全球抗草甘膦杂草的概况
2018-08-28张翼翾编译
张翼翾 编译
(上海市农药研究所,上海 200032)
草甘膦无选择性,具有内吸性,是杀草谱最广的芽后除草剂。其作用于莽草酸途径,抑制叶绿体酶5-烯醇丙酮酰莽草酸-3-磷酸合酶(EPSP),使芳香酸的合成受阻。草甘膦在1974年首次引入,当时价格相对较高,只用于高价值的园艺作物(果园)和一些非作物场所;随着价格的下降,被用于许多其他情况,包括作物播前和休耕地的杂草防除,可使种植者少耕地或不耕地。
在1996年引入了抗草甘膦大豆,其是开发和应用最早的抗草甘膦作物,这样草甘膦就可用于选择性地芽后除草。在同一年,在每年都使用草甘膦且连续使用 15年的苹果园中发现了首个抗草甘膦杂草——瑞士黑麦草(Lolium rigidum),此例抗性与抗草甘膦作物无关。自此,全球抗草甘膦杂草品种稳定增长(图1),其中大部分是由于在抗草甘膦作物田中草甘膦的使用增加所致。草甘膦防效高,杀草谱广,成本低,很快在抗草甘膦大豆、玉米、棉花、油菜和甜菜田应用。抗草甘膦作物的引入使杂草防治成本降低,对新除草剂的引入具有不利的影响,致使除草剂发现项目(图2)减少。如果不是抗草甘膦杂草的产生,新除草剂开发的减少将不会成为大的问题,然而,抗草甘膦杂草的出现以及在30多年中新作用位点除草剂的缺乏使作物受到抗性杂草的危害越来越严重。
1 抗草甘膦杂草发生的国家
国际抗性杂草调查网站(http://www.weedscience.org)有全球抗除草剂杂草发生情况。此调查数据是杂草科学家根据网站(http://www.weedscience.com/Documents/Resistance Criterion.pdf)上的调查要求进行重复田间试验验证的除草剂抗性所得到的。毫无疑问,调查数据要低于真实数据,因为许多抗性事例还没有得到科学证实,或没有呈交给网站,特别是不发达国家的抗性例。有38个杂草品种对草甘膦产生了抗性,其中单子叶杂草 18种,双子叶杂草20种(图 1)。其中一半为抗草甘膦作物系统中的杂草,另一半为果园、农场、谷物、休闲地和非作物场所。虽然抗草甘膦作物和其他作物系统中的抗性杂草种类数量相近,但抗草甘膦作物系统中的抗草甘膦杂草侵染的面积和造成的经济损失占 90%以上。美国、阿根廷、巴西和加拿大是种植抗草甘膦作物最早的国家,这4个国家是抗草甘膦杂草品种最多的前5位国家(图3),抗草甘膦杂草侵染的面积也最大。抗草甘膦作物最早引入美国,并被广泛种植,在美国抗草甘膦杂草问题也最严重。
