玉米隐花色素CRY1b和CRY2基因转录丰度对不同光质处理的响应
2018-09-11李红丹樊晓聪陈士瞻燕郭游光霞杨宗举杨建平
李红丹 闫 蕾 孙 蕾 樊晓聪 陈士瞻 张 燕郭 林 游光霞 李 庄 杨宗举 苏 亮,* 杨建平,*
玉米隐花色素和基因转录丰度对不同光质处理的响应
李红丹1,2闫 蕾1,2孙 蕾1,2樊晓聪1,3陈士瞻1,3张 燕1,3郭 林1游光霞1李 庄1,2杨宗举1,2苏 亮1,*杨建平1,3,*
1中国农业科学院作物科学研究所, 北京 100081;2中国农业科学院研究生院, 北京 100081;3河南农业大学农学院, 河南郑州 450002
玉米株高、开花期、产量、品质等性状与环境中的光密切相关。隐花色素是一类蓝光和近紫外光的受体, 主要参与植物的光形态建成及动、植物的生物钟调控。通过研究玉米隐花色素基因对不同光处理的表达模式, 可为进一步研究其对玉米光形态建成的作用奠定基础。本研究采用RT-PCR技术克隆了玉米和基因; 利用生物信息学相关网站和软件对其编码蛋白的结构域及氨基酸进行了系统发育分析; 利用qRT-PCR分析了玉米自交系B73中和基因在不同组织、以及响应不同光质及长日照和短日照处理的转录丰度。研究发现, 玉米与拟南芥、水稻和小麦的CRY蛋白有相同的结构域及较高的氨基酸序列的一致性, 表明它们具有相似的功能。和基因主要在玉米的叶片中表达; 二者能迅速响应各种持续光质、黑暗到不同光质转换及长日照和短日照处理, 且在各种处理下的转录丰度均高于, 可能暗示在玉米中功能更强。以上研究结果表明,和基因均能有效地响应各种光质和光周期处理, 并在玉米的光形态建成中发挥重要作用。本研究为进一步探明和基因的功能及其在玉米品种改良中的应用提供了研究基础。
玉米; 隐花色素; 光形态建成; 光处理; 转录丰度
环境中光的辐照度、方向与光质能显著影响植物的株高、株型、产量、品质等[1-5]。光照不仅是植物进行光合作用的能量来源, 更能调控其生长与发育,如光形态建成等。在长期的进化中, 植物逐步形成三大类光受体, 光敏色素(phytochrome), 主要负责感知红光(波长为620~700 nm)和远红光(700~800 nm); 隐花色素(cryptochrome)和向光素(phototropin), 主要感知蓝光(380~500 nm)和UV-A (320~380 nm); UVR8, 能特异性地吸收UV-B (280~ 320 nm)[6-8]。其中, 隐花色素是一种分子量为70~80 kD的黄素类蛋白, 有氨基端PHR和羧基端CCE两个明显的功能域, 其氨基端的PHR非共价地结合生色团FAD和叶酸[6]。隐花色素是一种类光解酶的光受体, 参与了动物、植物乃至微生物中的光反应及生物钟的调控[8]。通过研究在不同条件下玉米隐花色素的表达模式, 可为进一步研究其对玉米光形态建成的作用, 进而认知玉米光的反应能力奠定基础。
在拟南芥[9-10]、番茄[2-3,11]、苹果[12]、水稻[13-14]、豌豆[4,15]、蕨类[16]、苔藓[5,17]、大豆[18-19]、甘蓝型油菜[20]等植物中的研究表明, 隐花色素不仅控制下胚轴的伸长、光周期诱导的开花及生物钟的输出, 还参与了向性生长、顶端优势、细胞程序性死亡、种子休眠、根的发育、气孔开张等过程。模式植物拟南芥中存在3种类型的隐花色素, 即CRY1、CRY2和CRY3。在蓝光下, CRY1和CRY2负责调控植物的去黄化与开花期。在蓝光下, CRY1通过抑制COP1/SPA1的E3泛素连接酶活性, 导致正向调控因子HY5的积累, 从而促进光形态建成并抑制下胚轴的伸长, 而突变体则表现为下胚轴伸长且子叶生长缓慢[21-22]。CRY2在蓝光下与互作蛋白CIB结合促进开花,双突变体开花要晚于野生型[23]。CRY1在细胞核和细胞质中来回穿梭[24], CRY2在细胞核中行使功能[25], 而CRY3则主要作为光受体和单链DNA修复酶在线粒体和叶绿体中发挥作用[26]。番茄的隐花色素基因和能促进叶片中花青素苷和叶绿素的积累、促进腋分枝、缩短节间、延迟开花、促进果实中类黄酮和番茄红素的积累[2-3,11]。玉米隐花色素至少存在4个拷贝, 即、、和[27]。玉米基因在白光、红光、蓝光和远红光等条件下的转录丰度存在明显差异,基因对各种光质的响应则比较相似, 暗示着和存在功能上的分工, 可能参与玉米在红光和远红光下的光形态建成的调控[27]。
迄今, 双子叶模式生物拟南芥中隐花色素的研究已较为深入, 而玉米、水稻、马铃薯、小麦等禾本科单子叶作物中隐花色素的研究仅略有涉及, 对于隐花色素是否参与玉米的光形态建成以及是否调控玉米的光温反应也不清楚。通过比较玉米Z和基因响应不同光质及光周期处理的表达模式, 了解二者在不同条件下的表达差异, 比较二者响应光的能力, 可进一步丰富植物隐花色素基因的研究, 并可为作物的开花期、避荫性、株高、产量等重要农艺性状的改良提供指导意见。因此, 研究玉米和基因在不同光质及光周期处理下的表达模式对于了解玉米对光质及光周期的敏感性、开花机理及玉米的引种和高产栽培都具有重要的理论和实践意义。
1 材料与方法
1.1 试验材料及样品处理
1.1.1 试验材料 以玉米自交系B73的种子为试验材料(B73材料由中国农业科学院作物科学研究所李新海博士赠予)。以上材料适时取样,迅速置于液氮冻存,–80℃保藏备用。
1.1.2 组织特异性表达样品的准备 将B73的种子播种于自然条件下一定阶段后, 分别取幼苗的根、茎、叶、叶枕、叶鞘、花丝、花柄、雄花、苞叶和幼穗等部位为材料。
1.1.3 各种持续光质处理 将B73的种子放在28℃, 黑暗(Dk)、蓝光(B, 13 µmol m−2s−1)、白光(W, 17 µmol m–2s–1)、红光(R, 22.3 µmol m–2s–1)和远红光(FR, 0.25 µmol m–2s–1)等培养箱中, 培养13 d。取幼苗地上部为材料。
1.1.4 黑暗转换其他光质处理 将B73的种子放在28℃、黑暗中生长13 d。一批分别转入白光(W, 17 µmol m–2s–1)、红光(R, 22.3 µmol m–2s–11)以及蓝光(B, 13 µmol m–2s–1)等光质条件下, 并分别于0.25 h、0.5 h、1 h、2 h、4 h、8 h、12 h和24 h时对幼苗地上部分取样。另一批转入远红光(FR, 0.25 µmol m–2s–1)条件下并于0.16 h、0.25 h、0.5 h、1 h、1.5 h、2 h、3 h、4 h、6 h、8 h、10 h、12 h、24 h和24.08 h时对幼苗地上部取样。
1.1.5 长日照以及短日照处理 将B73种子分别放在28℃, 长日照培养箱(LD, 16 h光照/8 h黑暗)或者短日照培养箱(SD, 8 h光照/16 h黑暗)等条件下生长13 d, 每2 h取一次样, 光照和黑暗互相转换时, 在每个取样节点前5 min对幼苗地上部取样。
1.2 酶、试剂和载体
