甘蔗ScWRKY4基因的克隆与表达特性分析
2018-09-11戴明剑孙婷婷苏炜华王春风旭毛花英苏亚春阙友雄
王 玲 刘 峰 戴明剑 孙婷婷 苏炜华 王春风 张 旭毛花英 苏亚春,2, 阙友雄,2,
甘蔗基因的克隆与表达特性分析
王 玲1刘 峰1戴明剑1孙婷婷1苏炜华1王春风1张 旭1毛花英1苏亚春1,2,*阙友雄1,2,*
1福建农林大学 / 农业部福建甘蔗生物学与遗传育种重点实验室, 福建福州 350002;2福建农林大学 / 教育部作物遗传育种与综合利用重点实验室, 福建福州 350002
WRKY是植物中特有的转录因子家族之一, 在植物对生物和非生物逆境胁迫的应答反应中起重要调控作用。本研究基于课题组前期构建的甘蔗(spp.)转录组数据库, 从新台糖22 (ROC22)中成功克隆到1个基因, 命名为(GenBank登录号为MG852087)。序列分析发现,基因cDNA全长1265 bp, 包含1个741 bp的完整开放读码框, 编码246个氨基酸, 该蛋白具有1个WRKYGQK保守结构域和C2H2锌指结构域, 属于IIc类WRKY转录因子。生物信息学预测分析发现, ScWRKY4蛋白为碱性的不稳定亲水性蛋白, 不存在信号肽和跨膜结构域, 蛋白二级结构元件缺少β螺旋。在本氏烟()叶片瞬时表达中, ScWRKY4蛋白定位于细胞核。酵母杂交实验结果显示, ScWRKY4不具有转录激活活性。实时荧光定量PCR分析表明,基因在甘蔗的根、叶、芽和皮中的表达量无明显差异, 在蔗肉中的表达量最高, 为对照蔗根的18.38倍; 黑穗病菌侵染0~72 h,在抗病品种崖城05-179中下调表达, 在感病品种ROC22中表达较稳定; 受到外源激素脱落酸、水杨酸和茉莉酸甲酯以及非生物胁迫因子氯化钠和聚乙二醇胁迫后,基因均被诱导上调表达。上述研究结果表明,基因可能不参与甘蔗对黑穗病的抗性反应或在该防御方面起负调控作用, 但积极响应甘蔗对盐和干旱胁迫的应答。
甘蔗; WRKY转录因子; 生物信息学; 亚细胞定位; 转录激活活性; 实时荧光定量PCR
植物在生长过程中, 依赖其体内各种功能基因的表达来适应环境, 特别是在生物和非生物逆境胁迫条件下, 这些相关功能基因在转录水平上的表达和修饰就显得尤其重要。转录因子又称反式作用因子, 可以结合在靶基因上游的顺式作用元件上, 对相关功能靶基因的转录效率进行正向或负向调控[1]。目前, 对植物逆境相关转录因子的研究较多的是WRKY、MYB (myeloblastosis)、NAC [NAM, ATAF1(2), CUC2]、bZIP (basic letcine-zipper)和AP2/ERF (APETALA 2/ethylene-responsive element binding factor)五大类[2]。自1994年首个甘薯WRKY转录因子家族基因(SWEET POTATO FACTORS)[3]被克隆以来, 逐渐有大量的基因从多种植物中被分离鉴定。基因家族成员在模式植物拟南芥()中有72个[4]、水稻()中有105个[5]、高粱()中有68个[6]、大麦()中有45个[7]、玉米()中有119个[8]。
1996年Rushton等[9]发现欧芹()的WRKY转录因子WRKY1、WRKY2和WRKY3在基因介导的免疫应答中具有调控作用。拟南芥72个基因中有49个受到丁香假单胞菌()和水杨酸(salicylic acid, SA)的诱导表达[10]。利用全基因组分析大麦转录因子的表达图谱发现,和负调控大麦对白粉菌()的抗性[11]。水稻有15个WRKY转录因子可以被稻瘟病菌()诱导表达, 其中12个能够同时被白叶枯病菌(pv.)诱导表达[12]; 在水稻中过表达基因能够显著增强其对白叶枯病病菌和稻瘟病菌的抗性[13];在水稻的先天免疫过程中起负调节作用[14]。拟南芥的WRKY转录因子中有30个响应盐胁迫, 其中23个上调表达, 7个下调表达[15]。菜豆()的88个WRKY转录因子中, 有19个受干旱胁迫诱导表达, 其中11个下调, 8个上调[16]。转录组分析发现, 杨树()的100个WRKY转录因子中, 有61个受到杨树黑斑病菌()、SA、茉莉酸甲酯(methyl jasmonate, MeJA)、损伤、冷胁迫、盐胁迫等诱导表达[17]。此外, 玉米中有58个基因受干旱胁迫诱导表达[18]。上述报道为研究植物转录因子在应答不同生物和非生物逆境胁迫下的耐受机制提供了基础。
甘蔗(spp.)不仅是最重要的糖料作物, 也是最有潜力的生物能源作物, 具有重要的生物学和经济价值。研究甘蔗在生长发育及其适应外界环境变化过程中的信号转导, 特别是转录因子的功能, 对甘蔗的高产、高效分子育种理论具有重要的参考意义。目前, 在甘蔗上仅有1篇关于基因的研究报道[19]。Liu等[19]发现基因(GenBank登录号为GQ246458)在甘蔗品种福农22 (FN22)中的表达受甘蔗黑穗病菌()、SA、氯化钠(sodium chloride, NaCl)和聚乙二醇(polyethylene glycol, PEG)的诱导, 表明该基因可能在甘蔗对黑穗病菌、干旱和高盐胁迫的响应机制中发挥作用。本研究基于课题组前期构建的甘蔗转录组数据库, 从甘蔗品种新台糖22 (ROC22)中成功分离获得1个新的甘蔗转录因子基因, 借助生物信息学软件预测了该基因的理化性质, 并验证了其亚细胞定位情况及转录激活活性, 同时利用实时荧光定量PCR (qRT-PCR)技术分析了基因在黑穗病菌及不同植物激素和非生物胁迫下的表达水平变化, 目的是为甘蔗抗逆分子育种提供基因资源。
1 材料与方法
1.1 材料处理
试验所用的甘蔗材料由福建农林大学农业部福建甘蔗生物学与遗传育种重点实验室提供, 分别为感黑穗病品种ROC22和抗黑穗病品种崖城05-179。参照黄宁等[20]的方法, 从甘蔗田随机带土挖取9株健康并且长势一致的植株, 立即洗净根部泥土, 从乳白色幼嫩的蔗根取样; 从地面以上完整可见的第一节间算起, 选取第7~8节间, 用75%酒精擦净其表皮, 从侧芽、蔗皮和蔗肉取样; 选取+1叶的蔗叶, 用酒精擦拭干净。选择3株作为一个重复, 共3个生物学重复。用锡箔纸包好所有材料, 立即投入液氮冷冻, –80℃保存备用。
为了分析目的基因在不同外源胁迫下的表达特性, 将ROC22健康植株的蔗茎砍成单芽茎段, 清洗干净并温水脱毒处理后, 在塑料托盆中沙培育苗3个月左右, 至其长出第4或第5叶, 挑选健壮且长势一致的蔗苗, 将根清洗干净后放入盛有清水的塑料盆中炼苗培养10 d, 开始试验处理。第一组分别在250 mmol L–1NaCl和25.0% PEG-8000水溶液中培养, 于处理后0、0.5、3、6和24 h进行叶片取样; 第二组在叶面分别喷施100 μmol L–1脱落酸(abscisic acid, ABA)、5 mmol L–1SA (含0.01%吐温-20,)和25 μmol L–1MeJA, ABA处理在0、0.5、3、6和24 h进行叶片取样, SA和MeJA处理在0、3、12和24 h进行叶片取样。黑穗病菌处理材料为32℃培养箱催芽至2 cm左右的蔗芽, 针刺接种5×106个 mL–1(含0.01%吐温-20, v/v)浓度的黑穗病菌孢子悬浮液, 同时, 对照以无菌蒸馏水(含0.01%吐温-20, v/v)接种, 于28℃下16 h光照/8 h黑暗培养, 分别于处理后0、24、48和72 h进行蔗芽取样。每3株作为1次生物学重复, 每组设置3个生物学重复。用锡箔纸包好所有材料, 立即投入液氮冷冻, –80℃保存备用。
1.2 甘蔗ScWRKY4基因的克隆与生物信息学分析
利用TRIzol Reagent (Invitrogen, Carlsbad, CA, USA)提取ROC22叶片总RNA, 参照RevertAid First Strand cDNA Synthesis Kit (Fermentas, 中国上海)说明书将其逆转录为cDNA, 作为基因克隆的模板。从课题组前期构建的甘蔗受黑穗病菌侵染的转录组数据库[21]中, 挖掘到一条WRKY转录因子的Unigene序列(转录本ID: c128160.graph_c0), 命名为, 利用NCBI在线软件设计该基因全长扩增引物对-F/R (表1)。PCR反应总体系为25 μL, 包含cDNA模板1.0 μL、上下游引物(10 μmol L–1)各1.0 μL、ExDNA酶(5 U μL–1) 0.125 μL、10×Ex缓冲液(Mg2+plus) 2.5 μL、dNTPs混合物(2.5 mmol L–1) 2.0 μL和ddH2O 17.375 μL。PCR反应条件为94℃预变性4 min; 94℃变性30 s, 57℃退火30 s, 72℃延伸2 min, 35个循环; 最后72℃再延伸10 min。反应产物经1%琼脂糖凝胶电泳检测, 参照Gel Extraction Kit (天根, 中国北京)说明书纯化回收后, 再将其连接到pMD19-T (TaKaRa, 中国大连)克隆载体上并转化大肠杆菌DH5α感受态细胞(天根, 中国北京), 最后随机挑取单菌落进行PCR鉴定, 并将检测呈阳性的菌落送至尚亚有限公司测序分析。
