不同小麦品种叶锈菌的遗传多样性分析
2018-09-28张梦雅孟庆芳闫红飞刘大群
张梦雅,孟庆芳,张 林,高 颖,闫红飞*,刘大群,2*
(1.河北农业大学 植物保护学院/国家北方山区农业工程技术研究中心/河北省农作物病虫害生物防治工程技术研究中心,河北 保定 071000; 2.中国农业科学院 研究生院, 北京 100081)
由小麦叶锈菌(Pucciniatriticina)引起的小麦叶锈病是小麦生产上的重要病害之一,在世界各麦区均有发生且危害严重[1]。由于各麦区栽培品种、发病早晚不同,小麦叶锈病造成的产量损失在7%~30%,严重时达50%以上[2]。20世纪70年代我国曾多次流行小麦叶锈病,造成了巨大的经济损失[3]。近年来,随着小麦叶锈菌群体组成、环境气候和小麦品种的变化,该病害的发生日趋严重。2012年全国范围小麦叶锈病发生严重,2013年山东、河南和新疆局部地区发生严重,2015年黄淮海地区发生大流行,尤其以河南、安徽等省发生严重。小麦叶锈菌新生理小种的出现、群体组成和结构的变化对小麦叶锈病的发生流行起重要作用[4],研究小麦叶锈菌小种和寄主品种的关联性对掌握小麦叶锈菌的群体遗传结构和流行动态、病害防控及小麦抗病育种具有重要意义。
目前,小麦叶锈菌遗传多样性研究主要基于毒性和分子生物学2个方面。毒性分析法需要进行小种的分离纯化、扩繁和接菌鉴定等,虽然直观,但步骤繁琐、工作量大、耗时长,且容易受鉴别寄主、环境条件和人为因素的影响[5]。分子生物学手段快速、简便和精准,且不易受发育时期限制和环境因素影响,已成为分析小麦叶锈菌遗传多样性的一种有效方法[6]。简单重复序列标记(SSR)[7]、随机扩增多态性片段技术(RAPD)[8]、通用引物PCR技术(UP-PCR)[9]等均有应用报道。赵盼盼等[10]曾利用21对EST-SSR(基于表达序列标签开发微卫星的一种新型分子标记)引物对不同地区、不同年份的小麦叶锈菌菌株进行了分子多态性分析,发现叶锈菌群体间的亲缘关系与地理分布具有一定的相关性,用分子手段证明小麦叶锈菌遗传多样性较高,但不同群体间多样性水平有差异。这些分子生物学技术的广泛应用为小麦叶锈菌遗传多样性的研究提供了快速、简便的方法。
2015年小麦叶锈病在黄淮海地区大流行[11],河南省大部分地区发病更是严重。为了揭示不同小麦品种普遍发病的原因,以河南周口小麦试验田的品种为研究对象,采用分子快速检测方法对区试田小麦叶锈菌的群体组成及遗传结构进行分析,为小麦叶锈病防控、小麦品种合理布局提供指导。
1 材料和方法
1.1 小麦叶锈菌样品采集
供试小麦叶锈菌样品采集于周口市南、西的2块周口市农业科学院小麦品种区试苗圃,2015年随机采取不同品种叶片上的叶锈菌,以样本症状典型、发病充分、菌量大为标准,直接将小麦叶片上叶锈菌夏孢子堆收集于1.5 mL离心管中,每管约50 mg,干燥后储存于4 ℃冰箱中备用。共采集63份样品,详见表1。