使用TaKaRa宝生物工程(大连)有限公司生产的PrimeSTAR HS酶和DNase I (RNase-free)酶, Invitrogen (USA)公司生产的TRIzol; Thermo Scientific公司生产的Revert Aid First Strand cDNA Synthesis Kit; TaKaRa宝生物工程(大连)有限公司生产的定量PCR试剂SYBR Premix ExII, 北京全式金生物技术有限公司生产的pEASY-Blunt-Simple载体。
1.3 RNA提取及cDNA合成
用TRIzol法提取各种处理条件下的玉米B73幼苗的总RNA, 用DNase I酶处理并以此为模板, 利用RevertAid First Strand cDNA Synthesis Kit并以Oligo-dT18引物将其反转录成单链cDNA, 保存于-80℃冰箱备用。
1.4 ZmCRY1b和ZmCRY2基因的克隆
于NCBI 网站(http://www.ncbi.nlm.nih.gov/)查找(ZM02G13620)和(ZM09G 09240)序列并设计引物(表1), 以黑暗条件下生长13 d的B73幼苗的cDNA为模板, 经PCR扩增后连接pEASY-Blunt-Simple载体, 经过菌落PCR和双酶切鉴定后, 交北京奥科鼎盛生物科技有限公司测序。PCR程序为94℃ 5 min; 98℃ 10 s, 58℃ 15 s, 72℃ 2 min, 25个循环; 72℃ 10 min。
1.5 玉米实时荧光定量RT-PCR(qRT-PCR)分析
以不同处理条件下的玉米自交系B73的cDNA第1链为模板, 玉米基因为内参基因, 用软件Primer Premier 5.0设计RT-PCR引物(表2)。使用Roche 480荧光定量PCR仪, PCR程序为95℃ 30 s; 95℃ 5 s, 60℃ 20 s, 72℃ 10 s, 50个循环; 60~95℃绘制溶解曲线。采用的定量PCR试剂为SYBR Premix ExII, 用2–ΔΔCT的方法计算实验结果[28], 经过3次独立的生物学重复后, 计算其标准差。
1.6 ZmCRY1b和ZmCRY2蛋白的序列比对、结构域分析与系统发育分析
使用NCBI网站(http://blast.ncbi.nlm.nih.gov/ Blast.cgi)推导CRY蛋白的氨基酸序列; ProtParam网站(http://web.expasy.org/protparam)分析蛋白的分子量、分子式和等电点等性质; DNAMAN (Version 8)对其进行系列比对和系统发育分析; SMART网站(http://smart.embl-heidelberg.de/)分析蛋白质的结构域。
2 结果与分析
2.1 ZmCRY蛋白和禾本科其他植物CRY蛋白有相同的结构域和较高的同源性
通过RT-PCR方法得到和基因的全长cDNA序列, 其ORF分别含有2067和1965个核苷酸残基, 编码蛋白质的分子式分别为C3464H5295N997O1009S18和C3316H5116N894O990S28, 分别含有688个和654个氨基酸残基, 蛋白质分子量分别为77.6 kD和74.2 kD, 等电点分别为5.89和5.54。
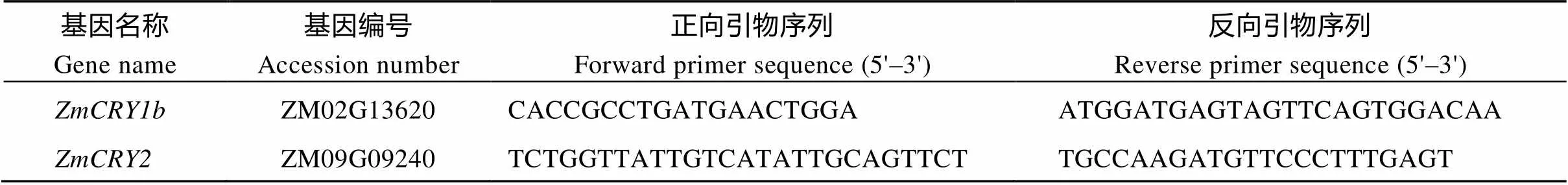
表1 基因克隆所用引物
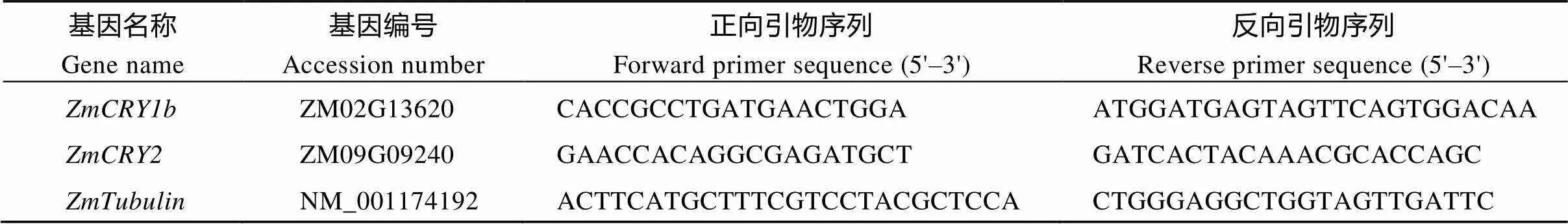
表2 qRT-PCR所用引物
利用NCBI网站和SMART网站对拟南芥及小麦、水稻和玉米的CRY蛋白进行结构域分析发现, 四者的CRY1蛋白都包含一个位于N端的PHR (Photolyase Homologous Region)结构域(由1个DNA photolyase结构域以及1个FAD binding 7 结构域组成)和1个位于C端的CCE (Cryptochrome C-terminal Extension)结构域(图1); 而CRY2蛋白则仅包含1个PHR结构域(图2)。玉米CRY1和CRY2蛋白结构域的不同暗示着其功能有异。用NCBI网站获得模式植物拟南芥()、玉米(L.)、小麦()、水稻(L.)、大豆()、番茄()等植物的CRY蛋白序列, 用DNAMAN进行系统发育分析发现, ZmCRY1b蛋白与拟南芥、小麦和水稻CRY1蛋白序列的一致性分别为58.6%、72.3%和82.9%; ZmCRY2蛋白与拟南芥、小麦和水稻CRY2蛋白序列的一致性分别为50.2%、73.5%和73.7% (图3)。说明ZmCRY蛋白与禾本科其他植物的一致性更高, 预示它们有更相似的功能。
2.2 ZmCRY1b和ZmCRY2器官特异性表达分析
基因的表达具有时空特异性, 为研究隐花色素基因在玉米生长和发育过程中的作用, 我们通过qRT-PCR的方法分析了和两个基因在玉米的根、茎、叶、叶枕、叶鞘、花丝、花柄、雄花、苞叶、幼穗等器官中表达丰度的差异。将基因在根中的表达丰度设为1, 并以此为对照。结果发现基因在花丝、幼穗、花柄和雄花和中的表达量较低(为根部丰度的0.1~0.4倍,<0.01), 在茎和苞叶中表达丰度与其根中的表达丰度相似(1.3~1.9倍), 在叶枕和叶鞘中表达丰度稍高(根中表达量的2.5~3.2倍, 差异极显著), 而在叶片中的表达丰度达到最大值, 是根中表达量的31.3倍(图4)。基因在幼穗和苞叶中的表达丰度也比较高(1.1~1.4倍), 且叶片中表达丰度最高(达到根中的10.4倍,<0.01), 而在其他器官中的表达丰度仅为根中的0.3~0.9倍, 具有显著差异(图4)。说明叶片是和基因在玉米中的主要作用部位, 且的表达丰度普遍高于, 暗示着在玉米中起着更重要的作用。

图1 玉米与拟南芥、小麦和水稻的CRY1蛋白等常见作物的氨基酸序列比对及结构域分析
使用NCBI网站和DNAMAN对其进行序列比对; 使用SMART网站对其进行结构域分析。AtCRY1: 拟南芥CRY1, AAB28724; OsCRY1a: 水稻CRY1a, BAB70686; OsCRY1b: 水稻CRY1b, BAB70688; TaCRY1a: 小麦CRY1a, ABX58028; ZmCRY1b: 玉米CRY1b, ZM02G13620。图中黑色、深灰色、浅灰色及白色分别代表一致性为100%、75%、50%和0。
Multiple sequence alignments at amino acid levels were analyzed by NCBI and DNAMAN, and the function domains were analyzed by SMART. AtCRY1:CRY1, AAB28724; OsCRY1a:CRY1a, BAB70686; OsCRY1b:CRY1b, BAB70686; TaCRY1a:CRY1a, ABX58028; ZmCRY1b:CRY1b, ZM02G13620. Black, dark grey, light grey, and white in the picture stand for 100%, 75%, 50%, and 0 of consistency, respectively.
