芥蓝miR172家族成员进化特性比较及时空表达分析
2018-10-16李文静王杏茹陈冰星赖钟雄郭容芳
李文静,王杏茹,刘 涛,陈冰星,赖钟雄,郭容芳*
(1 福建农林大学 园艺植物生物工程研究所,福州 350002;2 福建农林大学 园艺学院,福州 350002)
自从2002年植物的microRNA(miRNA)第一次被报道之后,人们逐渐获得了大量关于植物miRNA生物合成、作用机制以及可能的生物学功能等方面的信息[1-2]。植物中的miRNA是一类长度约为20~25 nt的单链非编码小分子RNA[3]。2008年,Brodersen等[4]发现,miRNA除了通过剪切靶基因的转录产物使其沉默外,还可以通过抑制蛋白质翻译的方式使靶基因沉默。
植物miRNA通过RNase Ⅲ家族Dicer-like1结合双链RNA结合蛋白、HYPONASIIC LEAVESI蛋白和 SERRATE蛋白,将miRNA双链核苷酸从miRNA初级体(pri-miRNA)转录本中切除[2]。pri-miRNA包含半自我互补的折叠或茎环结构,即miRNA的前体(pre-miRNA),可产生成熟的miRNA[5]。miR172是较早被发现的miRNA,在植物中非常保守,其作用方式和生物学功能已经研究得较为深入。2010年,Werner等[6]对拟南芥pri-miR172a进行了广泛的结构功能分析,确认了远端折叠结构对miR172a加工的重要性。
miR172参与到植物发育的多种过程中,如开花、块茎形成、结瘤和果实发育等[7]。在模式植物拟南芥中,miR172在转录和翻译上抑制其靶基因APETALA2(AP2)作用的发挥,从而参与到花器官的形态发生;miR172还可以通过调控靶基因TARGETOFEAT1 (TOE1)、TARGETOFEAT2 (TOE2)和TARGETOFEAT3 (TOE3)控制开花时间的早晚。在拟南芥中,共发现了5个功能冗余的pre-miR172a-e,经加工后形成3个成熟的miR172a-c。目前,miR172的同源物已经在水稻、大麦、番茄、大豆、马铃薯中分离得到。通过比较基因组学的方法,十字花科植物中的miR172也从其基因组序列中进行了预测,2014年,Shivaraj等[8]的研究表明,在甘蓝中,共有4种miR172,其中包括1个miR172a、3个miR172b、2个miR172d和2个miR172e。本研究筛选分析芥蓝miRNA数据库(SRP076430、SRS1497183和SRX1837757)[9],获得了20条miR172成熟体及其前体序列,通过与其他物种中的miR172序列进行比对并分析其进化特性、结构特征,预测其靶基因,最后检测了芥蓝不同发育时期各部位器官中miR172的表达水平,初步阐明了miR172在芥蓝发育过程中的表达模式,为进一步研究其功能特征提供了基础。