表1 小麦叶锈菌菌株编号及来源
续表1 小麦叶锈菌菌株编号及来源
注:菌株1—27来源于周口南地块,28—63来源于周口西地块。
1.2 EST-SSR扩增及电泳检测
参照康健等[12]的Chelex-100法对63份供试菌株提取DNA。试验采用Wang等[13]报道的17对EST-SSR引物。PCR反应体系10 μL:模板DNA(30 ng/μL) 1.0 μL,TaqDNA聚合酶(2.5 U/μL)0.1 μL,10×PCR Buffer 1.0 μL,dNTPs(10 mmol/L) 0.2 μL,上、下游引物(5 mmol/L)各0.2 μL,无菌超纯水7.3 μL。反应程序:94 ℃预变性5 min;94 ℃变性30 s,54~61 ℃退火30 s,72 ℃延伸1 min,35个循环;72 ℃延伸5 min;4 ℃保存。PCR产物经10%非变性聚丙烯酰胺凝胶电泳检测。
1.3 数据分析
统计PCR扩增产物清晰可重复的条带,构建0、1矩阵(有带为1,无带为0)。利用NTSYS 2.1软件对数据进行UPGMA聚类分析,利用POPGENE软件进行群体遗传多样性分析,利用Arlequin 3.11软件对小麦叶锈菌的分子变异进行分析。
2 结果与分析
2.1 小麦叶锈菌EST-SSR分析
检测结果表明,17对EST-SSR引物中仅Ptssr5649、Ptssr2948未扩增出多态性条带,表明该引物对叶锈菌小种不具有区分能力;而其余15对引物可在63个供试叶锈菌株基因组中扩增出稳定清晰的多态性条带,用以区分不同的叶锈菌小种。63份小麦叶锈菌材料扩增出62种带型,代表62种分子小种,14号小麦品种俊达109与35号小麦品种粮源A8上收集的叶锈菌材料扩增带型完全一致,表明其为同种小麦叶锈菌的分子小种。
2.2 小麦叶锈菌聚类分析
利用NTSYS 2.1软件对63个供试小麦叶锈菌株的EST-SSR引物扩增结果进行聚类分析,其相似系数为0.70~1.00,表明在小麦叶锈菌群体内存在一定的遗传多样性,结果见图1。在相似系数0.716处将63个菌株聚为V1、V2和V33组。V1组共包括6个菌株,其寄主品种为郑麦369、豫农306、安农1206、泉麦890、涡麦1212、秋乐2128,除豫农306和泉麦890外,其他4个品种亲本来源相同,均为辉县红。V3组包括7个菌株,其寄主品种为皖科981、安科1403、绿雨1408、瑞星麦618、郑品麦22、中育1152、新麦33,其中除皖科981和郑品麦22外,其他5个品种的亲本来源相同,均为豫麦2号。V2组的50个菌株在相似系数0.737处分为V2-1和V2-22个亚组。其中,V2-1共5个菌株,其寄主品种为偃高21、中金13、洛麦26、豫粮1688、商麦167,亲本来源均为豫麦2号。V2-2的45个菌株在相似系数0.775处分为V2-2-1和V2-2-22个组,其中品种阜麦18上的叶锈菌株独立形成V2-2-2组,与V2-2-1组的菌株遗传相似性较低,亲缘关系较远。