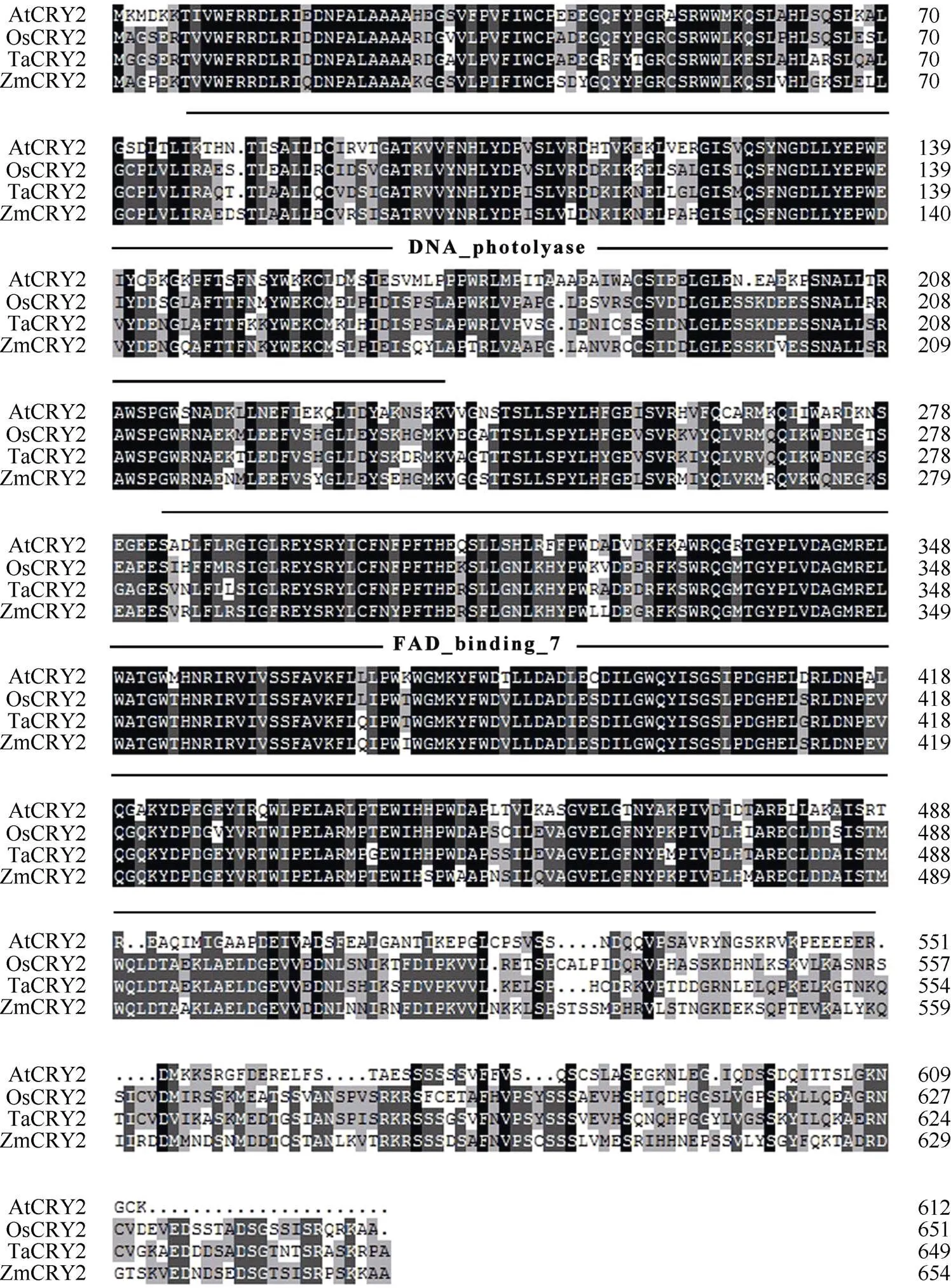
图2 玉米与拟南芥、小麦、水稻等常见作物的CRY2蛋白的氨基酸序列比对和结构域分析
使用NCBI网站和DNAMAN对其进行序列比对; 使用SMART网站对其进行结构域分析。AtCRY2: 拟南芥CRY2, AT1G04400.2; OsCRY2: 水稻CRY2, CAC82538.1; TaCRY2: 小麦CRY2, ABX58030.1; ZmCRY2: 玉米CRY2, ZM09G09240。图中黑色、深灰色、浅灰色及白色分别代表一致性为100%、75%、50%和0。
Multiple sequence alignments at amino acid levels were analyzed by NCBI and DNAMAN, and the function domains were analyzed by SMART. AtCRY2:CRY2, AT1G04400.2; OsCRY2:CRY2, CAC82538.1; TaCRY2:CRY2, ABX58030.1; ZmCRY2:CRY2, ZM09G09240. Black, dark grey, light grey, and white in the picture stand for 100%, 75%, 50%, and 0 of consistency, respectively.