通过NCBI的ORF finder (https://www.ncbi.nlm. nih.gov/orffinder/)在线程序和Conserved domains (http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)数据库, 分析测序获得的cDNA序列的开放阅读框(open reading frame, ORF)及保守功能结构域。运用ProtParam (https://web.expasy.org/protparam/)和GOR IV (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_ automat.pl?page=/NPSA/npsa_hnn.html/)程序, 预测ScWRKY4蛋白的一级和二级结构特征。利用在线程序SignalP 4.1 Server (http://www.cbs.dtu.dk/ services/SignalP/)、TMHMM Server v. 2.0 (http://www. cbs.dtu.dk/services/TMHMM/)和NetPhos 2.0 Server (http://www.cbs.dtu.dk/services/NetPhos/)分析基因编码氨基酸的信号肽、跨膜结构域和磷酸化位点。借助Psort (http://www.psort.org/)工具预测ScWRKY4蛋白的亚细胞定位情况。采用NCBI中Blastp程序, 查找与甘蔗ScWRKY4蛋白同源的氨基酸序列, 而后利用DNAMAN 6.0软件进行序列比对, 并通过ClustalW对ScWRKY4蛋白与甘蔗Sc-WRKY[19]、大麦[7]、拟南芥[15]、玉米[22]的基因编码的氨基酸序列进行多重比对分析, 使用MEGA 6.0软件中的最大似然法(maximum- likelihood, ML) (1000 BootStrap)构建系统进化树。
1.3 甘蔗ScWRKY4基因的亚细胞定位
根据基因克隆测序正确的序列的ORF (去除终止密码子)和亚细胞定位载体pCAMBIA 1300-(Clontech, 中国北京)图谱设计引物对-Subloc-F/R (表1), 以pMD19- T-质粒为模板, 进行PCR扩增, 将其胶回收产物和亚细胞定位载体质粒分别用相同的限制性内切酶H I和I (Fermentas, 中国上海)进行双酶切、胶回收及T4 DNA酶连接, 然后转化DH5α感受态细胞, 经菌液PCR检测、单酶切和双酶切验证及测序检测后, 得到pCAMBIA 1300-亚细胞定位融合表达载体。将pCAMBIA 1300-阳性质粒和pCAMBIA 1300-空载分别转化农杆菌菌株GV3101 (天根, 中国北京), 添加含有50 μg mL–1卡那霉素和35 μg mL–1利福平的LB液体培养基, 于28℃下250 转 min–1振荡培养过夜。设置离心机温度4℃、转速4000×、时间10 min, 收集菌体, 然后重悬于MS空白培养基并将浓度调至OD600=0.8, 加入200 μmol L–1乙酰丁香酮后黑暗静置30 min。选择5~8片叶龄且长势一致的本氏烟进行针刺注射, 在光照16 h/黑暗8 h条件下28℃培养2 d后, 将含有目的基因和空载的叶片剪下, 背光面朝下置1 μg mL–14’,6-二脒基-2-苯基吲哚(4’,6-diamidino-2-phenylindole, DAPI)染色剂中, 37℃暗培养30 min, 依次用NaCl水溶液脱色和清水洗净后, 背光面朝上置载玻片, 在Leica激光共聚焦显微镜(德国)下观察亚细胞定位情况[23]。

表1 引物序列及用途
下画线为添加的酶切位点H I (GGATCC)、I (TCTAGA)、I (CATATG)和H I (GGATCC); 双下画线为添加的保护碱基。
The added restriction enzyme sites ofH I (GGATCC),I (TCTAGA),I (CATATG), andH I (GGATCC) are underlined; the added protective bases in primers are double underlined.
1.4 甘蔗ScWRKY4的自激活活性分析
参照Matchmaker Gold酵母双杂交系统说明, 采用Y2HGold-GAL4酵母双杂交系统(含有4个报告基因:和)分析ScWRKY4的自激活活性。根据基因克隆测序正确的序列的ORF (去除终止密码子)和BD (DNA-binding domain)载体pGBKT7的图谱设计引物对-BD-F/R (表1), 以pMD19-T-质粒为模板, 进行PCR扩增, 将其胶回收产物和pGBKT7载体质粒分别用相同的限制性内切酶I和H I (Fermentas, 中国上海)进行双酶切、胶回收及T4 DNA酶连接, 转化DH5α感受态细胞, 经菌液PCR检测、单酶切和双酶切验证及测序比对后, 得到pGBKT7-重组载体。按照Y2HGold Chemically Competent Cell (TaKaRa, 中国大连)说明书操作, 将200 µg pGBKT7-重组质粒加入到100 µL酵母Y2HGold感受态细胞(全式金, 中国北京)中, 在SD/–Trp平板上29℃培养2~3 d, 至菌体长出。挑取3个单菌落同时于含有1 mL SD/–Trp液体培养基的2 mL无菌离心管中, 29℃、220 r min–1摇床震荡培养10 h左右, 稀释1×10–1、1×10–2和1×10–3三个浓度梯度, 取菌液8 µL分别于SD/–Trp、SD/–Trp (+X-α-Gal)和SD/–Trp (+X-α-Gal+AbA)平板上点样, 29℃培养箱中倒置培养, 对长出的菌落拍照记录。以杂交载体pGBKT7-53+pGADT7-T (已明确53蛋白和T蛋白在酵母细胞内能够发生结合, 该杂交载体在含有金担子素A (aureobasidin A, AbA)抗生素的平板上能激活报告基因)作为阳性对照, 空载pGBKT7 (含营养筛选标记)作为阴性对照, 分别转化Y2HGold, 并与pGBKT7-在相同平板上培养。通过观察酵母生长情况来判断ScWRKY4是否具有自激活活性。
1.5 甘蔗ScWRKY4基因表达模式分析
采用TRIzol法提取甘蔗不同组织(根、芽、叶、蔗肉和蔗皮)以及经ABA、SA、MeJA、PEG、NaCl和黑穗病菌胁迫处理样品的叶片总RNA, 按照PrimeScript RT Reagent Kit (Perfect Real Time) (TaKaRa, 中国大连)使用说明合成第1链cDNA, 以15倍稀释液作为qRT-PCR模板。在Primer Premier 5.0软件中设计基因的qRT-PCR检测引物-QF/R (表1), 以甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,) (GenBank Accession Number: CA254672)为内参基因[25], 引物-Q/F如表1所示。采用ABI 7500 Real-time PCR System (美国)进行qRT-PCR检测, 并按照SYBR Green PCR Master Mix Kit (Roche, 中国上海)说明书配置定量反应体系。qRT-PCR扩增程序为50℃ 2 min; 95℃预变性10 min; 95℃变性15 s, 60℃退火延伸1 min, 40个循环; 增加熔解曲线, 95℃ 15 s, 60℃ 1 min, 95℃ 15 s, 60℃ 15 s。每个样品设置3次技术重复, 阴性对照以无菌水作模板。采用2–ΔΔCt [26]计算基因相对表达量, 利用DPS 9.50软件分析试验数据的显著性水平, 并用Origin 8.0软件作柱状图。在黑穗病菌接种试验中, 由于目标基因的表达量会受到机械损伤的影响, 参照苏亚春等[27]的方法, 用接种黑穗病病原菌材料中基因的表达量减去对应时间接种无菌水材料中的表达量作为其相对表达水平。
2 结果与分析
2.1 甘蔗ScWRKY4基因的克隆
以甘蔗ROC22的叶片cDNA为模板、-F/R为PCR引物, 扩增到约1265 bp的单一条带(图1)。该扩增产物经纯化回收后与载体pMD19-T连接, 转化大肠杆菌DH5α感受态细胞, 经菌液PCR检测和测序鉴定, 得到图2序列。ORF finder分析显示,基因的ORF (137~877 bp)全长为741 bp, 编码246个氨基酸。
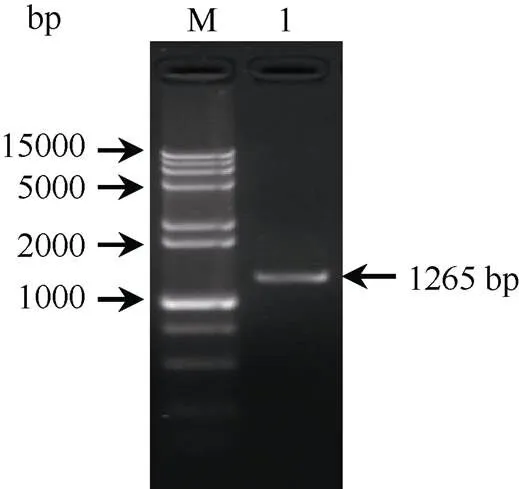
图1 甘蔗ScWRKY4基因的PCR扩增
M: 15000+2000 bp DNA marker; 1: PCR产物。
M: 15000+2000 bp DNA marker; 1: PCR product.