V2-2-1组包含44个菌株,寄主品种徐麦0054、乐麦Z1311273和龙科1109上的叶锈菌株聚在同一分支,其亲本来源相同,均来自豫麦2号;俊达109、粮源A8、郑麦113、金麦201、机麦211、耕丰麦1号、矮丰339上的7个菌株聚在同一分支,其寄主亲本也均来自豫麦2号。俊达109和郑育麦16两个品种系谱来源完全相同,均来自豫麦2号/郑91138//济麦4号,但2个菌株间的遗传相似性低于俊达109和粮源A8之间的遗传相似性。以上分析表明,叶锈菌遗传多样性与寄主品种的系谱来源有很大关系,一般情况下寄主品种的系谱来源越相似,其上的叶锈菌遗传相似性越高,亲缘关系越近。
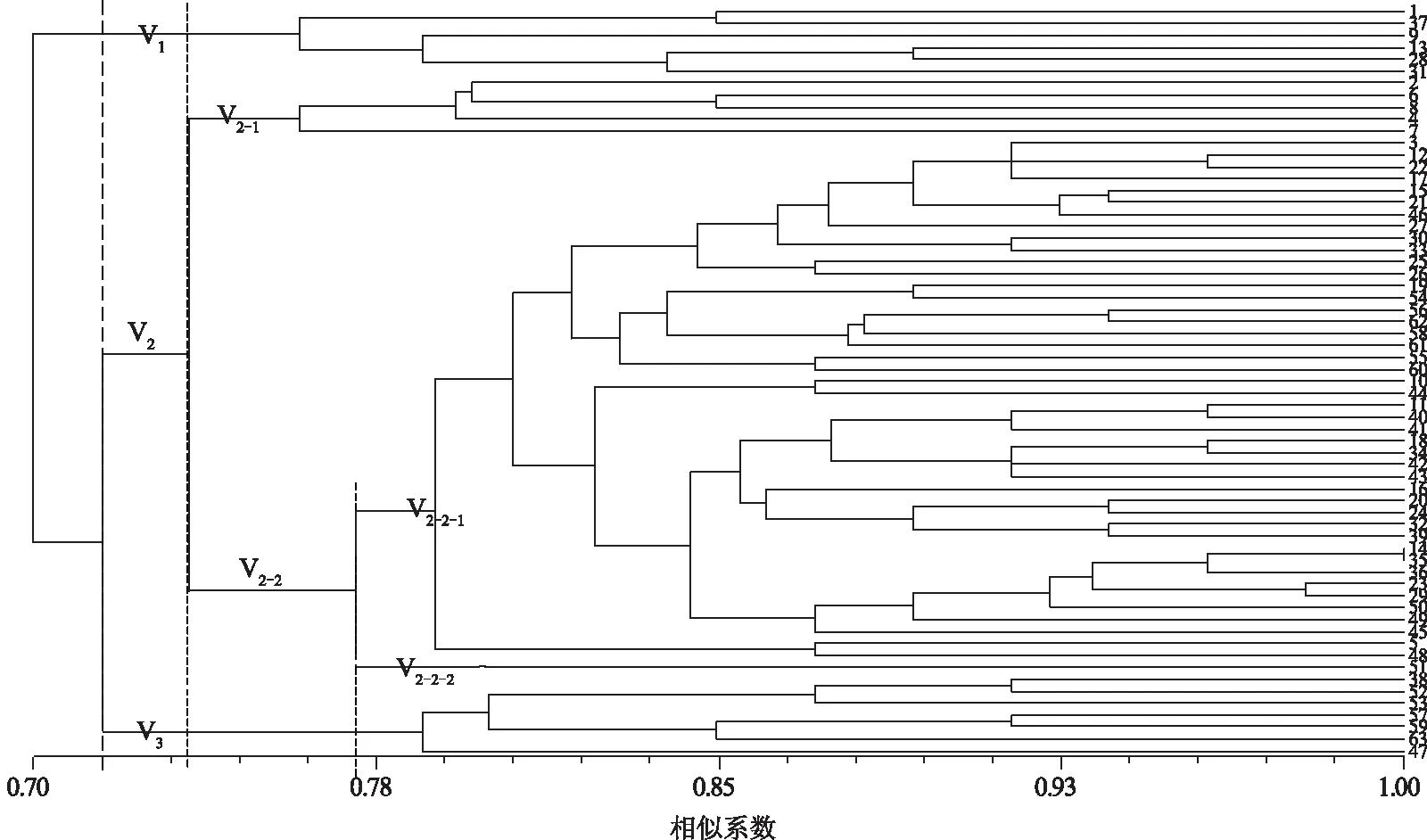
1—63代表小麦叶锈菌株
2.3 小麦叶锈菌小种和寄主品种的相关性分析
在遗传相似系数0.96处,11号菌株(寄主品种徐麦0054)比40号菌株(寄主品种乐麦Z1311273)多一条238 bp条带,少一条242 bp条带,其寄主亲本中均含有豫麦2号。在遗传相似系数0.98处,23号菌株(寄主品种郑麦113)比29号菌株(寄主品种金麦201)仅多一条238 bp的条带,其寄主亲本的来源为郑91138和豫麦2号。在相似系数1.00处,14号菌株(寄主品种俊达109)与35号菌株(寄主品种粮源A8)带型完全一致,其寄主亲本的来源均为豫麦2号。由此可见,品系来源越相似,该品种上的小麦叶锈菌分子小种越相似,遗传多样性越小。据此推测每个品种上的小麦叶锈菌可能由多种不同的叶锈菌株混合而成,聚在一个组里表明这些品种上的小麦叶锈菌菌株相同或相似。
2.4 小麦叶锈菌群体遗传多样性分析
利用POPGENE软件对河南周口2块苗圃叶锈菌株进行群体遗传多样性参数分析。在物种水平和群体水平上,Nei’s基因多样性指数(H)均为0.22,多态位点百分率(P)分别为78.72%和73.41%,表明2块苗圃供试小麦叶锈菌遗传多样性丰富。周口南和周口西地块叶锈菌群体的Nei’s基因多样性指数分别为0.21和022,Shannon信息指数(I)分别为0.32和0.34,多态位点百分率分别为72.34%和74.47%,表明周口西地块叶锈菌群体比周口南的遗传多样性稍高(表2)。
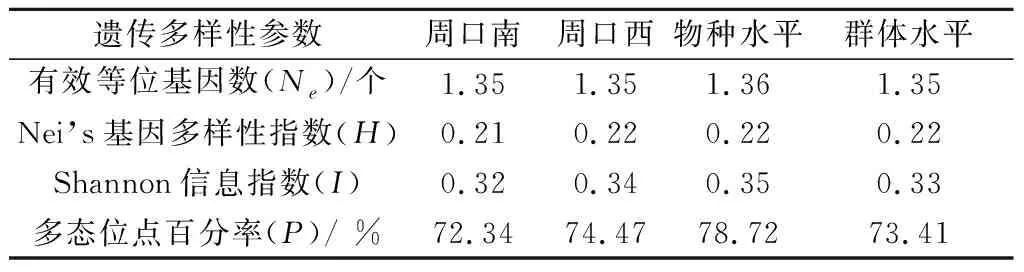
表2 小麦叶锈菌基于EST-SSR的群体遗传多样性参数
2.5 小麦叶锈菌群体的遗传变异分析