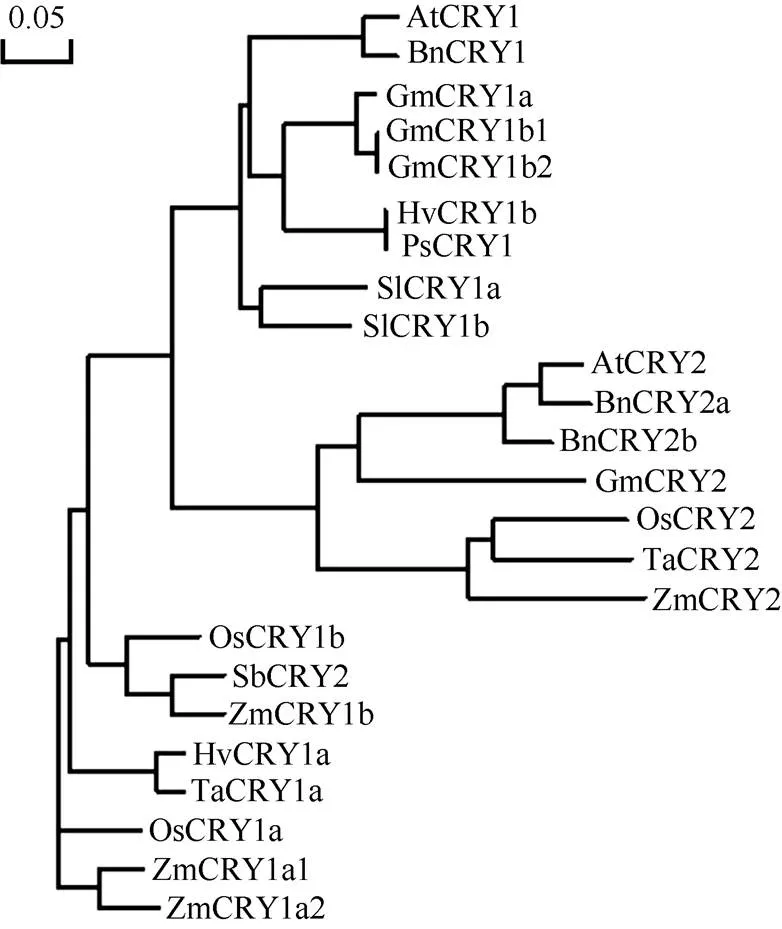
图3 玉米与拟南芥、小麦和水稻等常见作物的CRY蛋白在氨基酸水平的系统发育分析
使用NCBI网站获得全长氨基酸系列, 使用DNAMAN对其进行系统发育分析。AtCRY1: 拟南芥, AAB28724; AtCRY2: 拟南芥, AT1G04400.2; BnCRY1: 甘蓝型油菜, CAG28805; BnCRY2a: 甘蓝型油菜, AEA29690.1; BnCRY2b: 甘蓝型油菜油菜, AEA29691.1; GmCRY1a: 大豆, DQ401046; GmCRY1b1: 大豆, AB498929; GmCRY1b2: 大豆, AB498930; GmCRY2: 大豆, XP_006588364.1; OsCRY1a: 水稻, BAB70686; OsCRY1b: 水稻, BAB70688; OsCRY2: 水稻CAC82538.1; HvCRY1a: 大麦, ABB13328; HvCRY1b: 大麦, ABB13331; TaCRY1a: 小麦, ABX58028; TaCRY2: 小麦, ABX58030.1; PsCRY1: 豌豆, AAS79662; SbCRY2: 高粱, AAN37909; SlCRY1a: 番茄, AAD44161; SlCRY1b: 番茄, AAL02092; ZmCRY1a1: 玉米, ZM05G31560; ZmCRY1a2: 玉米, ZM04G17060; ZmCRY1b: 玉米, ZM02G13620; ZmCRY2: 玉米, ZM09G09240。
The amino acid sequences were obtained from the NCBI and the neighbor-joining tree was constructed by the DNAMAN. AtCRY1;CRY1, AAB28724; AtCRY2:CRY2, AT1G04400.2; BnCRY1:CRY1, CAG28805; BnCRY2a:CRY2a, AEA29690.1; BnCRY2b:CRY2b, AEA29691.1; GmCRY1a:CRY1a, DQ401046; GmCRY1b1:CRY1b1, AB498929; GmCRY1b2:CRY1b2, AB498930; GmCRY2:CRY2, XP_006588364.1; OsCRY1a:Group CRY1a, BAB70686; OsCRY1b:Group CRY1bBAB70688; OsCRY2:Group CRY2, CAC82538.1; HvCRY1a:subspCRY1a, ABB13328; HvCRY1b:subspCRY1b, ABB13331; TaCRY1a:CRY1a, ABX58028; TaCRY2:CRY2, ABX58030.1; PsCRY1:CRY1, AAS79662; SbCRY2:CRY2, AAN37909; SlCRY1a:CRY1a, AAD44161; SlCRY1b:CRY1b, AAL02092; ZmCRY1a1:CRY1a1, ZM05G31560; ZmCRY1a2:CRY1a2, ZM04G17060; ZmCRY1b:CRY1b, ZM02G13620; ZmCRY2:CRY2, ZM09G09240.
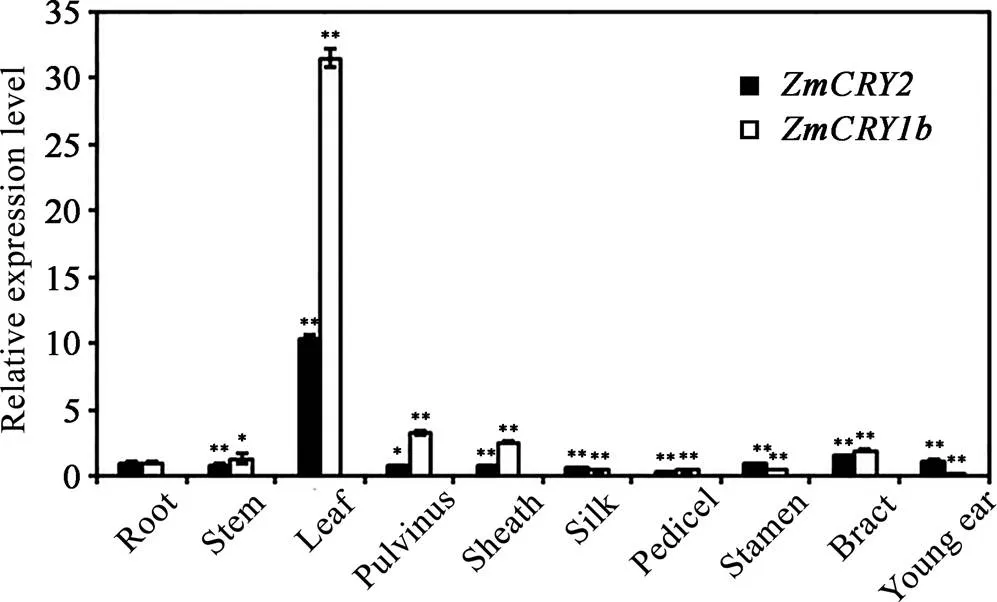
图4 ZmCRY1b和ZmCRY2基因在不同器官中的相对表达水平分析
分别取于自然条件下生长一定阶段的B73幼苗的不同器官(根、茎、叶、雄花、叶枕、叶鞘、花丝、幼穗、花柄和苞叶)为材料用于qRT-PCR分析。以根中的的转录丰度为对照, 将该的比值设为1。柱状图显示了在3次独立的生物学重复下比值的平均值, 其中, 误差线代表了标准差。用星号表示差异的显著性, *< 0.05, **< 0.01。
Different tissues of maize inbred line B73, including root, stem, leaf, stamen, pulvinus, sheath, pedicel, young ear, pistil and bract, were harvested for qRT-PCR assays. The transcription abundance ofin the root was set as the control, and the ratio ofin the root was set as 1. Each column shows the mean expression ofof three biological repeats. Error bars indicate the standard deviation. *< 0.05, **< 0.01.