图2 PCR扩增获得的ScWRKY4基因的核酸序列及其推导的氨基酸序列(*: 终止密码子)
红色框部分为WRKYGQK基序; 黑色框部分为C2H2基序(CX4CX23HXH)。
The sequence of WRKYGQK motif is highlighted in red box, and that of C2H2motif (CX4CX23HXH) in black box.
2.2 甘蔗ScWRKY4基因的生物信息学分析
利用生物学软件对Sc基因序列编码的蛋白进行理化性质预测, 发现该蛋白的分子式为C1166H1794N332O365S15, 理论分子量为26 767.4 Da, 平均疏水性(GRAVY)为–0.49, 不稳定系数(II)为42.69, 负电荷残基(Asp+Glu) 22个, 正电荷残基(Arg+Lys) 28个, 等电点为8.71, 推测ScWRKY4蛋白为一个亲水性的不稳定碱性蛋白。二级结构预测显示甘蔗ScWRKY4蛋白主要由无规则卷曲(70.73%)、α-螺旋(17.48%)和延伸链(11.79%)结构组成, 而缺少β-螺旋结构。另外, 预测发现ScWRKY4蛋白不具有信号肽和跨膜结构域。磷酸化位点分析结果显示ScWRKY4蛋白含有12个丝氨酸(serine)、8个苏氨酸(threonine)和2个酪氨酸(tyrosine), 推测这些位点上可能发生磷酸化反应, 从而对ScWRKY4蛋白的活性与功能产生调控作用。亚细胞定位预测发现, 甘蔗ScWRKY4蛋白位于细胞核的概率最大, 为73.9%。从图3可以看出, 甘蔗基因编码蛋白包含一个从第166~223位的WRKY保守结构域, 确定该蛋白属于WRKY转录因子家族。采用NCBI中的BlastP在线程序, 查找ScWRKY4的同源氨基酸序列, 结果显示ScWRKY4蛋白与高粱SbWRKY57 (XP_002448267.2)、玉米ZmWRKY36 (NP_001151912.1)、II型少花古尔德草() DoWWRKY12 (OEL21317.1)、谷子() SiWRKY12 (XP_ 004956420.1)和水稻OsWRKY12 (XP_015635033.1)的相似性分别为88%、85%、82%、79%和68%, 其WRKY结构域的氨基酸序列高度保守, 而在其他位置的多肽序列由于物种的差异而表现出较大的变化(图4)。参考前人报道的WRKY蛋白[7,15,22], 将ScWRKY4蛋白序列与甘蔗、大麦、拟南芥、玉米中不同家族类别的WRKY蛋白序列构建系统进化树(图5), 表明ScWRKY4与拟南芥AtWRKY12和AtWRKY13等聚在同一分支上, 属于WRKY家族中的IIc亚家族。
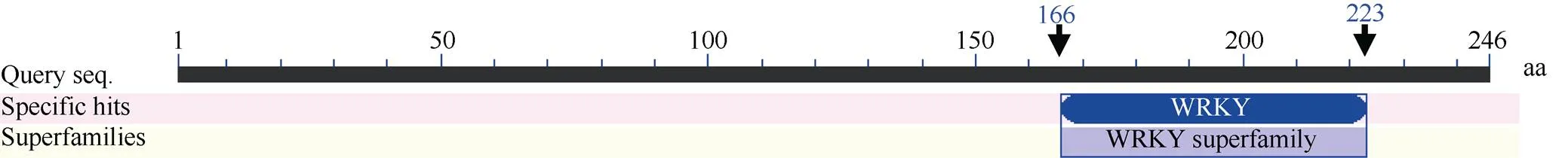
图3 甘蔗ScWRKY4蛋白的保守结构域分析
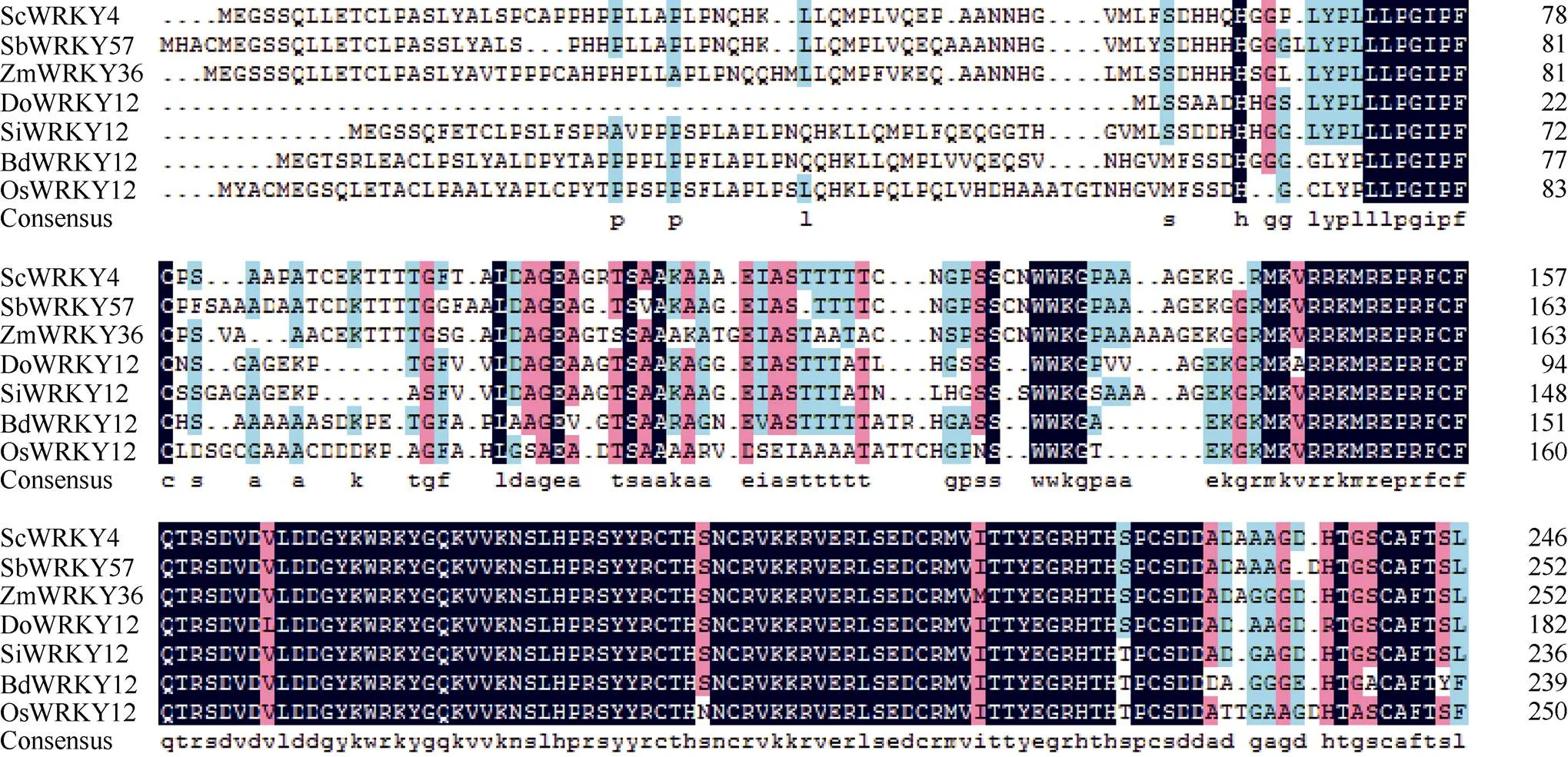
图4 甘蔗ScWRKY4与其他植物WRKY蛋白的氨基酸序列比对
高粱: SbWRKY57 (XP_002448267.2); 玉米: ZmWRKY36 (NP_001151912.1); II型少花古尔德草: DoWWRKY12 (OEL21317.1); 谷子: SiWRKY12 (XP_004956420.1); 二穗短柄草: BdWRKY12 (XP_014751531.1); 水稻: OsWRKY12 (XP_015635033.1)。
: SbWRKY57 (XP_002448267.2);: ZmWRKY36 (NP_001151912.1);: DoWWRKY12 (OEL21317.1);: SiWRKY12 (XP_004956420.1);: BdWRKY12 (XP_014751531.1);OsWRKY12 (XP_015635033.1).

图5 ScWRKY4蛋白与甘蔗及其他植物WRKY蛋白家族的系统进化树
2.3 甘蔗ScWRKY4基因的亚细胞定位分析
亚细胞定位表达载体35S::GFP和融合质粒35S::ScWRKY4::GFP分别用相同的内切酶H I和I进行双酶切验证, 结果如图6-A所示。采用农杆菌转化及注射本氏烟叶片的方法进行亚细胞定位观察, 阳性对照35S::GFP在细胞核、细胞质和质膜上都有绿色荧光分布, 而35S::ScWRKY4::GFP融合蛋白只在细胞核中有很强的绿色荧光(图6-B), 表明35S::ScWRKY4::GFP融合蛋白定位于细胞核中。
2.4 甘蔗ScWRKY4的自激活活性分析