1 材料和方法
1.1 材 料
本试验于2015年12月至2016年10月于福建农林大学园艺植物生物工程研究所进行,供试材料为‘福州黄花’芥蓝(Brassicaalboglabra),在2.8 m×1.1 m的畦中种植‘福州黄花’芥蓝,共6畦,每畦种植15株,选取生长势相似的植株进行取样标记,根据生长期的差异,分别对芥蓝营养生长期(根、茎、叶)和生殖生长期(根、茎、叶、成花、3.5 cm种荚及其中的种子)9个不同组织器官进行采样,采用RNAiso plus(SO1410)法提取总RNA,用TaKaRa SYBR ExScriptTM试剂盒(PK0498)反转录并进行实时荧光定量PCR,每个组织部位3次重复。
1.2 方 法
1.2.1植物miR172序列的获取与分析芥蓝miR172家族前体序列及其成熟体序列均来自于芥蓝数据库,耧斗菜(Aquilegiacaerulea)、山羊草(Aegilopstauschii)和拟南芥(Arabidopsisthaliana)等30种植物的前体与成熟体序列均下载于miRBase数据库(http://www.mirbase.org/index.shtml)。miR172a家族前体与成熟体序列分析均采用DNAMAN ver.6.0软件进行,二级结构采用mfold(21)在线软件(http://unafold.rna.albany.edu/?q=mfold)进行预测,系统发育进化树采用MEGA6.0软件进行构建。
1.2.2芥蓝miR172a靶基因预测miRNA通过其靶基因发挥生物学功能,本实验采用多个软件进行miR172靶基因的预测。首先通过在线软件(http://plantgrn.noble.org/psRNATarget/?function=3)预测芥蓝miR172a的靶基因,运行参数均为默认值。然后结合相应的过滤条件如自由能、得分值等进行过滤。
1.2.3pre-miR172a家族成员在芥蓝不同发育阶段组织中的表达分析检测芥蓝pre-miR172a家族成员在不同组织器官中的表达情况是进一步对芥蓝miR172a进行潜在功能分析的重要手段。本试验利用SYBR qPCR技术,首先利用DNAMAN ver.6.0对芥蓝pre-miR172不同成员的序列进行qPCR引物设计(表1),随后对上述取样9个组织部位反转录的材料进行实时荧光定量检测,3次重复,以Acting为内参基因,采用Graphpad软件进行数据统计与图表制作,用SPSS进行分析。
2 结果与分析
2.1 芥蓝miR172a家族成熟体序列分析