2.3 ZmCRY1b和ZmCRY2基因转录丰度对不同持续光质处理的响应
为比较和对各种光质响应的差异, 采用qRT-PCR的方法分析了玉米B73幼苗在黑暗(Dk)和持续远红光(FR)、红光(R)、蓝光(B)、白光(W)等条件下和的表达丰度。将在黑暗条件下的表达量设为1, 并以此为对照。结果发现,和基因对各种持续光质均有较强烈的响应, 尤其是对持续白光的响应最为强烈, 分别为黑暗下基因表达量的48.0倍和3.7倍(图5,<0.01)。和基因在持续的远红光下的表达丰度也比较高, 分别为黑暗下丰度的29.3倍和2.4倍(差异极显著)。在蓝光和红光下,接近自身黑暗下的丰度, 而则为黑暗下的11.9倍和14.7倍(<0.01)。结果说明玉米和在远红光、红光、蓝光和白光中均有作用, 而的表达丰度普遍高于, 暗示在各种持续光质下更重要。
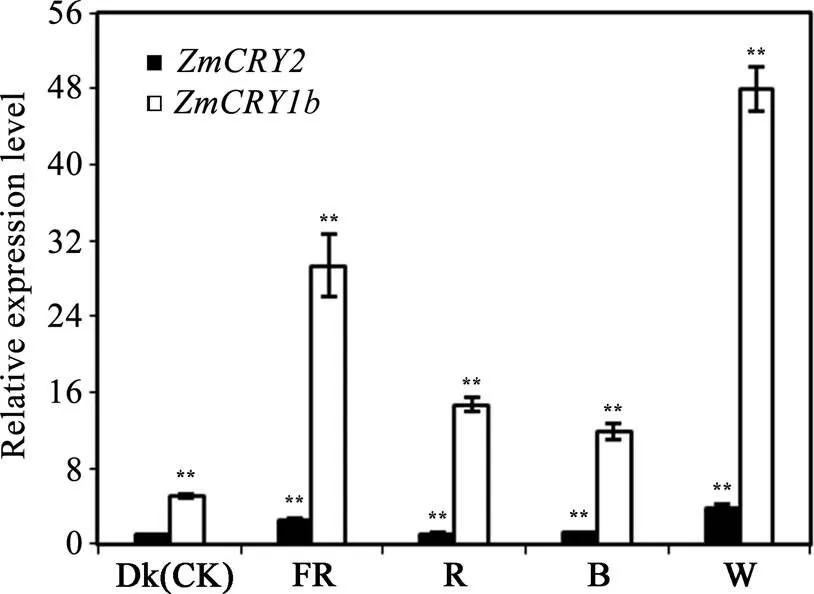
图5 ZmCRY1b和ZmCRY2基因对响应不同光质的相对表达水平分析
米自交系“B73”的幼苗在黑暗、持续远红光(FR, 0.25 μmol m–2s–1)、持续红光(R, 22.3 μmol m–2s–1)、持续蓝光(B, 13 μmol m–2s–1)或持续白光(W, 17 μmol m–2s–1)下生长了13 d。以黑暗条件下的的转录丰度为对照, 并将该的比值设为1。柱状图代表了在3次独立的生物学重复下的平均值, 误差线代表了标准差。用星号表示差异的显著性, *< 0.05, **< 0.01。
The seedlings of maize inbred line B73 were grown in continuous far-red light (FR, 0.25 µmol m−2s−1), red light (R, 22.3 µmol m−2s−1), blue light (B, 13 µmol m−2s−1), or white light (W, 170 µmol m−2s−1) for 13 d. The transcription abundance ofin the dark was set as the control, and the ratio ofin the dark was set as 1. Each column shows the mean expression ofof three biological repeats. Error bars indicate the standard deviation. *< 0.05, **< 0.01.
2.4 ZmCRY1b和ZmCRY2对黑暗到不同光质转换的响应
将黑暗条件下生长13 d的 B73幼苗依次转入到红光、蓝光和白光下0.25 h、0.5 h、1 h、2 h、4 h、8 h、12 h和24 h以及远红光下0.16 h、0.25 h、0.5 h、1 h、1.5 h、2 h、3 h、4 h、6 h、8 h、10 h、12 h、24 h和24.08 h后取材, 用于进一步研究和的表达丰度响应不同光质的差异。将基因在黑暗条件下的表达量设为1, 并以此为对照。
在由黑暗转换到蓝光0.5 h内,的表达丰度持续上升并达到最大值(为黑暗时的42.9倍)(图6-A); 随后在2 h时下降到自身黑暗条件下的水平并保持平缓; 最终在24 h时其转录丰度又回升至黑暗时的5.4倍。从黑暗转换至蓝光时,基因的表达模式与比较类似, 只是整体表达丰度为的12%~55%, 且在由黑暗转到蓝光12~24 h内,的表达丰度略有下降, 而平缓上升。在由黑暗转换到蓝光0.5 h内,的表达丰度达到峰值(为自身黑暗时的13倍); 最后在24 h时其丰度又回升到自身黑暗时的2.15倍。结果表明在由黑暗到蓝光光质转换的响应中比更强烈。
在由黑暗转换到白光的0.25 h时表达丰度出现最大峰值(为黑暗时的24.7倍)(图6-B); 之后在1 h时下降至黑暗时的0.29倍并保持平缓; 在4 h时达到第2个峰值(为黑暗时的3.4倍); 最终在24 h时其转录丰度上升至黑暗时的5.3倍。由黑暗转换至白光时,基因的表达模式与比较类似, 但整体表达丰度为的20%~55%。在0.25 h时同样出现最大峰值, 为自身黑暗时的5.9倍; 在1 h 时下降至自身黑暗时的0.18倍; 在4 h时达到第2个峰值(为自身黑暗时的0.96倍); 在24 h时上升为自身黑暗时的0.9倍(图6-B)。说明在由黑暗到白光光质转换的响应中比更强烈。
尽管编码的是蓝光受体, 但和基因在持续的远红光和红光光质下也有着相对较高的转录丰度(图5), 于是进一步检测了其对红光和远红光的响应(图6-C和D)。结果发现, 在由黑暗转换至红光时,基因的表达丰度维持在较低的水平, 并于1 h和4 h时达到峰值, 分别为黑暗时的4.8倍和14.2倍(图6-C); 在8 h时下降至2.2倍, 此后开始上升, 并于24 h时达到黑暗时的58.0倍。在黑暗转换到红光时表达模式与较类似, 但整体表达丰度为的12%~55%。在1 h和4 h时达到峰值, 分别为自身黑暗时的1.4倍和2.2倍; 在8 h时下降至自身黑暗时的0.6倍, 并于24 h时达到自身黑暗时的7.4倍。结果说明在由黑暗到红光光质转换的响应中也比更强烈。
在由黑暗转到远红光时,的丰度整体低于, 为其表达丰度的15%~55% (图6-D)。在由黑暗转到远红光0.5 h时,的表达丰度下降至黑暗时的1.8倍; 此后分别于1.5 h、4 h、8 h和24 h时达到峰值(分别为黑暗时的4.3、8.4、3.0和16.4倍)。的表达模式与整体类似但略有不同: 在0.25 h时下降至自身黑暗时的0.67倍, 此后开始上升并于1 h时达到峰值(自身黑暗时的1.7倍); 此后分别于4 h、8 h和24 h时再次达到峰值(为自身黑暗时的1.3、0.9和1.0倍)。不同之处在于在0.25~0.50 h内表达丰度上升, 在1.0~1.5 h内下降, 在2~3 h内下降, 这三处与正好相反。结果表明在黑暗到远红光转换光质中比更强烈。

图6 ZmCRY1b和ZmCRY2基因由黑暗到不同光质下的相对表达丰度
玉米自交系B73的幼苗在黑暗生长13 d后, 分别转入蓝光(B, 13 μmol m–2s–1)、白光(W, 17 μmol m–2s–1)、红光(R, 22.3 μmol m–2s–1)下0 h、0.25 h、0.5 h、1 h、2 h、4 h、8 h、12 h和24 h以及远红光(FR, 0.25 μmol m–2s–1)下0 h、0.16 h、0.25 h、0.5 h、1 h、2 h、4 h、8 h、12 h、24 h和24.08 h取材, 用于qRT-PCR分析。把黑暗条件下的比值设为1, 并以此为对照。折线图代表了在3次独立的生物学重复下的平均值, 其中误差线代表了标准差。
The seedlings of maize inbred line B73 were grown in the dark for 13 d, then transferred to blue light (B, 13 µmol m−2s−1), white light (W, 17 µmol m−2s−1), or red light (R, 22.3 µmol m−2s−1) for 0.25 h, 0.5 h, 1 h, 2 h, 4 h, 8 h, 12 h, or 24 h and far-red light (FR, 0.25 µmol m−2s−1) for 0.16 h, 0.25 h, 0.5 h, 1 h, 2 h, 4 h, 8 h, 12 h, 24 h, or 24.08 h. The transcription abundance ofin the dark was set as the control, and the ratio ofin the dark was set as 1. Each line graph shows the mean expression ofof three biological repeats. Error bars indicate the standard deviation.
