基于人工神经网络的L-天冬酰胺酶发酵培养基优化
2018-09-06王云龙刘松堵国成陈坚
王云龙,刘松*,堵国成,陈坚
1(江南大学 生物工程学院,江苏 无锡,214122) 2(江南大学,工业生物技术教育部重点实验室,江苏 无锡,214122)
L-天冬酰胺酶(EC 3.5.1.1,L-Asparaginase,L-ASNase)是一种酰胺基水解酶,可以将天冬酰胺脱氨基生成天冬氨酸和氨[1]。该酶具有抗肿瘤活性,已被用于治疗淋巴系统恶性肿瘤、儿童急性淋巴细胞白血病、网状细胞肉瘤及霍金森病等疾病[2]。最新报道显示,该酶也可用于油炸食品中以减少丙烯酰胺的生成[3-4]。由于L-ASNase在食品与医药领域中的重要应用,已引起国内外广大学者的极大兴趣。
L-ASNase广泛存在于动植物及微生物中[5]。但是由于动植物中L-ASNase含量少,分离提取困难,而微生物发酵法具有培养简单、提取纯化简便以及易于大规模生产等优点,已成为商品化L-ASNase的主要来源[6]。由于野生菌发酵L-ASNase产量往往较低[7-8],应用重组菌发酵成为提高L-ASNase产量的重要途径[9-11]。KHUSHOO[12]等通过控制重组菌E.coliBLR(DE3)比生长速率并优化诱导剂添加时间,L-ASNase酶活在2 L发酵罐水平达到870 U/mL,其摇瓶水平L-ASNase产量为30.67 U/mL。龙水清[13]通过补料和溶氧控制策略将B.subtilis168L-ASNase的最终产量在5 L发酵罐水平提高到112.61 U/mL。SUSHMA[14]等通过培养基优化和持续诱导策略使B.subtilisWB800NL-ASNase产量在3 L发酵罐水平达到525.98 U/mL,其摇瓶水平为381.4 U/mL。
培养基是重组菌株生长和产物合成的基础,优化培养基组成对重组L-ASNase生产具有重要意义。目前,一系列试验设计方法和算法应用于培养基优化。KRISHNAN等[15]通过Plackett-Burman实验优化乳酸菌产乳酸培养基,转化率提高了30%。LI等[16]通过响应面优化培养基使吩嗪-1-羧酸产量提高3倍。WU等[17]通过人工神经网络-遗传算法模型使核黄素的产量提高了76.4%。其中,人工神经网络是模仿人脑处理情报方式的100%黑箱性质的模型,对于高度非线性且含有噪声的生物系统非常适用[18]。
本研究以研究室前期构建的菌株Bacillussubtilis/ASNΔ25/B2为生产菌株[11],通过单因素实验、Plackett-Burman实验、最陡爬坡实验、中心组合实验和神经网络模型对B.subtilis/ASNΔ25/B2产L-ASNase发酵培养基进行优化,以充分发挥菌株生产性能,提高L-ASNase产量。
1 材料与方法
1.1 材料与试剂
菌株:B.subtilis/ASNΔ25/B2[11],由江南大学生物系统与生物加工工程研究室构建并保藏。
试剂:酵母粉,胰蛋白胨,英国Oxoid公司,BR;安琪酵母蛋白胨FP103,安琪胰蛋白胨FP318,安琪酵母股份有限公司,BR,安琪胰蛋白胨FP319,安琪酵母股份有限公司,BR;思宾格酵母蛋白胨,思宾格0805酵母浸粉,思宾格0806酵母浸粉,广州一品鲜生物科技有限公司;L-天冬酰胺,美国Sigma-Aldrich公司;其他常用试剂均为国药集团化学试剂有限公司分析纯。
卡那平板培养基(g/L):胰蛋白胨10,酵母粉5,NaCl 10,琼脂20,硫酸卡那霉素0.05。
种子培养基(g/L):胰蛋白胨10,酵母粉5,NaCl 10,硫酸卡那霉素0.05。
初始发酵培养基(g/L):蔗糖35,胰蛋白胨20,尿素0.8,玉米浆8,K2HPO4·3H2O 3.26,KH2PO42.5,MgSO4·7H2O 1.8,NaCl 3,L-天冬酰胺1.2。初始pH 7.5。
优化发酵培养基(g/L):蔗糖65,酵母蛋白胨28,玉米浆11,KH2PO411.5,NaCl 3.3,(NH4)2SO44,K2HPO4·3H2O 22.5,MgSO4·7H2O 1,L-天冬酰胺2。初始pH 7.5。
1.2 仪器与设备
XWY-240恒温摇床,上海智诚仪器有限公司;台式高速离心机,德国eppendorf公司;UV-2450型紫外-可见分光光度计,日本Shimadzu公司;pH计,瑞士Mettler公司。
1.3 方法
1.3.1 种子活化
取适量保存在冷冻甘油管中的菌液涂布至卡那平板上,37 ℃培养过夜。
1.3.2 种子培养
将活化好的种子接种至装有60 mL种子培养基的500 mL三角瓶中,添加60 μL的50 mg/mL硫酸卡那霉素,摇床温度37 ℃,转速220 r/min,培养8~10 h。
1.3.3 摇瓶发酵培养
按4%的接种量将种子培养基接入装有25 mL发酵培养基的250 mL摇瓶中,温度37 ℃,转速220 r/min。
1.4 实验设计
1.4.1 Plackett-Burman 实验设计
在单因素实验的基础上使用Design-Expert.V 8.0.6进行N=12的Plackett-Burman实验设计和结果分析[19],对发酵培养基中的9种成分进行研究。每个因素取高(+1)、低(-1)两个水平,2个虚拟变量用来估计实验误差,以酶活为响应值。各组分及其水平设置如表1所示。