重组质粒pGBKT7-经酶切验证及测序分析, 初步显示酵母融合表达载体构建成功(图7-A)。利用Y2HGold-GAL4酵母双杂交系统对ScWRKY4的自激活活性分析表明, 在SD/–Trp平板上, pGBKT7-与阳性对照pGADT7+ pGBKT7-p53和阴性对照pGBKT7一致, 均能长出菌落, 说明pGBKT7-重组质粒已成功转入Y2HGold中, 且GAL4-BD与ScWRKY4蛋白结合, 能成功表达色氨酸。在添加X-α-Gal的SD/–Trp平板上发现, 由于阳性对照中基因被激活, X-α-Gal检测显蓝色, 而pGBKT7-与阴性对照均不显蓝色, 说明ScWRKY4没有自激活活性。经过AbA抗性筛选, 阳性对照可正常生长, 且X-α-Gal检测显蓝色, 表明报告基因和基因都被激活, 而pGBKT7-和阴性对照一样, 均不能正常生长, 进一步说明ScWRKY4蛋白不具有自激活活性(图7-b)。
2.5 ScWRKY4基因的组织特异性表达分析
基因在甘蔗ROC22不同组织中的表达分析结果显示, 该基因在根、芽、叶和皮中的表达量差异不显著, 但在蔗肉中的表达量最高, 为对照蔗根中表达量的18.38倍(图8)。
2.6 ScWRKY4基因在不同外源胁迫下的表达特性分析
2.6.1基因在黑穗病菌侵染下的表达特性 黑穗病菌侵染24 h时, S基因在崖城05-179中的表达量显著下调至对照组的0.20倍, 随后稳定在同一表达水平。而在ROC22中, 该基因在受黑穗病菌侵染72 h内的表达量与对照组相比, 并无显著差异(图9)。

图6 甘蔗ScWRKY4亚细胞定位分析
(A) 35S::ScWRKY4::GFP亚细胞定位重组载体的酶切验证结果。1: 15000+2000 bp DNA marker; 2: 35S::GFP/I; 3:ORF PCR产物; 4: 35S::ScWRKY4::GFP/I; 5: 35S::ScWRKY4::GFP/IHI; 6: 100 bp ladder DNA marker。(B)农杆菌介导转化的及空载体注射本氏烟叶片48 h的亚细胞定位结果。本氏烟叶片表皮细胞被用于明场、绿色荧光、蓝色荧光、明场和绿色及蓝色荧光叠加后的图像分析; 白色箭头1、2和3分别表示质膜、细胞核和细胞质, 比例尺=25 μm; 35S::GFP: 携带空载pCAMBIA 1300-的农杆菌菌株; 35S::ScWRKY4::GFP: 携带重组载体pCAMBIA 1300--的农杆菌菌株; DAPI: 4’,6-二脒基-2-苯基吲哚。
(A) The enzyme digestion to identify the insert-integrated subcellular localization expression vector 35S::ScWRKY4::GFP. 1: 15000+2000 bp DNA marker; 2: 35S::GFP/I; 3:ORF PCR product; 4: 35S::ScWRKY4::GFP/I; 5: 35S::ScWRKY4::GFP/I+HI; 6: 100 bp ladder DNA marker. (B) Subcellular localizations of themediated transformation ofand empty vector inleaves 48 h after infiltration. The epidermal cells ofwere used for taking images of visible light, green fluorescence, blue fluorescence, merged visible light and green fluorescence and blue fluorescence; white arrows 1, 2, and 3 indicate plasma membrane, nucleus and cytoplasm, respectively; scale bar = 25 μm; 35S::GFP: thestrain carrying the empty vector pCAMBIA 1300-; 35S::ScWRKY4::GFP: thestrain carrying the recombinant vector pCAMBIA 1300--; DAPI: 4’,6-diamidino-2-phenylindole.