综合和基因在不同的持续光质以及由黑暗转换到不同光质(白光、蓝光、红光和远红光)下的表达丰度可以发现, 二者均能有效地响应各种不同光质的处理。
2.5 ZmCRY1b和ZmCRY2基因在光周期(长日照和短日照)处理下的表达水平
为了了解玉米对光周期处理的响应能力, 我们采用qRT-PCR分析了和响应长日照和短日照处理的表达模式。将在黑暗阶段结束时的表达量设为1, 并以此为对照。
在长日照条件下,和的表达模式相似, 但整体上的转录丰度低于, 为的3%~10%。在进入光照阶段2 h、6 h和14 h时出现了3个峰值(分别为黑暗阶段结束时的78、88和31倍); 在进入黑暗阶段后, 表达丰度迅速上升, 且在18 h时达到峰值(为黑暗阶段结束时的46倍)。则在进入光照阶段4 h和12 h时出现了峰值(分别为自身黑暗阶段结束时的5.4倍和4.0倍); 在进入黑暗阶段后, 表达丰度迅速上升, 且在18 h时达到峰值(为自身黑暗阶段结束时的4.5倍)(图7-A)。说明和在由白光转到黑暗时表达丰度均能迅速上升至较高的水平, 但能更强烈地响应长日照处理。
在短日照的条件下,和的表达出现了极其相似的模式, 二者的转录水平保持稳定的状态, 但整体上的转录丰度低于, 为其4%~10%。和在光照阶段的峰值均出现在4 h时(此时二者的转录丰度分别为黑暗阶段结束时的30.2倍和2.7倍)。当转入黑暗时, 二者的转录丰度迅速上升。和分别在转入黑暗阶段12 h和16 h时达到峰值(的转录丰度分别为在黑暗阶段结束时的71.5倍和94.1倍,则分别为4.4倍和4.3倍)(图7-B)。可见和的表达能响应长日和短日处理, 且的响应能力更强。
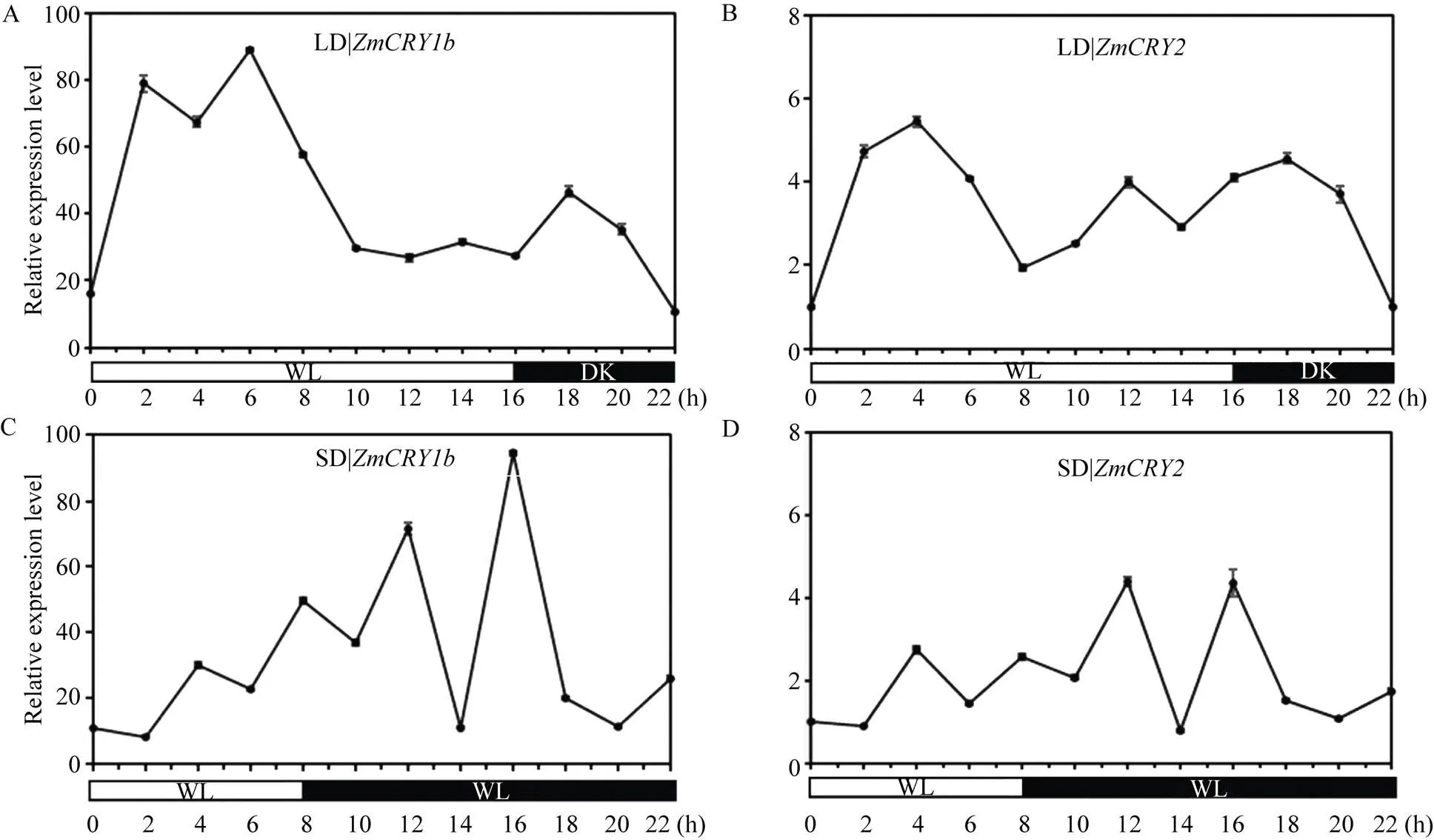
图7 ZmCRY1b和ZmCRY2基因在光周期(长日照和短日照)处理的表达水平
“B73”的幼苗分别在长日照条件下(LD, 16 h光照/8 h黑暗)或短日照条件下(SD, 8 h光照/16 h黑暗)生长13 d后, 每2 h取一次样。把在黑暗结束时期的比值设为1, 并以此为对照。折线图代表了在3次独立的生物学重复下的平均值, 其中误差线代表了标准差。
The seedlings of maize inbred line B73 were grown in long-day condition (LD, 16 h light/8 h dark) or short-day (SD, 8 h light/16 h dark) for 13 d, then sampled every two hours. The transcription abundance ofin the dark was set as the control, and the ratio ofat the end of the dark period was set as 1. Each line graph shows the mean expression ofof three biological repeats. Error bars indicate the standard deviation.