为进一步了解miR172的功能,本实验分析了芥蓝中miR172的成熟体。首先从芥蓝miRNA数据库中筛选获取了20条miR172成熟体序列,它们的序列长度为19~21 nt。利用 DNAMAN ver.6.0软件对上述20条序列进行比对(图1),发现20条序列都存在一个重叠区域,该区域中ACTAGATC 8个碱基高度保守,并且在芥蓝pre-miR172的3′和5′端均能形成成熟体。
2.2 芥蓝pre-miR172a家族二级结构预测
植物体内的活性分子均以一定形态出现,其形态结构的稳定性可能影响其功能的强弱,因此对芥蓝pre-miR172a家族进行结构预测显得非常重要。本试验采用3.5版mfold在线软件对5条芥蓝pre-miR172a序列进行结构预测(图2),结果显示,所有序列折叠后均能形成典型、复杂的发夹结构,其序列的最小折叠自由能在-54.55~-78.60 kal/mol之间,可见pre-miR172a在芥蓝体内均能自发形成稳定结构。通过其结构进一步分析发现,芥蓝pre-miR172a的最小折叠自由能在一定程度上与序列的碱基数呈负相关,即碱基数越多,最小折叠自由能的负值越大,但它同样也可能受其他因素的影响,如序列G/A比值、茎环数目及茎环大小等。
2.3 植物pre-miR172a家族系统发育进化树分析
构建系统发育进化树是对植物miR172a家族进行进化特征分析的重要手段。试验从miRBase数据库中筛选获得耧斗菜、山羊草、拟南芥等30个pre-miR172a成员,利用MEGA6.0软件对5个芥蓝pre-miR172a与上述30个序列构建进化树(图3)。由图3可知,整个进化树分为2大分支,bal-pre-miR172a-5p-2、cme-pre-miR172a、cpa-pre-miR172a、bal-pre-miR172a-1、bal-pre-miR172a-2和bal-pre-miR172a-3为其中一支,其余成员共为另一支。分析发现芥蓝miR172a成员总体分布比较集中,除bal-pre-miR172a-4外,其他4个成员:bal-pre-miR172a-5p-2、bal-pre-miR172a-1、bal-pre-miR172a-2和bal-pre-miR172a-3为一支,可见芥蓝pre-miR172a家族在进化过程中也相对保守,但从总体上分析芥蓝与番木瓜的pre-miR172a亲缘关系较近。
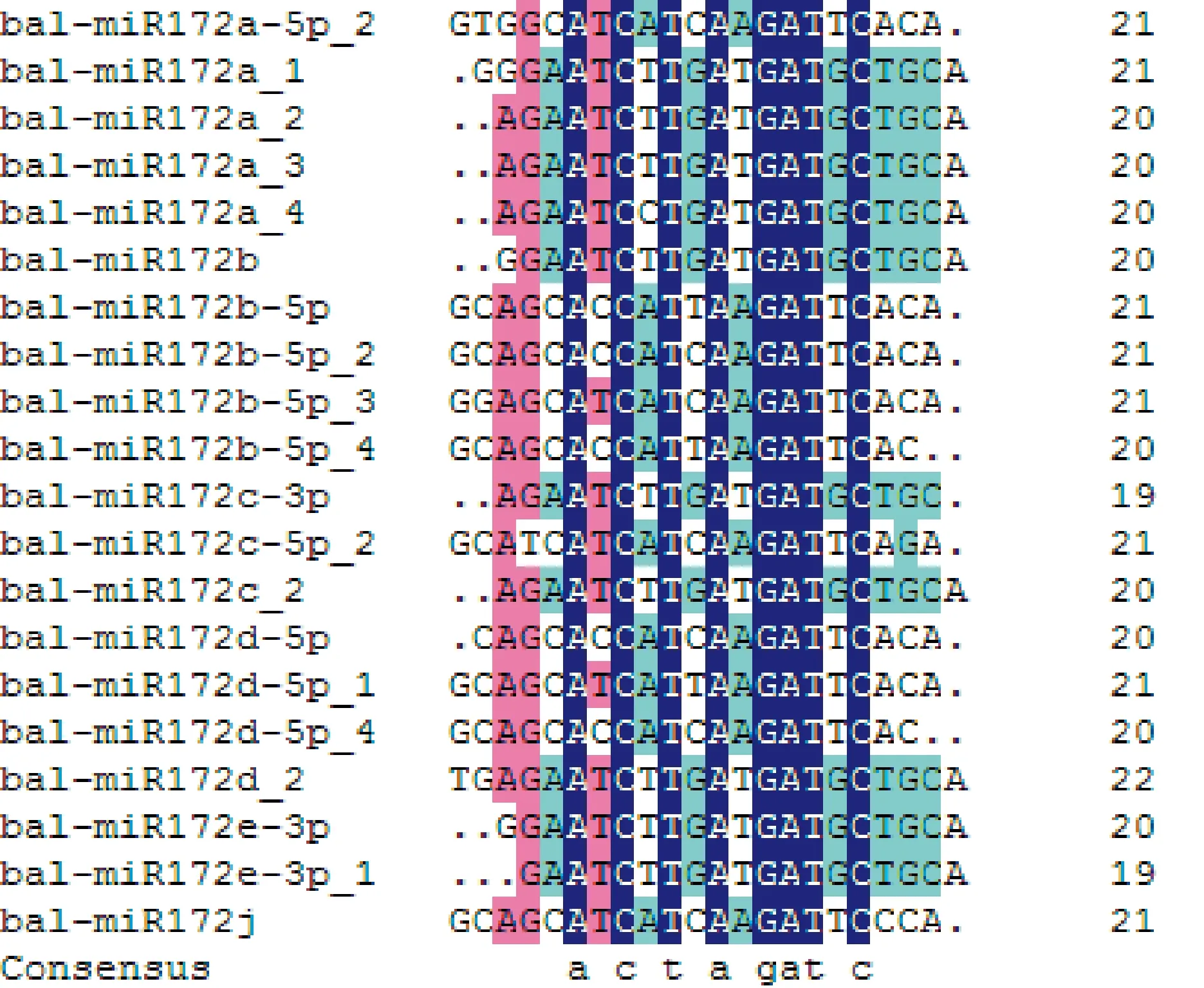
图1 芥蓝miR172序列比对分析Fig.1 Analysis of pre-miR172a sequences in B. alboglabra
2.4 植物miR172a家族成熟体系统发育进化树构建与序列分析
