萄糖含量则根据CTec2 的用量,分别增加2~5个百分点,这与总糖得率的增加幅度基本一致。前面甘露聚糖酶的底物特异性研究显示,此甘露聚糖酶中几乎没有纤维素酶活性,因此,甘露聚糖酶不会直接作用于葡聚糖。针叶材中的D-吡喃式葡聚糖与D-吡喃式甘露聚糖基以β-1,4糖苷键连接成主链,葡萄糖基与甘露糖基的比例一般为1∶1.5~6.5[19]。分析认为,因预处理后的酶解底物中甘露聚糖的含量已经很少,甘露聚糖酶的作用是破坏了甘露糖之间的连接,进而促进纤维素酶的作用,因此,甘露聚糖酶的作用不仅仅是其本身对甘露聚糖的降解,更大的作用在于促进葡聚糖的转化。
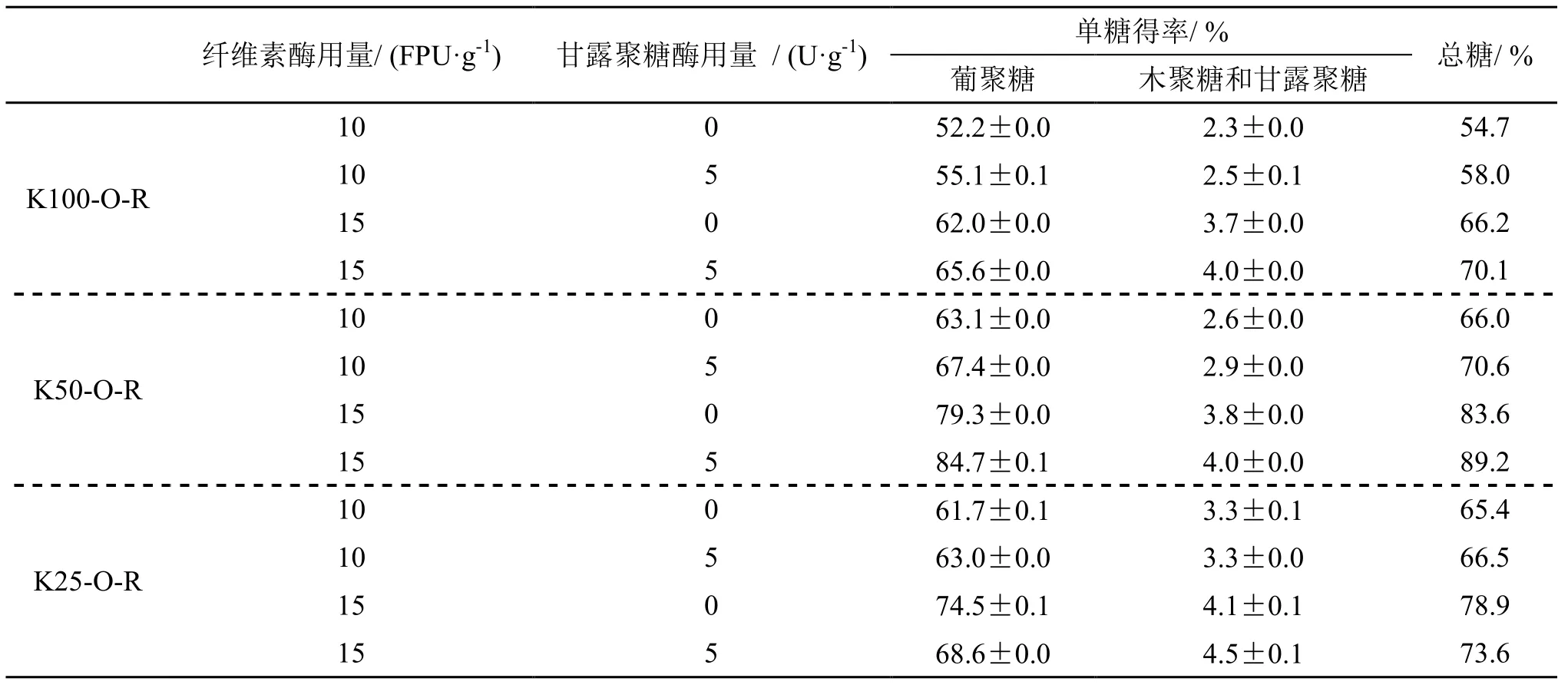
表4 马尾松预处理样品酶解后几种单糖得率及添加甘露聚糖酶的影响
上述总糖得率反映的是预处理后样品酶解的难易程度,相对而言,脱木素程度高,则酶解聚糖转化相对容易。但是,脱木素程度越高,条件越剧烈,那么聚糖在预处理过程中损失也较多。因此,对于低木素含量的样品,尽管酶解单糖得率可能比较高,但相对于原料中的聚糖,其总糖转化率却未必随之增高。这里总糖转化率为酶解得到的单糖总量(以聚糖计)占原料中相应的总聚糖的百分比。样品经不同条件的蒸煮和氧脱木素预处理,对不同样品的整个预处理及酶解过程的总糖转化效率进行评价,结果如图 8所示。由前述酶解总糖得率可知(如图6),K25-O-R无论在CTec2用量为10 FPU/ g 还是15 FPU/ g时,也无论是否添加甘露聚糖酶,其酶解总糖得率总是高于K100-O-R的相应条件的总糖得率。然而,如果计算总糖转化率,则K25-O-R则总是低于K100-O-R相应条件的总糖转化率。这是由于K25-O-R蒸煮用碱量较高,聚糖损失较多,蒸煮氧脱木素后,其总糖损失率较K100-O-R高约6个百分点。而样品K50-O-R,由于预处理条件相对比较合适,无论酶解总糖得率还是整个过程总糖转化率都相对较高。对于 K100-O-R和K50-O-R,CTec2用量为10 FPU/g和15 FPU/ g时,添加甘露聚糖酶均可提高总糖转化率 3~5个百分点,两个样品酶解总糖转化率最高分别可达 66.0%和68.1%。而添加甘露聚糖酶却不能提高样品K25-O-R的总糖转化率。这主要由于后者预处理条件比较剧烈,预处理样品中甘露聚糖保留不多所致。
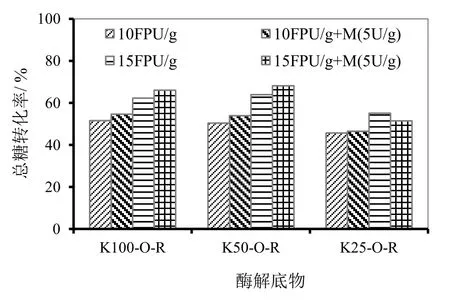
图8 添加甘露聚糖酶对酶解总糖转化率的影响
3 结论
马尾松原料经硫酸盐蒸煮联合氧脱木素及打浆处理后,采用复合纤维素酶CTec2进行酶解过程添加甘露聚糖酶可以促进酶解作用,其促进纤维素酶水解作用的效果因预处理程度和方法的不同而不同,其中打浆处理、氧脱木素处理或者二者联合处理均可提高甘露聚糖酶的作用效果。甘露聚糖酶的作用不仅仅是其本身对甘露聚糖的降解,更大的作用在于促进纤维素酶对葡聚糖的转化。当纤维素酶水解至3小时左右再添加甘露聚糖酶,则甘露聚糖酶促进纤维素酶水解的效果更好。