2.6.2基因在不同植物激素和非生物胁迫下的表达特性 在SA和MeJA处理下,基因的表达水平均高于对照组, 其中SA处理12 h时达到峰值, 为对照组的2.69倍, MeJA处理12 h时达到峰值, 为对照组的2.38倍。ABA胁迫下,基因的表达量在各个时间点存在显著变化, 0.5 h时该基因显著上调至对照组的1.59倍, 3 h时恢复到对照组水平, 6 h时达到峰值且为对照组的2.87倍, 随后在24 h时表达量有所下调, 但仍高于对照(1.26倍)。经NaCl处理,基因的表达量随胁迫时间的延长而呈现出逐渐上升的趋势, 在24 h时达到峰值, 其表达量为对照组的4.91倍。PEG处理下,基因的表达量在3 h时达到高峰, 为对照组12.10倍, 而在该时间点前后, 该基因的表达水平与对照组相比没有明显差异(图10)。总体上看,基因对SA、MeJA、ABA、NaCl和PEG胁迫都有一定程度的应答。
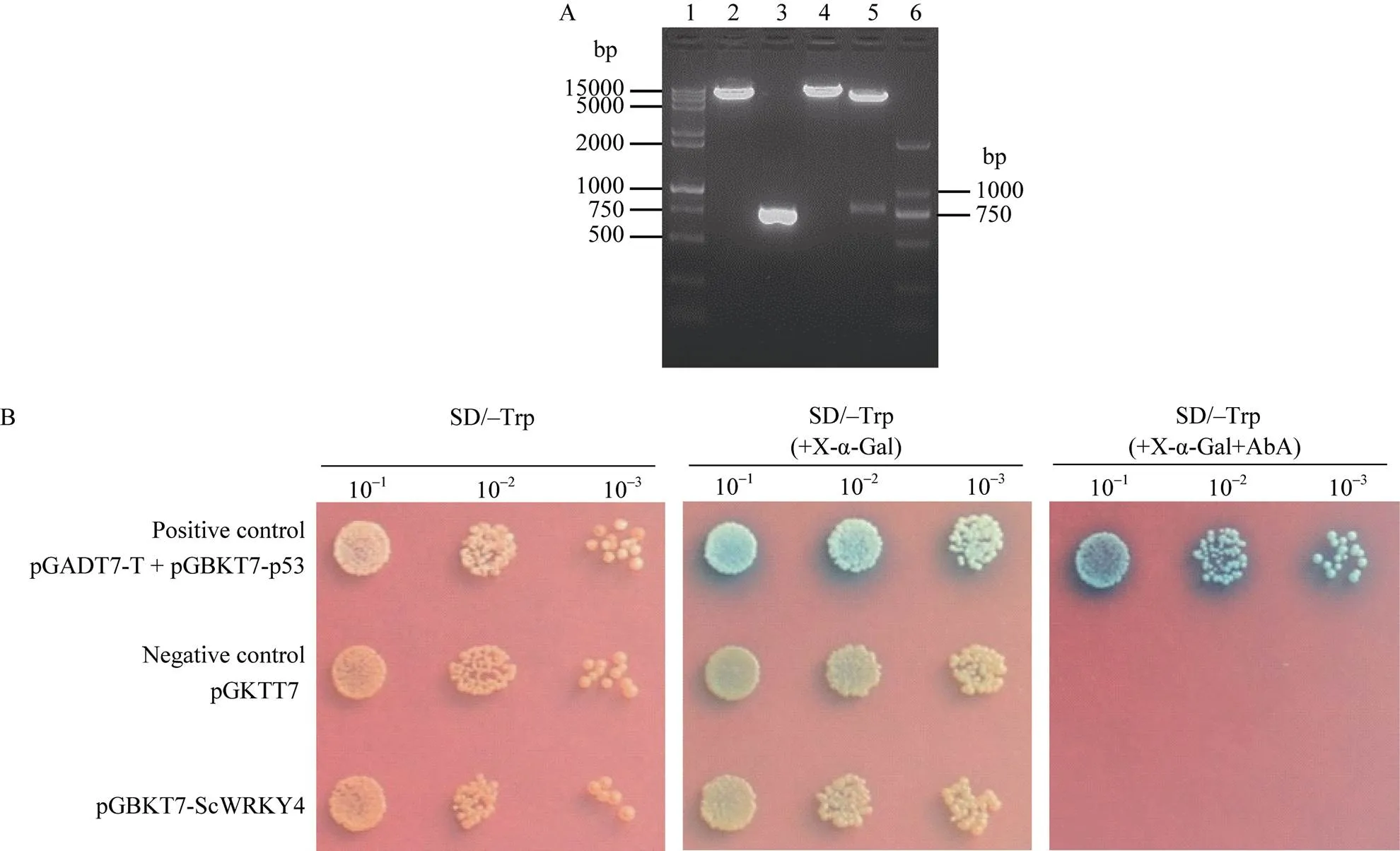
图7 甘蔗ScWRKY4转录激活活性分析
(A) pGBKT7-重组载体的酶切验证结果。1: 15000+2000 bp DNA marker; 2: pGBKT7/I; 3:ORF PCR产物; 4: pGBKT7-I; 5: pGBKT7-IHI; 6: 2000 bp ladder DNA marker。(B) ScWRKY4转录激活活性验证。SD/–Trp: 色氨酸营养缺陷型平板培养基; SD/–Trp (+ X-α-Gal): 色氨酸营养缺陷型平板培养基(添加5-溴-4-氯-3-吲哚-α-D-半乳糖苷); SD/–Trp (+ X-α-Gal+AbA): 色氨酸营养缺陷型平板培养基(添加5-溴-4-氯-3-吲哚-α-D-半乳糖苷和金担子素A)。
(A) The enzyme digestion to identify the insert-integrated vector pGBKT7. 1: 15000+2000 bp DNA marker; 2: pGBKT7/I; 3:ORF PCR product; 4: pGBKT7-I; 5: pGBKT7-IHI; 6: 2000 bp ladder DNA marker. (B) The test of transactivation activity assay of ScWRKY4. SD/–Trp: synthetic dropout medium plate; SD/–Trp (+ X-α-Gal): synthetic dropout medium plate without tryptophan (plus 5-bromo-4-chloro-3-indoxyl-α-D-galactopyranoside); SD/–Trp (+ X-α-Gal+ AbA): synthetic dropout medium plate without tryptophan (plus 5-bromo-4-chloro-3-indoxyl-α-D-galactopyranoside and aureobasidin A).
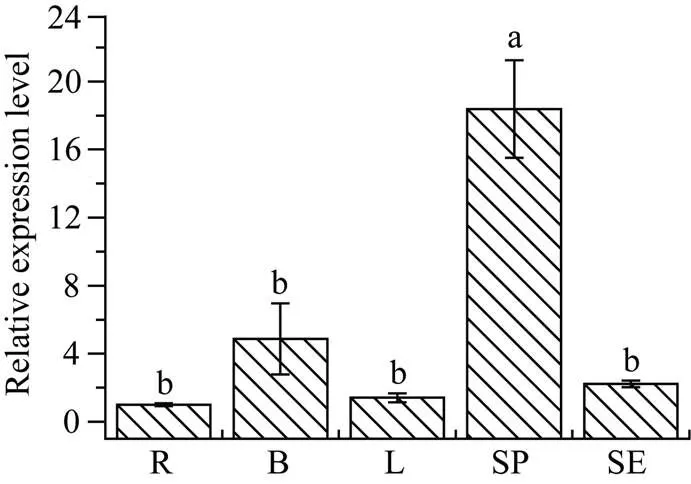
图8 ScWRKY4基因在甘蔗ROC22组织中的表达情况
柱上不同的小写字母代表显著性的差异(≤ 0.05);= 3; 内参基因为甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,); R: 蔗根; B: 蔗芽; L: 蔗叶; SP: 蔗肉; SE: 蔗皮。
Bars superscripted by different lowercase letters are significantly different (≤ 0.05);= 3; Glyceraldehyde-3-phosphate dehydrogenase () was used as a reference gene; R: root; B: bud; L: leaf; SP: stem pith; SE: stem epidermis.
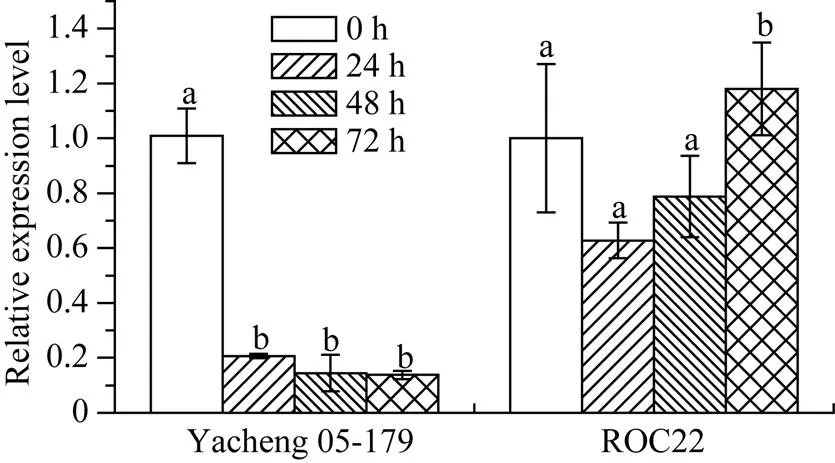
图9 甘蔗ScWRKY4基因在黑穗病侵染下的表达情况
Yacheng 05-179: 甘蔗抗黑穗病品种; ROC22: 甘蔗感黑穗病品种。柱上不同的小写字母代表显著性的差异(≤ 0.05);= 3; 内参基因为甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,)。
Yacheng 05-179: smut-resistant sugarcane variety; ROC22: smut-susceptible sugarcane variety. Bars superscripted by different lowercase letters are significantly different (≤ 0.05);= 3; Glyceraldehyde-3-phosphate dehydrogenase () was used as a reference gene.