3 讨论
目前对玉米中隐花色素的研究较少且浅显, 而模式植物拟南芥中的隐花色素研究得较为深入。本研究表明, 玉米与拟南芥及小麦和水稻等禾本科植物的CRY蛋白具有相似的结构域和较高的序列一致性。这暗示着它们可能存在相似的功能, 也为如何深入研究玉米的隐花色素提供了技术思路。作为蓝光受体,和能够强烈地响应各种持续光质、转换光质的处理, 暗示着它们可能参与了白光、蓝光甚至是红光和远红光下的光信号转导途径。此外, 二者还能响应长日照和短日照的处理, 说明其可能参与光周期诱导的开花。
拟南芥的CRY1和CRY2在蓝光下可抑制下胚轴的伸长并促进花蕾的形成, 二者分工不同却又功能冗余[29]。在蓝光下, CRY1通过与SPA1结合来抑制COP1/SPA1复合物的活性, 导致HY5的积累以促进光形态建成并抑制下胚轴的伸长, 而CRY2通过结合SPA1以抑制COP1/SPA1复合物的活性, 导致CO蛋白的积累, 或与CIB蛋白结合以促进CO蛋白的积累以及基因的转录, 从而加速开花[21,23]。CRY不仅促进拟南芥的开花, 还能调控植物品种的纬度分布[9]。近期的研究表明, 除拟南芥[9-10]外, 水稻[13-14]、大豆[18-19]、甘蓝型油菜[20]等植物中的CRY均能促进开花。水稻中的OsCRY1介导了蓝光下水稻幼苗的去黄化反应, 而OsCRY2则促进水稻的开花[13-14]。大豆的GmCRY1a和GmCRY2a蛋白与拟南芥的AtCRY1和AtCRY2蛋白序列高度一致, 且均为核蛋白, 但GmCRY1a与AtCRY2的功能类似, 均能促进植物开花[18-19]。本研究显示玉米隐花色素和基因能强烈地响应各种光质及光周期处理, 推测二者也参与了玉米光形态建成和开花期的调控, 但二者对玉米花期的调控作用及其机制尚待验证。
研究表明, 植物中的光敏色素和隐花色素相互作用调控其光形态建成及光周期反应, 进而调控植物的生长和发育过程[10,30-32]。玉米中隐花色素至少存在4个拷贝, 即、、和, 光敏色素则存在6个拷贝, 即、、、、和。它们之间是否存在互作、互作方式及形成的信号网络都值得进一步研究。玉米和基因对玉米开花是否存在调控作用及其与开花的调控基因(、和等)之间的关系也未被阐明。因此, 玉米和基因对玉米光形态建成和光周期的调控作用及机制, 及对玉米的引种和高产的作用也需要进一步研究。
4 结论
玉米CRY1b蛋白与拟南芥、水稻、小麦等CRY1蛋白的结构相似, 均含有PHR和CCE结构域, 而它们的CRY2蛋白则都含有PHR结构域, 且它们的氨基酸序列一致性较高。和基因在叶片中表达丰度最高, 能强烈响应蓝光、白光、红光和远红光持续光质及由黑暗到白光、蓝光、红光、远红光等各种转换光质的处理, 并且对长日照和短日照处理也能很强烈地响应。结果暗示和参与玉米光形态建成及开花调节, 且在作物改良上比更有效, 其在作物改良中的应用价值值得更深入的研究。
[1] 詹克慧, 李志勇, 侯佩, 习雨琳, 肖阳, 孟凡华, 杨建平. 利用修饰光敏色素信号途径进行品种改良的可行性. 中国农业科学, 2012, 45: 3249–3255 Zhan K H, Li Z Y, Hou P, Xi Y L, Xiao Y, Meng F H, Yang J P. A new strategy for crop improvement through modification of phytochrome signaling pathways., 2012, 45: 3249–3255 (in Chinese with English abstract)
[2] Weller J L, Perrotta G, Schreuder M E, van Tuinen A, Koornneef M, Giuliano G, Kendrick R E. Genetic dissection of blue-light sensing in tomato using mutants deficient in cryptochrome 1 and phytochromes A, B1 and B2., 2001, 25: 427–440
[3] Giliberto L, Perrotta G, Pallara P, Weller J L, Fraser P D, Bramley P M, Fiore A, Tavazza M, Giuliano G. Manipulation of the blue light photoreceptor cryptochrome 2 in tomato affects vegetative development, flowering time and fruit antioxidant content., 2005, 137: 199–208
[4] Platten J D, Foo E, Elliott R C, Hecht V, Reid J B, Weller J L. Cryptochrome 1 contributes to blue-light sensing in pea., 2005, 139: 1472–1482
[5] Sharma P, Chatterjee M, Burman N, Khurana J P. Cryptochrome 1 regulates growth and development in Brassica through alteration in the expression of genes involved in light, phytohormone and stress signalling., 2014, 37: 961–977
[6] Yang Z H, Liu B B, Su J, Liao J K, Lin C T, Oka Y. Cryptochromes orchestrate transcription regulation of diverse blue light responses in plants., 2017, 93: 112–127
[7] Sadanandom A, Ádám É, Orosa B, Viczián A, Klose C, Zhang C, Josse E, Kozma-Bognár L, Nagy F. SUMOylation of phytochrome-B negatively regulates light-induced signaling in, 2015, 112: 11108–11113
[8] Liu B, Yang Z H, Adam Gomez, Liu B, Lin C T, Oka Y. Signaling mechanisms of plant cryptochromes in, 2016, 129: 137–148
[9] Yuan S, Zhang Z W, Zheng C, Zhao Z Y, Wang Y, Feng L Y, Niu G Q, Wang C Q, Wang J H, Feng H, Xu F, Bao F, Hua Y, Cao Y, Ma L G, Wang H Y, Kong D D, Xiao W, Lin H H, He Y K.cryptochrome 1 functions in nitrogen regulation of flowering., 2016, 113: 7661–7666
[10] Xu F, He S B, Zhang J Y, Mao Z L, Wang W X, Li T, Hua J, Du S S, Xu P B, Li L, Lian H L, Yang H Q. Photoactivated CRY1 and phyB interact directly with AUX/IAA proteins to inhibit auxin signaling in., 2017, 11: 523–541
[11] Facella P, Daddiego L, Perrotta G. CRY1a influences the diurnal transcription of photoreceptor genes in tomato plants after gibberellin treatment., 2012, 7: 1034–1036
[12] Li Y Y, Mao K, Zhao C, Zhang R F, Zhao X Y, Zhang H L, Shu H R, Zhao Y. Molecular cloning of cryptochrome 1 from apple and its functional characterization in., 2013, 67: 169–177
[13] Zhang Y C, Gong S F, Sang F, Yang H Q. Functional and signaling mechanism analysis of rice CRYPTOCHROME 1. 2006,, 46: 971–983
[14] Hirose F, Shinomura T, Tanabata T, Shimada H, Takano M. Involvement of rice cryptochromes in de-etiolation responses and flowering., 2006, 47: 915–925
[15] Platten J D, Foo E, Foucher F, Hecht V, Reid J B, Weller J L. The cryptochrome gene family in pea includes two differentially expressedgenes., 2005, 59: 683–696
[16] Imaizumi T, Kanegae T, Wada M. Cryptochrome nucleocytoplasmic distribution and gene expression are regulated by light quality in the fern-veneris., 2000, 12: 81–96
[17] Imaizumi T, Kadota A, Hasebe M, Wada M. Cryptochrome light signals control development to suppress auxin sensitivity in the moss physcomitrella patens., 2002, 14: 373–386
[18] Meng Y Y, Li H Y. Blue light-dependent interaction between cryptochrome 2 and CIB1 regulates transcription and leaf senescence in soybean., 2013, 25: 4405–4420
[19] Zhang Q Z, Li H Y, Li R, Hu R B, Fan C M, Chen F L, Wang Z H, Liu X, Fu Y F, Lin C T. Association of the circadian rhythmic expression ofwith a latitudinal cline in photoperiodic flowering of soybean.2008, 105: 21028–21033
[20] Chatterjee M, Sharma P, Khurana J P. Cryptochrome 1 fromis up-regulated by blue light and controls hypocotyl/stem growth and anthocyanin accumulation., 2006, 141: 61–74