表1 Plackett-Burman实验设计中变量的真实值Table 1 Real values of variables in Plackett-Burman experimental design
1.4.2 最陡爬坡实验
根据Plackett-Burman实验结果,综合考虑各因素效应正负和原料成本来确定最陡爬坡实验中各因素水平。
1.4.3 中心组合实验
以最陡爬坡实验的最优条件为水平中心,使用Design-Expert.V8.0.6进行5因素5水平的中心组合实验(简称CCD实验)[19],如表2所示。

表2 CCD实验中的变量和水平Table 2 Variable and its level in CCD design
1.4.4 神经网络模型
使用JMP10.0以CCD实验结果作为训练和验证数据样本,选择“K重”交叉验证的方法执行模型启动。设置合适的折数和隐藏节点数,过拟合罚项0.001,历程数20,最大迭代数50,收敛准则0.000 01。
1.5 分析方法
1.5.1 菌体浓度的测定
采用浊度法测定。取适量发酵液稀释到适当倍数后使用紫外分光光度计于波长600 nm下测定吸光度,即OD600值。
1.5.2L-ASNase粗酶液制备
取一定量的发酵液于12 000 r/min离心10 min,上清液即为L-ASNase粗酶液。
1.5.3L-ASNase酶活测定
采用奈氏试剂法[11]。取900 μL磷酸盐缓冲液(20 mmol/L K2HPO4-KH2PO4缓冲液,pH 7.5)加入200 μL底物(L-天冬酰胺,0.15 mol/L)于37 ℃保温10~20 min后,加入100 μL稀释到适当倍数的L-ASNase粗酶液,37 ℃反应10 min后加入100 μL终止剂(三氯乙酸,1.5 mol/L)终止反应,对照组在保温前即加入终止剂。反应结束后于12 000 r/min离心2 min,取100 μL上清,加入3 400 μL去离子水和500 μL奈氏试剂,混匀后在波长436 nm下测定吸光度。
酶活力单位定义:37 ℃每分钟水解L-天冬酰胺生成1 μmol氨所需要的酶量定义为1个L-ASNase活力单位。
2 结果与分析
2.1 单因素实验
2.1.1 蔗糖质量浓度对L-ASNase酶活的影响
贾明媚[20]、陈璇[21]、SUSHMA[14]研究结果表明,蔗糖是B.subtilis合成L-ASNase的最适碳源。碳源浓度对菌体生长和产酶有较大的影响[21]。对初始培养基中的蔗糖质量浓度进行优化,结果如图1所示。随着蔗糖质量浓度的增加,菌体量和L-ASNase产量持续提高。在蔗糖50 g/L时,L-ASNase产量达到最大值,继续增加蔗糖质量浓度虽然有利于菌体生长,但不利于产酶。