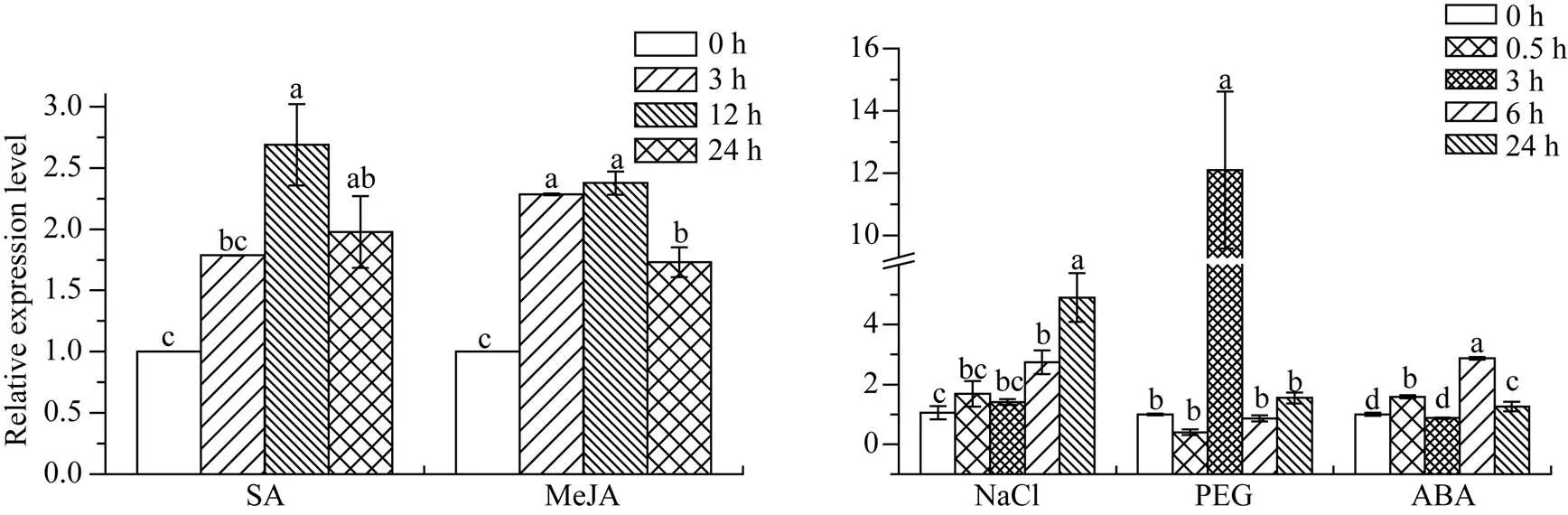
图10 甘蔗ScWRKY4基因在不同植物激素和非生物胁迫下的表达水平
柱上不同的小写字母代表显著性的差异(≤ 0.05);= 3; 内参基因为甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,); SA: 水杨酸(5 mmol L–1); MeJA: 茉莉酸甲酯(25 μmol L–1); NaCl: 氯化钠(模拟盐胁迫) (250 mmol L–1); PEG: 聚乙二醇(模拟干旱胁迫) (25.0%); ABA: 脱落酸(100 μmol L–1)。
Bars superscripted by different lowercase letters are significantly different (≤ 0.05);= 3; Glyceraldehyde-3-phosphate dehydrogenase () was used as a reference gene; SA: salicylic acid (5 mmol L–1); MeJA: methyl jasmonate (25 μmol L–1); NaCl: sodium chloride (simulating salt stress) (250 mmol L–1); PEG: polyethylene glycol (simulating drought treatment) (25.0%); ABA: abscisic acid (100 μmol L–1).
3 讨论
甘蔗是食糖总产占比最大的糖料作物, 同时也是可再生能源作物[28]。但是甘蔗在生产上常受到各种病害以及低温、干旱、高盐等非生物环境的影响, 导致其生物产量和质量的下降, 挖掘抗逆相关基因, 有助于利用转基因技术定向改良甘蔗品种的抗性[29]。当植物处于逆境环境时, 可以通过体内功能基因的表达变化来适应环境, 而转录因子对功能基因的表达和修饰起着重要的调控作用[4,6]。目前, 在甘蔗中, 与生物和非生物胁迫相关的转录因子及其相关基因已有报道[30]。其中WRKY转录因子的研究仅有一例-报道[19]。
WRKY转录因子包含WRKY蛋白结构域, 由60个左右的氨基酸残基组成, 可与DNA域结合, 且其N-末端有一个高度保守的核心序列WRKYGQK, C-末端则包含有锌指结构域[3]。根据WRKY蛋白所含有的WRKY结构域的个数和锌指结构的特点, 可将WRKY家族分为三类, 不同亚类可能反映其不同的功能, 其中第I类包含有2个WRKY结构域和锌指结构C2H2(Cx4-5Cx22-23HxH); 第II类含1个WRKY结构域和锌指结构C2H2(Cx4-5Cx22-23HxH), 根据其WRKY保守域外的保守结构序列特征, 又分IIa-IIe共5个亚类; 第III类含1个WRKY结构域, 锌指结构类型为C2HC (Cx7Cx23HxH)[30]。研究发现, 高度保守七肽序列WRKYGQK也并非完全保守, 如水稻中的高度保守的WRKY结构域, 共有9种变异类型, 其中较常见的为WRKYGEK和WRKYGKK[31]。已报道的甘蔗Sc-WRKY蛋白含有一个WRKY结构域, 且存在变异, 为WRKYGKK类型, 锌指结构为Cx4Cx23HxH[19]。在本研究中,基因编码蛋白与Sc-WRKY的氨基酸序列一致性仅为24.65%, 但其WRKY保守结构域七肽序列为普遍存在的WRKYGQK类型, 锌指结构与Sc-WRKY相同, 系统进化树分析显示两者都属于第IIc类WRKY转录因子, 推测Sc-WRKY和ScWRKY4可能具有某些相似的功能。
蛋白质的生物学功能与其在细胞中的定位紧密相关。向小华等[32]对普通烟草中的164个NtWRKY转录因子的亚细胞定位进行了预测分析, 发现其中有118个定位于细胞核, 主要为第I类和第II类WRKY家族成员, 而第III类家族成员中, 有77.4%定位于细胞质。亚细胞定位预测分析发现, ScWRKY4蛋白位于细胞核的概率最大, 为73.9%。本研究在本氏烟草叶片中对甘蔗ScWRKY4进行亚细胞定位分析, 发现其定位在细胞核, 与生物学软件及向小华等[32]的预测结果一致, 说明ScWRKY4可能作为核蛋白发挥作用。大部分WRKY转录因子的典型特征之一是具有转录激活活性, 但有些WRKY转录因子并不具有转录激活活性, 如大豆GmWRKY7、GmWRKY8、GmWRKY13和GmWRKY15转录因子在酵母中都无转录激活活性[33]。香蕉() MaWRKY11蛋白有2个WRKY保守结构域, N端具有转录自激活活性, 而C端没有[34]。没有自激活活性的转录因子序列可作为诱饵, 在cDNA文库中筛选与之互作的功能基因, 有助于进一步分析转录因子在响应外界信号过程中的作用。GmWRKY7能够与其下游功能基因的启动子相结合, 通过抑制基因的表达来影响大豆对铝毒害的响应[33]。除此之外, WRKY不仅作为转录因子起作用, 还可能具有信号传递功能, 参与到植物对外界胁迫信号的传递过程中[35]。本研究通过酵母体内转录激活试验, 发现ScWRKY4转录因子不具有自激活活性, 该研究结果有待在植物系统上作进一步鉴定, 关于ScWRKY4转录因子对下游基因的表达调控作用仍有待研究。
前人研究表明, 普通烟草中的基因, 大多数在根、茎和叶等不同组织中具有不同程度的表达[32]。本研究发现,在甘蔗ROC22的根、芽、叶、肉、皮中均有表达, 且在蔗肉中的表达量最高, 说明在甘蔗中的表达具有组成型表达特性。WRKY转录因子在植物对生物和非生物胁迫的应答反应中具有重要作用。有些WRKY转录因子在植物对病原菌等生物侵染的基础防御过程中起负调节作用, 如拟南芥中过表达或, 使得与抗病防御相关的基因的表达量下调[36]。Liu等[19]发现-基因在FN22中的表达受黑穗病菌的诱导, 说明-基因在对黑穗病菌侵染的防御反应中具有积极作用。本研究中的与-同属于WRKY转录因子家族中的第IIc类, 然而, 研究发现, 受黑穗病菌侵染后,基因在抗病甘蔗材料崖城05-179中表现出下调的趋势, 在感病品种ROC22中不受该病原菌诱导表达, 推测可能不参与甘蔗对黑穗病的抗性反应或在该防御系统中起负调控作用, 暗示隶属于相同基因亚族的与-, 可能在响应黑穗病菌胁迫方面存在功能的分化。基因在SA和MeJA处理下的表达量均高于对照水平, 且该基因均能被ABA、NaCl和PEG胁迫诱导表达。甘蔗-基因也受SA、NaCl、PEG等诱导表达[19]。研究结果表明,可能参与了甘蔗对盐胁迫和干旱胁迫的信号传导过程。同样, 水稻的表达水平在PEG、NaCl和ABA胁迫处理后均可提高, 并通过ABA途径提高对渗透胁迫的耐受能力[37]。拟南芥在ABA处理下诱导表达, 并参与到由ABA介导的信号途径中,积极响应对干旱的耐受性[38]。
4 结论
从甘蔗ROC22中成功分离到1个基因, 该基因长1265 bp, 包含1个741 bp的ORF, 编码246个氨基酸, 含1个WRKYGQK保守结构域和C2H2(CX4CX23HXH)锌指结构域, 属于WRKY转录因子家族的IIc亚类。ScWRKY4蛋白为碱性的不稳定亲水性蛋白, 不存在信号肽和跨膜结构域; 其蛋白二级结构元件缺少β螺旋结构; 包含有22个磷酸化位点。ScWRKY4被定位于细胞核, 不具有自激活活性。基因表达量在蔗肉中最高, 而在其他组织中差异不大; 该基因在甘蔗抗病品种崖城05-179与黑穗病菌互作过程中下调表达, 在感病品种ROC22中的表达水平较为稳定, 但在外源激素SA、MeJA和ABA以及非生物胁迫因子NaCl和PEG处理条件下均被诱导上调表达, 暗示基因不参与甘蔗对黑穗病的抗性反应或在该防御系统起负调控作用, 并且有可能参与到甘蔗对盐胁迫及干旱胁迫的信号传导过程中。
[1] Porto M S, Pinheiro M P N, Batista V G L, Santos R C D, Filho P A M, Lima L M. Plant promoters: an approach of structure and function., 2014, 56: 38–49
[2] 李田, 孙景宽, 刘京涛. 植物转录因子家族在耐盐抗旱调控网络中的作用. 生命科学, 2015, 27: 217–227 Li T, Sun J K, Liu J T. Role of different transcription factor families in the regulatory networks of drought and salinity tolerance in plants., 2015, 27: 217–227 (in Chinese with English abstract)
[3] Ishiguro S, Nakamura K. Characterization of a cDNA encoding a novel DNA-binding protein,, that recognizes SP8 sequences in the 5’ upstream regions of genes coding for sporamin and beta-amylase from sweet potato., 1994, 244: 563–571
[4] Eulgem T, Somssich I E. Networks of WRKY transcription factors in defense signaling., 2007, 10: 366–371
[5] Song Y, Ai C R, Jing S J, Yu D Q. Research progress on function analysis of ricegenes., 2010, 17: 60–72
[6] Pandey S P, Somssich I E. The role of WRKY transcription factors in plant immunity., 2009, 150: 1648–1655
[7] Mangelsen E, Kilian J, Berendzen K W, Kolukisaoglu Ü H, Harter K, Jansson C, Wanke D. Phylogenetic and comparative gene expression analysis of barley () WRKY transcription factor family reveals putatively retained functions between monocots and dicots., 2008, 9: 194
[8] Wei K F, Chen J, Chen Y F, Wu L J, Xie D X. Molecular phylogenetic and expression analysis of the complete WRKY transcription factor family in maize., 2012, 19: 153–164