[21] Liu H T, Liu B, Zhao C X, Pepper M, Lin C T. The action mechanisms of plant cryptochromes., 2011, 16: 684–691
[22] Ahmad M, Cashmore A R.gene ofencodes a protein with characteristics of a blue-light photoreceptor., 1993, 366: 162–166
[23] Guo H W, Yang H Y, Mockler T C, Lin C T. Regulation of flowering time byphotoreceptors., 1998, 279: 1360–1363
[24] Wu G, Spalding E P. Separate functions for nuclear and cytoplasmic cryptochrome 1 during photomorphogenesis ofseedlings., 2007, 104: 18813– 18818
[25] Yu X H, Klejno J, Zhao X Y, Dror S, Maskit M, Yang H Y, Janet L, Liu X M, Javier L, Lin C T.cryptochrome 2 completes its posttranslational life cycle in the nucleus., 2007, 19: 3146–3156
[26] Kleine T, Lockhart P, Batschauer A. Anprotein closely related tois targeted to organelles., 2003, 35: 93–103
[27] 闫蕾, 杨宗举, 苏亮, 肖阳, 郭林, 宋梅芳, 孙蕾, 孟凡华, 白建荣, 杨建平. 2个玉米基因的克隆及其响应光质处理的表达模式. 作物学报, 2016, 42: 1298–1308 Yan L, Yang Z J, Su L, Xiao Y, Guo L, Song M F, Sun L, Meng F H, Bai J R, Yang J P. Molecular cloning of two maize ()genes and their expression patterns of in response to different light treatments., 2016, 42: 1298–1308 (in Chinese with English abstract)
[28] Rajeevan M S, Ranamukhaarachi D G, Vernon S D, Unger E R. Use of real-time quantitative PCR to validate the results of cDNA array and differential display PCR technologies., 2001, 25: 443–451
[29] Yang Y J, Zuo Z C, Zhao X Y, Li X, John K, Lia Y, Chen P, Liang S P, Yu X H, Liu X M, Lin C T. Blue-light-independent activity ofcryptochromes in the regulation of steady-state levels of protein and mRNA expression., 2008, 1: 167–177
[30] Wang Q, Liu Q, Wang X, Zuo Z, Oka Y, Lin C. New insights into the mechanisms of phytochrome-cryptochrome coaction., 2018, 217: 547–551
[31] de Wit M, Keuskamp D H, Bongers F J, Hornitschek P, Gommers C M M, Reinen E, Martínez-Cerón C, Fankhauser C, Pierik R. Integration of phytochrome and cryptochrome signals determines plant growth during competition for light., 2016, 26: 3320–3326
[32] Xu P B, Lian H L, Wang W X, Xu F, Yang H Q. Pivotal roles of the phytochrome-interacting factors in cryptochrome signaling., 2016, 9: 496–497
Transcription Abundances ofandGenes in Response to Different Light Treatments in Maize
LI Hong-Dan1,2, YAN Lei1,2, SUN Lei1,2, FAN Xiao-Cong1,3, CHEN Shi-Zhan1,3, ZHANG Yan1,3, GUO Lin1, YOU Guang-Xia1, LI Zhuang1,2, YANG Zong-Ju1,2, SU Liang1,*, and YANGJian-Ping1,3, *
1Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China;2Graduate School, Chinese Academy of Agricultural Sciences, Beijing 100081, China;3College of Agronomy, Henan Agricultural University, Zhengzhou 450002, Henan, China
Light is lightly related to the important agronomic traits in maize such as plant height, flowering time, yield and quality. Cryptochromes are blue and ultraviolet-A photoreceptors generally existing in animal, plant and microbial, which mainly regulate photomorphogenesis in plants and circadian rhythms in both of plant and animal. Therefore, the expression pattern analysis of cryptochrome in maize could lay a research foundation in the photomorphogenesis in maize. Theandgenes were cloned by RT-PCR. Their proteins’ function domains and the phylogenetic analysis of amino acid sequences were carried out through bioinformatics analysis. The transcription abundances ofandgenes in different tissues of inbred line B73 under different light treatments were analyzed by qRT-PCR. We found that the function domains of ZmCRY1b or ZmCRY2 protein was consistent with CRY1 or CRY2 in, rice and wheat, which contains the PHR and CCE domains or the PHR domain, respectively. Phylogenetic analysis indicated that the three gramineous CRYs from maize, wheat, and rice belonged to the same branch, while showing low similarity to other CRY1 proteins from dicotyledons.andgenes highly expressed in leaf. Meanwhile, they could respond to all treatments of different continuous light conditions, the transitions from the dark to light conditions, as well as long-day and short-day conditions. The transcription abundances ofin all treatments were higher than those of, indicating thatwas more important thanin maize. In conclusion, both ofandgenes can greatly respond to different light conditions and light cycle treatments, and play an important role in maize photomorphogenesis. Our results also provide a research basis for functional exploration ofandin crop improvement.
maize; cryptochrome; photomorphogenesis; light treatment; transcription abundance
2018-01-25;
2018-06-12;
2018-07-02.
10.3724/SP.J.1006.2018.01290
杨建平, E-mail: yangjianping02@caas.cn, Tel: 010-82105859; 苏亮, E-mail: suliang_mp5@163.com, Tel: 010-82105851
E-mail: danielle_li2014@163.com, Tel: 010-82105851
本研究由国家转基因生物新品种培育科技重大专项(2016ZX08010002-003-002),北京市自然科学基金(重点)项目(6151002)和国家自然科学基金项目(31570268)资助。
This study was supported by the Major Project of China on New Varieties of GMO Cultivation (2016ZX08010002-003-002), the Key Project of Beijing Natural Science Foundation (6151002), and the National Natural Science Foundation of China (31570268).
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20180629.1608.010.html