为进一步了解植物miR172a家族的分布与序列进化特性,试验从miRBase数据库中筛选获取30种植物,从芥蓝转录组数据中筛选出5条miR172a成熟体,发现所有成员的碱基数非常稳定,均在20~21nt间。利用MEGA6.0和DNAMAN ver.6.0软件对芥蓝、耧斗菜、山羊草等41条成熟体序列进行比对与进化树构建(图4),发现植物miR172a的41个成员被分成两支,csi-miR172a-5p、ppe-miR172a-5p、stu-miR172a-5p、aly-miR172a-5p、bal-miR172a-5p-2、ata-miR172a-5p、bdi-miR172a-5p和gra-miR172a为一支,其它29个成员为另一支,可见csi-miR172a-5p、ppe-miR172a-5p、stu-miR172a-5p、aly-miR172a-5p、bal-miR172a-5p-2、ata-miR172a-5p、bdi-miR172a-5p和gra-miR172a与其他成员的亲缘性较远,序列比对也充分验证了该观点。通过对大分支进行分析,发现有20个碱基高度重叠,再次验证了植物miR172a在进化过程中具有高度的保守性,这也暗示了植物miR172a家族成员间功能的相似性与协同性。
2.5 芥蓝miR172a家族靶基因预测分析
对芥蓝miR172a家族成员进行靶基因预测是对其生物学功能分析的间接手段。本试验采用psRNATarget在线软件对miR172a家族成员进行预测。结果显示,5个miR172a家族成员共存在13个靶标基因。表2也发现miR172a-1、miR172a-2、miR172a-3和miR172a-4这4个成员均可作用于相同靶标——AP2家族基因,再次验证了芥蓝miR172a家族在进化过程中功能上确实具有保守性和协同性。
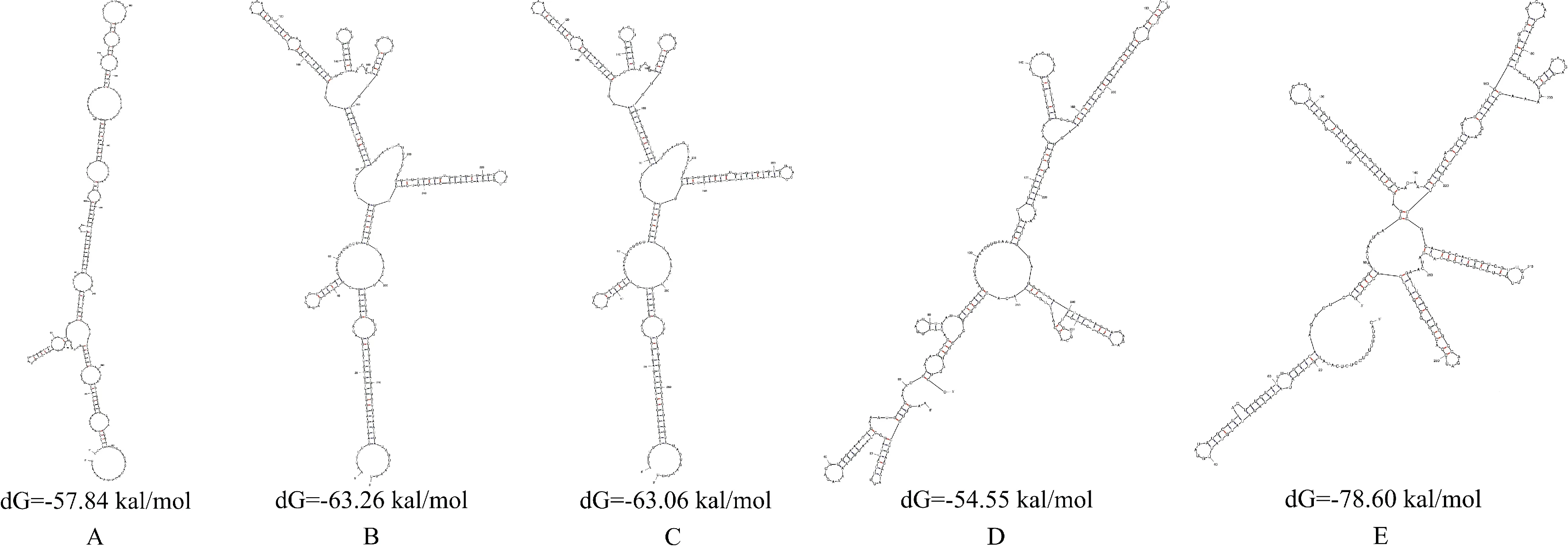
图2 芥蓝pre-miR172a家族二级茎环结构A. pre-miR172a-1;B.pre-miR172a-2;C.pre-miR172a-3;D.pre-miR172a-4;E.pre-miR172a-5p-2Fig.2 Analysis of pre-miR172a structure in B. alboglabra
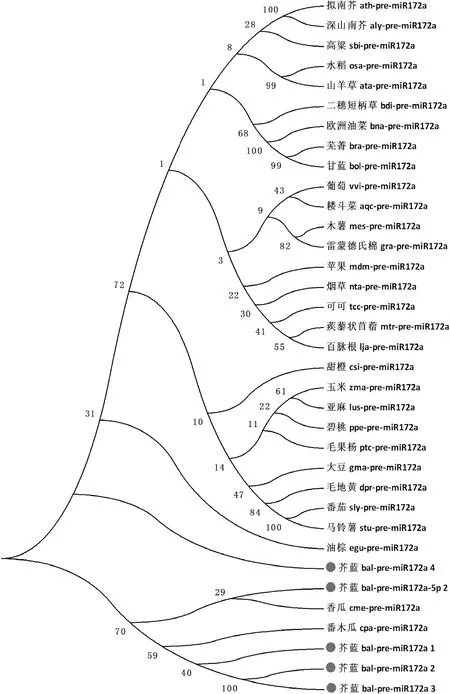