图1 蔗糖质量浓度对L-ASNase酶活的影响Fig.1 Effect of sucrose concentration on L-ASNase activity
2.1.2 氮源对L-ASNase酶活的影响
在优化了蔗糖质量浓度的基础上,添加相同质量的不同有机氮源替换初始培养基中的胰蛋白胨,结果如图2所示。思宾格酵母蛋白胨和胰蛋白胨效果较好且接近,而酵母蛋白胨价格要远远低于胰蛋白胨,故选择酵母蛋白胨做进一步研究。酵母蛋白胨质量浓度对L-ASNase酶活的影响如图2-B所示,随着酵母蛋白胨浓度的增加,菌体量和酶活一直在增加。

图2 有机氮源对L-ASNase酶活的影响Fig.2 Effect of organic nitrogen on L-ASNase activity


图3 无机氮源对L-ASNase酶活的影响Fig.3 Effects of inorganic nitrogen sources on L-ASNase activity
2.2 Plackett-Burman实验
Plackett-Burman实验是一种从多个因素中筛选出对实验结果有显著影响的因素的实验方法[19],可以用最少的试验次数使因素的主效果得到尽可能精确估计。在单因素实验确定的因素水平的基础上,按照表1对发酵培养基中9种组分进行考察,以筛选出对酶活影响显著的因素作进一步优化。Plackett-Burman实验结果及方差分析如表3和表4所示。

表3 Placket-Burman实验设计及对应的L-ASNase酶活Table 3 Placket-Burman experimental design and corresponding L-ASNase activity

表4 Placket-Burman实验中各因素的影响Table 4 Effects of variables in the Placket-Burman experiment
注:R2=0.966 2,校正系数=0.907 0。
由表4中效应或预测系数可知,9种成分中蔗糖、酵母蛋白胨、玉米浆、MgSO4·7H2O对酶活表现为正效应,其他因素表现为负效应。由p值可知,蔗糖、酵母蛋白胨、玉米浆、KH2PO4和NaCl对酶活的影响显著(p值<0.05),其他4种因素对酶活影响不显著(p值>0.05)。对酶活影响不显著的因素(NH4)2SO4、K2HPO4·3H2O和L-天冬酰胺效应均为负,接下来的实验质量浓度均取Plackett-Burman实验的低水平,即质量浓度分别设为:4、22.5、2 g/L,MgSO4·7H2O效应为正,取高水平,即质量浓度设为1 g/L。下一步将对上述5种显著因素的质量浓度水平进行优化。
2.3 最陡爬坡实验
在Plackett-Burman实验基础上,运用最陡爬坡实验对蔗糖、酵母蛋白胨、玉米浆、KH2PO4和NaCl等5种对酶活影响显著的因素进行优化,以确定各因素进行CCD实验的水平中心。综合考虑正负效应和原料成本,确定最陡爬坡实验各因素的水平,如表5所示。由表5可知,第4组实验获得的酶活最大,故以该培养基组成为CCD实验的水平中心。

表5 最陡爬坡实验设计及结果Table 5 Experiment design of path of steepest ascent and the corresponding results
2.4 CCD实验
按照表2采用CCD实验来确定神经网络优化需要输入的数据,结果如表6所示。