[9] Rushton P J, Torres J T, Parniske M, Parniske M, Wernert P, Hahlbrock K, Somssich L M. Interaction of elicitor-induced DNA-binding proteins with elicitor response elements in the promoters of parsleygenes., 1996, 15: 5690–5700
[10] Dong J, Chen C H, Chen Z X. Expression profiles of thegene superfamily during plant defense response., 2003, 51: 21–37
[11] Liu D L, Leib K, Zhao P Y, Kogel K H, Langen G. Phylogenetic analysis of barley WRKY proteins and characterization ofand repressors of the pathogen-inducible gene., 2014, 289: 1331–1345
[12] Ramamoorthy R, Jiang S Y, Kumar N, Venkatesh P N, Ramachandran S. A comprehensive transcriptional profiling of thegene family in rice under various abiotic and phytohormone treatments., 2008, 49: 865–879
[13] Qiu D, Xiao J, Xie W B, Liu H B, Li X H, Xiong L Z, Wang S P. Rice gene network inferred from expression profiling of plants overexpressing, a positive regulator of disease resistance., 2008, 1: 538–551
[14] Peng Y, Bartley L E, Chen X W, Dardick C, Chern M, Ruan R, Canlas P E, Ronald P C.is a negative regulator of basal and-mediated defense againstpv.in rice., 2008, 1: 446–458
[15] 魏晓爱, 姚文静, 姜廷波, 周博如. 拟南芥基因家族应答非生物胁迫基因的鉴定. 东北林业大学学报, 2016, 44(10): 45–48 Wei X A, Yao W J, Jiang T B, Zhou B R. Identification ofgene in response to abiotic stress fromtranscirption factor gene family of., 2016, 44(10): 45–48 (in Chinese with English abstract)
[16] Wu J, Chen J B, Wang L F, Wang S M. Genome-wide investigation of WRKY transcription factors involved in terminal drought stress response in common bean., 2017, 8: 380
[17] Jiang Y Z, Duan Y J, Jia Y, Ye S L, Zhu J R, Zhang F Q, Lu W X, Fan D, Luo K M. Genome-wide identification and characterization of theWRKY transcription factor family and analysis of their expression in response to biotic and abiotic stresses., 2014, 65: 6629–6644
[18] Zhang T, Tan D, Zhang L, Zhang X, Han Z. Phylogenetic analysis and drought-responsive expression profiles of thetranscription factor family in maize., 2017, 3: 99–108
[19] Liu J X, Que Y X, Guo J L, Xu L P, Wu J Y, Chen R K. Molecular cloning and expression analysis of a WRKY transcription factor in sugarcane., 2012, 11: 6434–6444
[20] 黄宁, 张玉叶, 凌辉, 罗俊, 吴期滨, 阙友雄. 甘蔗二氨基庚二酸异构酶基因的克隆与表达分析. 热带作物学报, 2013, 34: 2200–2208 Huang N, Zhang Y Y, Ling H, Luo J, Wu Q B, Que Y X. Cloning and expression analysis of a diaminopimelate epimerase gene in sugarcane., 2013, 34: 2200–2208 (in Chinese with English abstract)
[21] Que Y X, Su Y C, Guo J L, Wu Q B, Xu L P. A global view of transcriptome dynamics duringchallenge in sugarcane by RNA-Seq., 2014, 9: e106476
[22] Wei K, Chen J, Chen Y F, Wu L J, Xie D X. Multiple-strategy analyses ofsubgroups and functional exploration ofgenes in pathogen responses., 2012, 8: 1940–1949
[23] 苏亚春. 甘蔗应答黑穗病菌侵染的转录组与蛋白组研究及抗性相关基因挖掘. 福建农林大学博士学位论文, 福建福州, 2014 Su Y C. Transcriptomics and Proteomics of Sugarcane Response toInfection and Mining of Resistance-Related Genes. PhD Dissertation of Fujian Agriculture and Forestry University, Fuzhou, Fujian, China, 2014 (in Chinese with English abstract)
[24] Zhou X J, Yan S W, Sun C, Li S Z, Li J, Xu M Y, Liu X Q, Zhang S J, Zhao Q Q, Li Y, Fan Y L, Chen R M, Wang L. A maize jasmonate zim-domain protein, ZmJAZ14, associates with the JA, ABA, and GA signaling pathways in transgenic., 2015, 10: e0121824
[25] 阙友雄, 许莉萍, 徐景升, 张积森, 张木清, 陈如凯. 甘蔗基因表达定量PCR分析中内参基因的选择. 热带作物学报, 2009, 30: 274–278 Que Y X, Xu L P, Xu J S, Zhang J S, Zhang M Q, Chen R K. Reference gene selection in quantitative PCR analysis of gene expression in sugarcane., 2009, 30: 274–278 (in Chinese with English abstract)
[26] Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2–ΔΔCTmethod., 2001, 25: 402–408
[27] 苏亚春, 王竹青, 李竹, 刘峰, 许莉萍, 阙友雄, 戴明剑, 陈允浩. 甘蔗过氧化物酶基因的克隆与功能鉴定. 作物学报, 2017, 43: 510–521 Su Y C, Wang Z Q, Li Z, Liu F, Xu L P, Que Y X, Dai M J, Chen Y H. Molecular cloning and functional identification of peroxidase genein sugarcane., 2017, 43: 510–521 (in Chinese with English abstract)
[28] Yadav R L, Solomon S. Potential of developing sugarcane by-product based industries in India., 2006, 8: 104–111
[29] 付瑜华, 卢加举, 李向勇. 甘蔗主要病害抗病遗传育种研究进展. 生物技术通报, 2014, 45(2): 1–6 Fu Y H, Lu J J, Li X Y. Progress on disease-resistant breeding of major diseases in sugarcane., 2014, 45(2): 1–6 (in Chinese with English abstract)
[30] Eulgem T, Rushton P J, Robatzek S, Somssich I E. The WRKY superfamily of plant transcription factor., 2000, 5: 199–206
[31] Zhang Y J, Wang L J. The WRKY transcription factor superfamily: its origin in eukaryotes and expansion in plants., 2005, 5: 1–12