ath.拟南芥;aly.深山南芥;sbi.高粱;osa.稻;ata.山羊草;bdi.二穗短柄草;bna.欧洲油菜;bra.芜菁;bol.甘蓝;vvi.葡萄; aqc. 耧斗菜;mes.木薯;gra.雷蒙德氏棉;mdm.苹果;nta.烟草;tcc.可可;mtr.蒺藜状苜蓿;lja.百脉根;csi.甜橙;zma.玉米; lus.亚麻;ppe.碧桃;ptc.毛果杨;gma.大豆;dpr.毛地黄;sly.番茄;stu.马铃薯;egu.油棕;bal.芥蓝;cme.香瓜;cpa.番木瓜;图4同图3 利用NJ法构建植物pre-miR172a进化树ath. Arabidopsis thaliana;aly.Arabidopsis lyrata;sbi.Sorghum bicolor;osa.Oryza sativa;ata.Aegilops tauschii;bdi.Brachypodium distachyon;bna.Brassica napus;bra.Brassica rapa;bol.Brassica oleracea;vvi.Vitis vinifera; aqc. Aquilegia caerulea;mes.Manihot esculenta;gra.Gossypium raimondii;mdm.Malus domestica;nta.Nicotiana tabacum;tcc.Theobroma cacao;mtr.Medicago truncatula;lja.Lotus japonicus;csi.Citrus sinensis;zma.Zea mays; lus:Linum usitatissimum;ppe.Prunus persica;ptc.Populus trichocarpa;gma.Glycine max;dpr.Digitalis purpurea;sly.Solanum lycopersicum;stu.Solanum tuberosum;egu.Elaeis guineensis;bal.Brassica alboglabra;cme.Cucumis melo;cpa.Carica papaya; The same as Fig.4Fig.3 Phylogenetic relationships among pre-miR172a sequences using NJ method
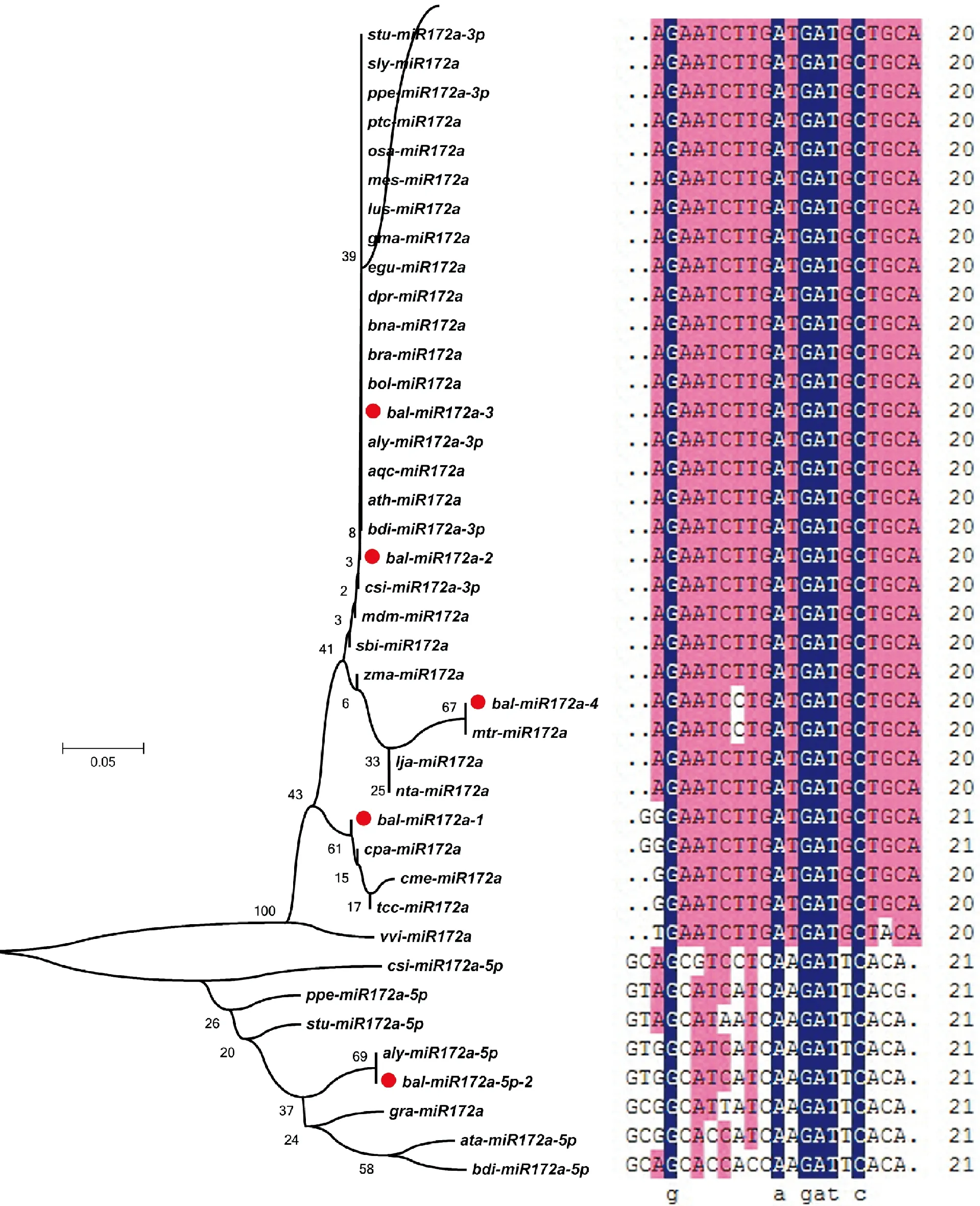
图4 miR172a家族成熟体序列分析Fig.4 Analysis of mature miR172a sequences
2.6 pre-miR172家族不同成员在芥蓝不同组织部位中的表达分析