表6 CCD实验设计及结果Table 6 CCD experimental design and results
2.5 神经网络模型的建立
“K重”交叉验证的方法可以将建立的神经网络模型很好的推广到新数据,故选择“K重”交叉验证的方法对CCD实验建立的数据样本执行模型启动。建立神经网络模型时,隐藏节点数是需要指定的重要数值。如果将此值设定得过低,则会拟合不足;而如果过高,则会过度拟合[22-23]。在经过大量神经网络训练后,确定折数为5,隐藏节点数为7,采用5×7×1的3层神经网络结构,如图4所示。即5个输入神经元,分别代表蔗糖、酵母蛋白胨、玉米浆、KH2PO4和NaCl;7个隐含层神经元;1个输出神经元,代表酶活。设置折数5,隐藏节点数为7,过拟合罚项0.001,历程数20,最大迭代数50,收敛准则0.000 01,执行神经网络模型的拟合迭代过程。训练拟合决定系数R2为0.978 5(图5-A),验证拟合决定系数R2为0.995 1(图5-B),说明该5×7×1的3层神经网络结构能很好地对数据样本进行拟合。

图4 神经网络结构图Fig.4 Structure of artificial neural network
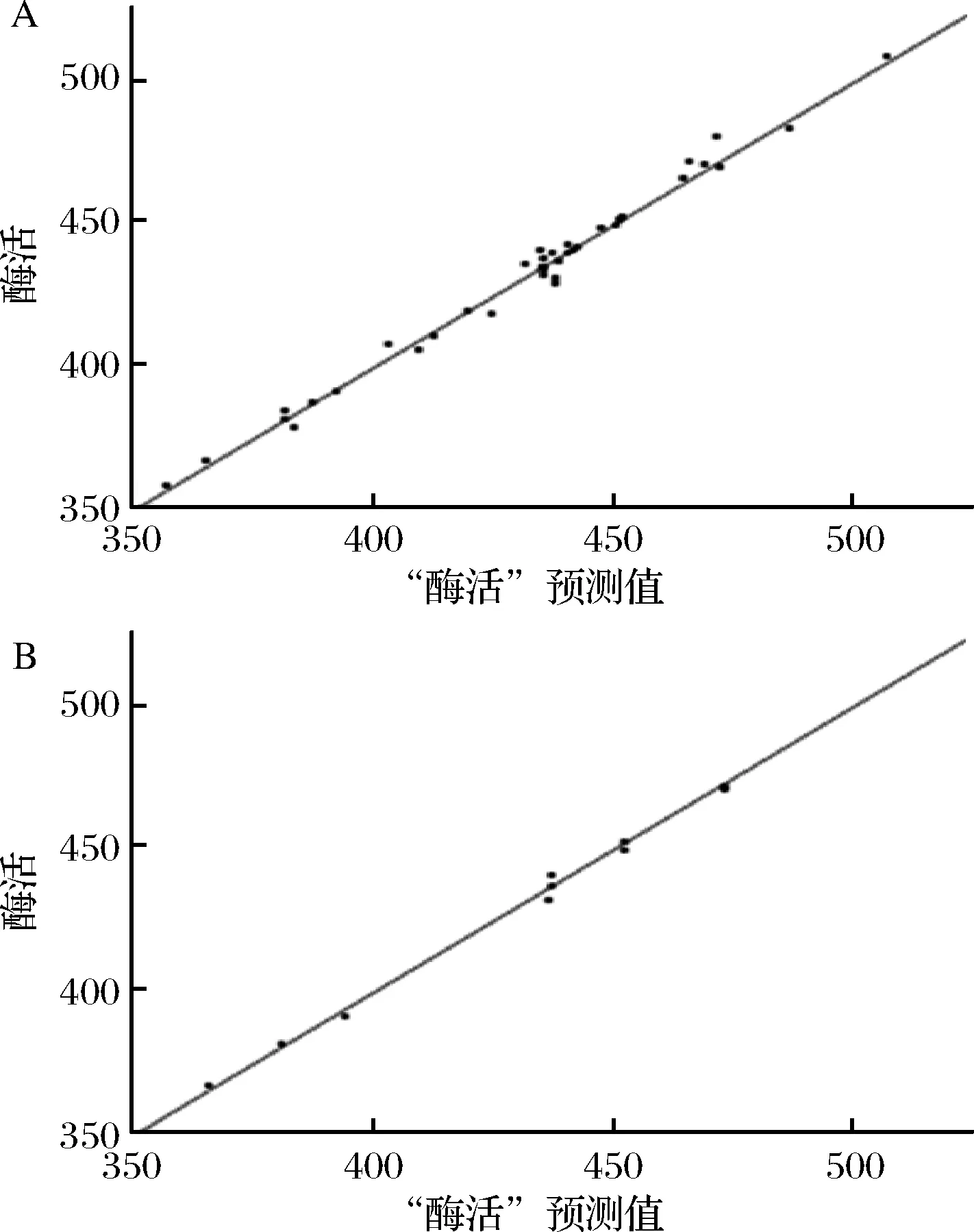
图5 酶活预测值与实际值相关性Fig.5 The correlation of actual values and the predicted values
2.6 神经网络模型优化结果分析
为了考察5种显著成分对L-ASNase酶活的影响,利用JMP10.0中的刻画器功能进行分析,结果如图6所示。