[32] 向小华, 吴新儒, 晁江涛, 杨明磊, 杨帆, 陈果, 刘贯山, 王元英. 普通烟草基因家族的鉴定及表达分析. 遗传, 2016, 38: 840–856 Xiang X H, Wu X R, Chao J T, Yang M L, Yang F, Chen G, Liu G S, Wang Y Y. Genome-wide identification and expression analysis of thegene family in common tobacco (L.).(Beijing), 2016, 38: 840–856 (in Chinese with English abstract)
[33] 彭俊楚. 大豆基因的克隆和功能研究. 华南农业大学博士学位论文, 广东广州, 2016 Peng J C. Isolation and Function Analysis of Soybeans. PhD Dissertation of South China Agricultural University, Guangzhou, Guangdong, China, 2016 (in Chinese with English abstract)
[34] 徐群刚, 邝健飞, 单伟, 陆旺金, 陈建业. 香蕉果实冷胁迫相关MaWRKY11转录因子的特性、互作蛋白筛选与鉴定. 热带亚热带植物学报, 2015, 23: 543–552 Xu Q G, Kuang J F, Shan W, Lu W J, Chen J Y. Characterization and interacting-protein identification of MaWRKY11 transcription factor related to cold stress from banana fruits., 2015, 23: 543–552 (in Chinese with English abstract)
[35] Jing J J, Ma S, Ye N H, Jing M, Cao J S, Zhang J H. WRKY transcription factors in plant responses to stresses., 2017, 59: 86–101
[36] Kim K C, Lai Z, Fan B, Chen Z.WRKY38 and WRKY62 transcription factors interact with histone deacetylase 19 in basal defense., 2008, 20: 2357–2371
[37] Song Y, Jing S J, Yu D Q. Overexpression of the stress-induced, improves osmotic stress tolerance in., 2009, 54: 4671–4678
[38] Ren X, Chen Z, Liu Y, Zhang H R, Zhang M, Liu Q, Hong X H, Zhu J K, Gong Z Z. ABO3, a WRKY transcription factor, mediates plant responses to abscisic acid and drought tolerance in., 2010, 63: 417–429
Cloning and Expression Characteristic Analysis ofGene in Sugarcane
WANG Ling1, LIU Feng1, DAI Ming-Jian1, SUN Ting-Ting1, SU Wei-Hua1, WANG Chun-Feng1, ZHANG Xu1, MAO Hua-Ying1, SU Ya-Chun1,2,*, and QUE You-Xiong1,2,*
1Key Laboratory of Sugarcane Biology and Genetic Breeding (Fujian), Ministry of Agriculture / Fujian Agriculture and Forestry University, Fuzhou 350002, Fujian, China;2Key Laboratory of Crop Genetics and Breeding and Comprehensive Utilization, Ministry of Education / Fujian Agriculture and Forestry University, Fuzhou 350002, Fujian, China
WRKY is one of the specific transcriptional regulators in plants and widely involved in the response of plants to diverse biotic and abiotic stresses. In this study, based on our previous transcriptome data of sugarcane (spp.), we successfully cloned agene from sugarcane variety ROC22, named as(GenBank accession number MG852087). Sequence analysis showed that the full-length cDNA ofgene was 1265 bp, containing a 741 bp complete open reading frame and encoding 246 amino acids residues. ScWRKY4 protein had a conservative WRKYGQK domain and a zinc finger motif C2H2, belonging to IIc subgroup of WRKY transcription factor family. Bioinformatics analysis indicated that ScWRKY4 was an alkaline unstable hydrophilic protein with no signal peptide and transmembrane structure. The secondary structural element of ScWRKY4 protein was lack of beta helix structure. ScWRKY4 was located in the nucleus after transient expression in. The result of yeast hybridization experiment demonstrated that ScWRKY4 did not possess transcriptional activity. Real-time fluorescent quantitative PCR showed that the expression ofgene had no significant difference in root, leaf, bud and stem epidermis, while had the highest expression level in stem pith, which was 18.38 times higher than that in root. During inoculation with smut pathogen () at 0–72 h, the gene expression level ofwas down-regulated in sugarcane smut-resistant cultivar Yacheng 05-179, while was more stable in the susceptible one ROC22. The transcript ofwas up-regulated by exogenous hormone stresses of abscisic acid, salicylic acid, and methyl jasmonate, as well as by the abiotic treatments of sodium chloride and polyethylene glycol. These results suggest thatgene may not participate in sugarcane smut resistance, or only play a negative role in this defense reaction, but positively involve in the response mechanism of salt and drought resistance in sugarcane.
sugarcane; WRKY transcription factor; bioinformatics; subcellular localization; transcriptional activity; real-time flourescent quantitative PCR
2018-01-28;
2018-06-12;
2018-07-02.
10.3724/SP.J.1006.2018.01367
苏亚春, E-mail: syc2009mail@163.com; 阙友雄, E-mail: queyouxiong@126.com
E-mail: lingw2017@126.com
本研究由国家自然科学基金项目(31671752和31101196), 福建省杰出青年基金项目(2015J06006), 福建省高校杰出青年科研人才计划项目(苏亚春-2017)和国家现代农业产业技术体系建设专项(CARS-17)资助。
This study was supported by the National Natural Science Foundation of China (31671752, 31101196), the Natural Science Foundation of Fujian Province for Distinguished Young Scholars (2015J06006), the Research Funds for Distinguished Young Scientists in Fujian Provincial Department of Education (SYC-2017), and China Agriculture Research System (CARS-17).
URL:http://kns.cnki.net/kcms/detail/11.1809.S.20180630.1120.002.html