通过实时荧光定量PCR,对芥蓝5个pre-miR172a成员和5个pre-miR172b成员在9个组织部位中的转录水平分别检测(图5、6),结果发现,10个成员在9个组织部位中的表达各不相同。pre-miR172a的5个成员中pre-miR172a-2和pre-miR172a-3表达比较高,pre-miR172a-4和pre-miR172a-5p-2表达比较低,pre-miR172a-1表达居中。pre-miR172a-2和pre-miR172a-3在生殖生长阶段主要在荚和种子里表达;pre-miR172a-1在营养生长阶段主要在叶中表达,在生殖生长阶段则主要在荚和种子里表达; pre-miR172a-4在营养生长阶段主要在根中表达;而pre-miR172a-5p-2在9个部位均有表达。pre-miR172b 5个成员中pre-miR172b-5p和pre-miR172b-5p-4表达比较高,pre-miR172b、pre-miR172b-5p-2和pre-miR172b-5p-3表达次之。pre-miR172b-5p、pre-miR172b-5p-2、pre-miR172b-5p-3和pre-miR172b-5p-4在营养生长阶段主要在叶中表达,在生殖生长阶段主要在花、荚和种子里表达,并且表达量都是花中最高;pre-miR172b在生殖生长阶段则主要在荚、花和种子中表达。由此看出芥蓝从营养生长阶段到生殖生长阶段miR172b的表达有从叶片到花的一个很明显的转移。


A~C.营养生长阶段的根、茎、叶;D~I. 生殖生长阶段的根、茎、叶、花、荚和种子图5 芥蓝营养生长期和生殖生长期的器官A-C. Root, stem and leaf in the stage of vegetative growth; D-I. Root, stem, leaf, flower, pod and seed in the stage of reproductive growthFig.5 Different tissues of B. alboglabra
3 讨 论
3.1 植物miR172家族进化特性
miR172是一类保守的miRNA,在植物中广泛存在。虽然miR172家族成员的长度和数目在不同的植物中各不相同,但在进化上保守性极强,近缘物种中序列几乎一致,说明miR172的功能和作用方式在不同物种中可能也具有潜在的保守性。芥蓝中,miR172有6个成员,分别是bal-miR172a、bal-miR172b、bal-miR172c、bal-miR172d、bal-miR172e和bal-miR172j。其中miR172a又有5个成员:bal-miR172a-1、bal-miR172a-2、bal-miR172a-3、bal-miR172a-4和bal-miR172a-5p-2;miR172b也有5个成员:bal-miR172b、bal-miR172b-5p 、bal-miR172b-5p-2、bal-miR172b-5p-3和bal-miR172b-5p-4。