图6 神经网络预测刻画图Fig.6 Prediction plot of the neural network
由图6可知,蔗糖质量浓度对酶活有较大的影响。随着蔗糖质量浓度的增加,酶活逐渐增加,在蔗糖质量浓度为65 g/L时达到最大。继续增加蔗糖质量浓度,酶活反而下降。这可能是因为过高的蔗糖质量浓度会导致有机酸积累,抑制菌体生长[20],从而导致产量下降。随着酵母蛋白胨质量浓度的增加,酶活也逐渐增加,在酵母蛋白胨质量浓度为28 g/L时达到最大。继续增加酵母蛋白胨质量浓度,酶活反而下降。这可能氮源质量浓度过高会导致菌体失水[20]。随玉米浆质量浓度的增加,酶活反而下降。这可能是因为玉米浆质量浓度增加时耗糖速率也随之增加,副产物也会增加[24],不利于产酶。随着KH2PO4质量浓度的增加,酶活先增加后减小。这可能是因为KH2PO4和K2HPO4的加入可以促进菌体生长和外源蛋白的合成,但过高的KH2PO4浓度会导致菌体衰退[25]。随着NaCl质量浓度的增加,L-ASNase酶活先增大后减小,过高的盐浓度会破坏渗透调节,对菌体生长不利[26]。
由预测刻画图可知,当蔗糖为65 g/L、酵母蛋白胨28 g/L、玉米浆11 g/L、KH2PO411.5 g/L、NaCl 3.3 g/L时,L-ASNase酶活最大预测值为517.3 U/mL。为了验证神经网络模型预测的准确性,利用此培养基进行3次独立实验。结果表明,在优化的培养基条件下,L-ASNase酶活平均值为515.6 U/mL,和预测值十分接近,说明建立的神经网络模型能有效地进行预测。
3 结论
本研究使用单因素实验和Plackett-Burman实验筛选出培养基中影响酶活的5个显著因素分别为:蔗糖、酵母蛋白胨、玉米浆、KH2PO4和NaCl。通过CCD实验建立数据样本,使用JMP10.0神经网络平台构建了5×7×1三层神经网络结构。该模型预测值和实际值十分接近,能准确地反映出网络输入元与输出元之间的映射关系。最终得到最优培养基组成为:蔗糖65 g/L、酵母蛋白胨28 g/L、玉米浆11 g/L、KH2PO411.5 g/L、NaCl 3.3 g/L、(NH4)2SO44 g/L、K2HPO4·3H2O 22.5 g/L、MgSO4·7H2O 1 g/L、L-天冬酰胺2 g/L。在最优培养基条件下,L-ASNase酶活可达515.6 U/mL,和优化前相比酶活提高了90.9%。本研究的L-ASNase最终产量虽然低于KHUSHOO[12]和SUSHMA[14]报道的发酵罐水平产量,但比其摇瓶水平高很多,具有较大的潜在能力,为实现重组B.subtilis高效生产L-ASNase提供了基础数据。