利用POPGENE软件对周口2个地块的所有供试叶锈菌群体进行遗传多样性分析,结果显示,供试叶锈菌株总遗传多样性(Ht)为0.22,群体内遗传多样性(Hs)为0.21,群体间遗传多样性(Dst)为0.01,遗传分化系数(Gst)为0.05,群体内遗传多样性占总遗传多样性的95.45%,而群体间遗传多样性仅占4.55%,说明叶锈菌群体内遗传多样性显著高于群体间。Nei’s遗传相似系数为0.978 7,遗传距离为0.021 5,表明2个地块间不同寄主小麦品种的叶锈菌群体分子结构类似。利用Arlequin 3.11软件的AMOVA程序对2个群体的分子变异分析表明,2个地块的小麦叶锈菌遗传变异发生在群体间和群体内,其中群体内遗传变异占总遗传变异的94.44%,而群体间遗传变异仅占总变异的5.56%,这表明小麦叶锈菌的遗传变异主要发生在群体内部(表3)。
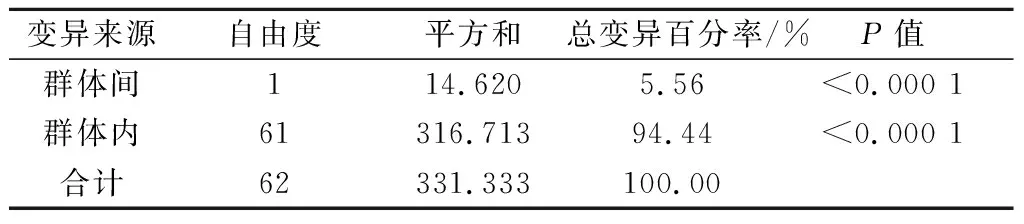
表3 小麦叶锈菌群体间和群体内分子变异的AMOVA分析结果
3 结论与讨论
小麦叶锈菌遗传多样性研究有毒性鉴定和分子标记2种方法,前者主要依据病原菌对小麦苗期的致病能力划分生理小种,操作繁琐,费工费时,准确性易受环境条件影响;后者是从基因产生的变异反映病原菌个体的差异,从本质揭示群体遗传结构等特性,操作简便且准确性高。2007年,Ordoez等[14]对北美硬粒小麦和普通小麦上叶锈菌群体的SSR分子遗传多态性进行了分析,结果发现,北美硬粒小麦品系来源相似且单一,其叶锈菌遗传多样性远远低于亲本来源不同的普通小麦遗传多样性,因此,不同亲本来源小麦品种可对小麦叶锈菌的遗传多样性造成一定的影响。刘二明[15]在研究稻瘟病菌遗传多样性时发现,在某一水稻种植区内,水稻栽培品种越多,其上分离出的稻瘟病菌株越多,分属的谱系和单元型越多,则该稻区的稻瘟病菌群体遗传多样性也更丰富,即稻瘟病菌群体的遗传多样性与水稻栽培品种的多样性呈较明显的正相关。在本研究中,聚类分析图显示小麦叶锈菌遗传多样性与寄主品种有一定关系,例如V2-1组包括偃高21、中金13、洛麦26、豫粮1688、商麦167上的菌株,而这5个菌株寄主的亲本来源均包含豫麦2号,表明品种系谱来源越相似,其上叶锈菌的遗传相似性越高,亲缘关系越近,遗传多样性越低。在遗传相似系数0.844处,可将63个小麦叶锈菌株分为20个组(S1—S20),其中周口南地块的27个菌株分属于13个组,而周口西地块的36个菌株分属于15个组。利用POPGENE软件对两苗圃叶锈菌进行群体遗传多样性分析表明,周口西地块小麦叶锈菌群体遗传多样性水平稍高于周口南,由此可见,小麦叶锈菌遗传多样性与地理来源并无明显关系。
本研究通过EST-SSR分析表明,群体内变异为小麦叶锈菌的主要变异来源,与群体间遗传多样性相比,群体内遗传多样性明显较高,这与Dadrezaie等[16]、Wang等[17]、许敏青等[6]、赵盼盼等[10]的研究结果一致,同样证明叶锈菌存在丰富的遗传多样性。小麦寄主品种亲本来源越相似,其上的叶锈菌小种类型越一致,亲缘关系越近,遗传多样性越低。本研究揭示了寄主小麦品种及其亲本来源与小麦叶锈菌遗传多样性的关系,为小麦的育种及抗病研究奠定理论基础,对小麦生产实践具有现实的指导意义。