miR172不同成员的启动子结构和顺式调控元件组成并不相同,故而其表达具有时空特异性,说明这些成员在植物发育的不同阶段发挥着各不相同或者协同调控的作用[10]。对于miR172不同成员的研究,在拟南芥、水稻、大豆和月季中均有报道。拟南芥中miR172共有5个成员,包括2个miR172a、2个miR172b和1个miR172c。拟南芥生长过程中miR172a-1、miR172a-2和 miR172b-1的表达量逐渐增加,miR172b-2 和miR172c的表达水平则非常低,未表现出与植物营养生长和生殖生长发育转换的相关性[11]。在水稻中,miR172共有4个成员,miR172a、miR172b、miR172c和miR172d。水稻中4个miR172成员的表达量在幼苗期均上升,在谷粒中miR172a、miR172b和miR172d均有表达,但miR172c则未检测到[12]。在大豆中,共发现12个miR172成员,其中miR172a、miR172b、miR172c、miR172d和miR172e前体的表达水平较高,且主要在大豆的真叶中表达[13]。月季中共有12个miR172成员,其中miR172a在花柱中大量表达,且随着日照变化呈现规律性的波动,而miR172c表达很少[14]。
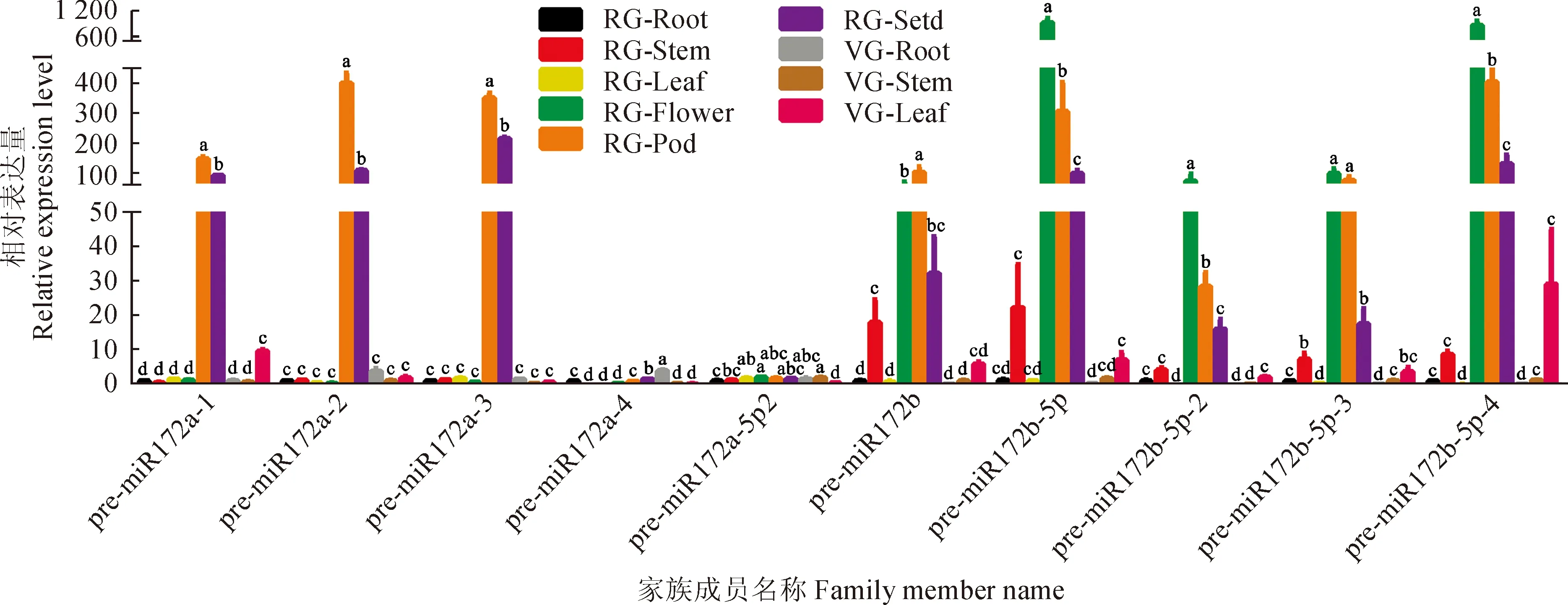
图中柱形图上不同小写字母表示组织间差异显著(P<0.05)。RG-Root.生殖生长阶段的根;RG-Stem.生殖生长阶段的茎;RG-Leaf.生殖生长阶段的叶;RG-Flower.生殖生长阶段的花;RG-Pod.生殖生长阶段的3.5 cm长的除去种子的荚;RG-Seed.生殖生长阶段的3.5 cm长的荚对应的种子;VG-Root.营养生长阶段的根;VG-Stem.营养生长阶段的茎;VG-Leaf.营养生长阶段的叶图6 pre-miR172a和pre-miR172b家族在芥蓝不同组织部位中的表达分析The different lowercase letters on the graph indicate significant difference among tissues and stages at 0.05 level(P<0.05).RG-Root. Root of the reproductive growth; RG-Stem. Stem of the reproductive growth; RG-Leaf. Leaf of the reproductive growth; RG-Flower. Flower of the reproductive growth; RG-Pod. Pod