eEF2单克隆抗体制备及在免疫组化中的应用
2018-09-27何成彦令狐志宏宋丽娜
王 海,何成彦,令狐志宏,宋丽娜
(吉林大学中日联谊医院 检验科,吉林 长春130033)
真核延伸因子2 (eEF2)是一种GTP依赖的转位酶,可介导核糖体的移位,因此与蛋白质的延伸和翻译密切相关[1]。eEF2主要受eEF2K (真核延伸因子2激酶)的调控,是蛋白质合成过程中的关键调控因子,在肿瘤的发生、发展中起到重要的作用[2-5]。本研究以纯化的eEF2融合蛋白为抗原免疫Bal b/c小鼠,制备eEF2单克隆抗体,探究大肠癌组织中eEF2的表达情况,为进一步研究其对大肠癌生物学行为的影响奠定基础。
1 材料和方法
1.1材料
主要试剂有不完全弗氏佐剂、完全弗氏佐剂购自 Sigma公司,胎牛血清、RPMI 1640 购自Gibco公司,100×HT、100×HAT、PEG 4000购自Sigma 公司,辣根过氧化物酶标记链霉素抗生物素蛋白工作液(S-A/HRP)、山羊抗鼠Ig G(H+L)-HRP购自CW bio。
1.2实验方法
1.2.1小鼠的免疫 首次免疫选取6周龄的雌性Bal b/c小鼠,将纯化的eEF2融合蛋白与完全弗氏佐剂按1∶1的比例混合乳化后在小鼠背部皮下进行多点免疫。2周后进行第二次免疫(不完全弗氏佐剂)。再两周后进行第三次加强免疫(不完全弗氏佐剂)。3天后从小鼠的尾静脉采血,用ELISA法检测抗体效价。
1.2.2间接ELISA法检测抗体效价 取浓度为2 μg/ml的eEF2抗原包被聚苯乙烯微孔板,每孔100 μl,4℃过夜。洗板后每孔加封闭液150 μl,37℃放置2小时,洗板后加入1∶1000、1∶3000、1∶9000、1∶27000等倍比稀释的待测血清、空白对照和阴性对照,每孔100 μl, 37℃孵育30 min,洗板。每孔加入1∶5 000倍稀释的HRP标记的羊抗鼠IgG 100 μl,37℃孵育30 min,洗板。每孔加入1×TMB 100 μl,37℃避光显色20 min。加入终止液。用酶标仪测定OD450nm,测定孔与阴性对照孔的比值(P/N)>2.1为确定效价的临界点。
1.2.3细胞融合及阳性克隆筛选 将骨髓瘤细胞Sp2/0与小鼠脾细胞以1∶10的比例混合,用50%PEG为融合剂,将融合细胞加入96孔板中,置于37℃、5%CO2的培养箱内培养。于细胞融合后第8-14天,取细胞上清,用间接ELISA方法进行筛选。
1.2.4单克隆抗体大量制备 取12周龄Bal b/c小鼠,腹腔内注射0.5 ml灭菌的石蜡油,1周后接种 2×106个细胞/ml生理盐水,观察腹水的产生情况。1-2周后收集腹水,离心收集上清,间接ELISA法检测效价。
1.2.5采用亲和层析法纯化腹水抗体 用protein G亲和层析法纯化抗体。
1.2.6Western Blot 鉴定制备出的eEF2单克隆抗体的特异性 以纯化的eEF2融合蛋白上样,进行Western blot测定,以制备的eEF2单克隆抗体为一抗,HRP-羊抗鼠IgG为二抗,用ECL溶液进行显影分析。
1.2.7应用制备的单克隆抗体对大肠癌组织进行免疫组化染色 大肠癌组织切片用二甲苯脱蜡;水化;柠檬酸钠抗原修复;3%H2O2室温孵育;5%兔血清封闭;滴加一抗(制备的eEF2单克隆抗体)、二抗(HRP-羊抗鼠IgG)、S-A/HRP;DAB显色;苏木素复染;0.1%盐酸分化;梯度酒精逐级脱水;二甲苯透明;中性树脂封片。显微镜下观察、拍照。
2 结果
2.1WesternBlot结果
将纯化的eEF2融合蛋白进行Western Blot 鉴定,一抗用本研究制备的eEF2单克隆抗体,二抗用山羊抗鼠Ig G。如图1所示,该单抗可与eEF2融合蛋白特异性结合,可确定其为eEF2的特异性抗体。
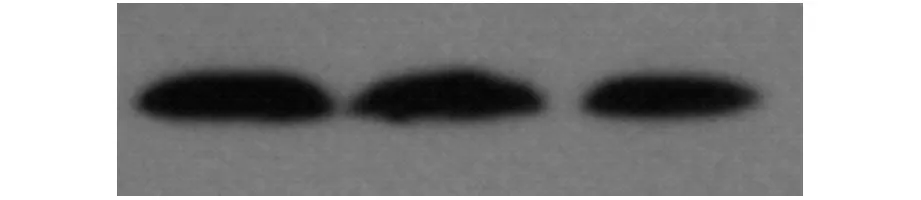
图1 Western Blot结果
2.2大肠癌组织的免疫组织化学检测结果
应用制备的单克隆抗体对大肠癌组织切片进行免疫组化染色,阳性结果的判定标准为:细胞内出现清晰的棕褐色或棕黄色颗粒。如图2所示,大肠癌组织中的eEF2主要表达在细胞浆。
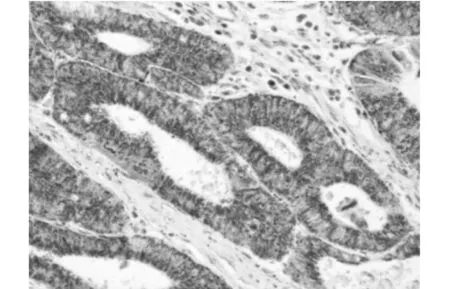
图2 eEF2在大肠癌组织中的表达(╳200)
3 讨论
自从开发抗体生产技术以来,已经有许多抗体被研发并应用于免疫学、生物技术、诊断学和治疗药物以及科研领域。多克隆抗体最常用,但不同批次之间具有性能差异,每个新批次需要进行验证,同时,多克隆抗体含有大量不同浓度的具有未知特异性的不同抗体。相反,单克隆抗体性能一致,是具有高度特异性的一类生物试剂[6],因此现在的很多研究都用单克隆抗体代替多克隆抗体,以提高实验的稳定性和特异性。应用于抗体研发的方法有很多,应用杂交瘤技术开发的第一种单克隆抗体于1975年报道,随后于1986年获得许可[7],本研究应用该方法制备了具有高特异性的eEF2单克隆抗体,为将来对eEF2的研究工作奠定了基础。研究显示,eEF2蛋白在食管癌、肺癌、乳腺癌、前列腺癌、胰腺癌等多种肿瘤组织中过表达[8-12],并且它可以促进细胞周期的G2/M的进展,并伴随着Akt和G2 / M调节剂cdc2蛋白的激活,导致体外增强和体内癌细胞生长[13]。然而,eEF2在肿瘤发生中的作用仍然大部分未知,此外,eEF2基因产物是免疫原性的,eEF2是否可以成为癌症的免疫疗法的靶分子仍有待进一步深入研究。