of the reproductive growth; RG-Seed. Seed of the reproductive growth; VG-Root. Root of the vegetative growth; VG-Stem. Stem of the vegetative growth; VG-Leaf. Leaf of the vegetative growthFig.6 Expression pattern of pre-miR172a and pre-miR172b family in different developmental stages and tissues of B. alboglabra
3.2 芥蓝中miR172的表达模式及其潜在功能
芥蓝中miR172a和miR172b的表达模式不同,miR172a主要在生殖生长期的荚和种子中表达,miR172b则主要在花中表达,说明miR172a和miR172b在功能上有分工。miR172可以与植物花型同源基因AP2的信使RNA进行碱基配对,在翻译水平上调控其表达。miR172通过调控靶基因AP2促进植物由营养生长向生殖生长转换的功能在很多物种中已经发现,研究认为这种促进作用在高等植物中广泛存在。大豆中gma-miR172a有一种昼夜节律表达的模式,随着植物的生长,直到大豆开花,它的丰度急速增加。gma-miR172a的过表达植株中成花素的含量增加,出现了早花表型,抗gma-miR172a的Glyma03g33470基因的过度表达削弱了早期开花显型的表现[15]。在水稻中,miR172通过调控属于AP2家族的OsTOEl和SNB(SUPERNUMERARYBRACT)进而参与水稻的生殖生长过程[10]。芥蓝的pre-miR172b-5p和pre-miR172b-5p-2在花中的大量表达说明其在芥蓝花的发育中起着重要作用。
除了对开花的促进作用,近年来miR172调控果实发育的功能在高等植物中也有报道,miR172对果实发育的调控依赖于物种果实的类型。在拟南芥中,miR172的过表达可以促进拟南芥花荚的生长,拟南芥的荚果来源于心皮组织,受到AP2的负调控,增加miR172的表达水平会减少靶基因AP2的含量,进而减轻对种荚生长的抑制作用[4]。在苹果中,过表达miR172却使得苹果的果实变小。苹果的果实是梨果,由萼片、花瓣和雄蕊等组成的萼筒发育而来,其中萼片主要发育为果实。由于AP2正调控萼片的发育,过表达苹果的miR172会减少其AP2的积累,影响萼片发育,最终导致果实变小[5]。番茄的浆果是由子房发育而来的,过表达miR172则会使得番茄出现只剩心皮的花,最终发育成为单性果实[11]。