1.1 美国
在美国有17种杂草对草甘膦产生了抗性(表1)。其中13种发生于抗草甘膦作物田中,7种在抗草甘膦作物和其他场所中都发现,4种只在其他场所发生(主要果园、路边、高尔夫球场和休闲地)。在抗草甘膦作物体系中发现的第一个抗草甘膦杂草为小飞蓬(Conyza canadensis),在2000年美国的特拉华州的抗草甘膦大豆田中发现。小飞蓬是分布最广的抗草甘膦杂草,在11个国家和美国的25州发现。然而,由于具有有效的、费用不高的可代替防治措施,其造成的经济损失不大。在2004年在美国佐治亚州的棉花田中首次发现抗草甘膦长芒苋(Amaranthus palmeri),目前在美国的 27个州出现。此抗性杂草是全球经济危害最大的抗草甘膦杂草,一般发生在抗草甘膦棉花、玉米和大豆田中。糙果苋(Amaranthus tuberculatus)是另一重要的抗草甘膦杂草,是美国18个州的难防除杂草。抗性长芒苋主要发生在美国南部州,而抗性糙果苋在北部州更常见,是中西部玉米和大豆生产中最大的威胁。2种苋都让人不安,它们已对这些主要作物中使用的其他大部分作用位点的除草剂产生了抗性。国际抗性杂草调查数据库中有许多植物对4个或更多作用位点的除草剂产生多抗的例子。
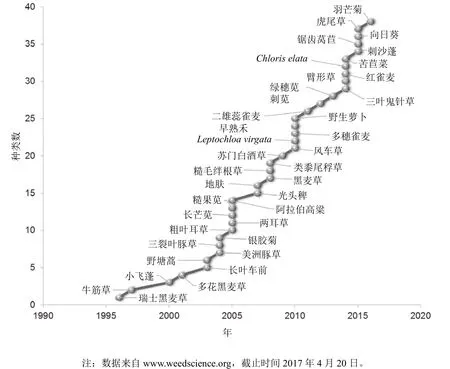
图1 全球抗草甘膦杂草品种数量的增长情况
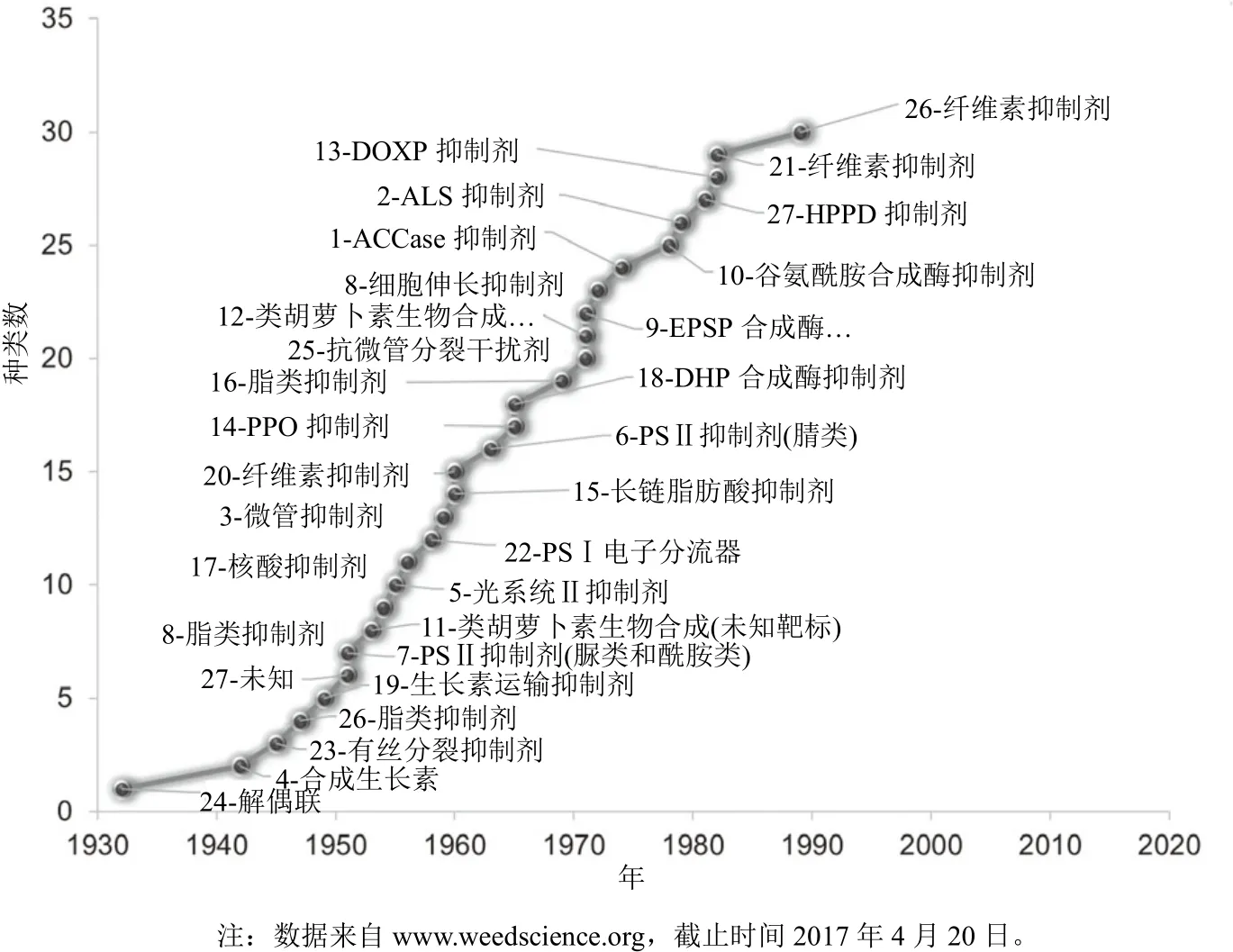
图2 新作用位点除草剂的引入时间
在 2007年在美国堪萨斯州首次确定了抗草甘膦的地肤(Kochia scoparia),现在在美国10个州出现。由于地肤具有“风滚草”分散特性,故其扩展速度特别快。抗草甘膦美洲豚草(Ambrosia artemisiifolia)和三裂叶豚草(Ambrosia trifida)分别在美国的15个和25个州发现,二者分布虽然没有白酒草属(Conyza spp.)、苋属(Amaranthus spp.)和地肤广泛,但一旦发生就会对玉米和大豆生产产生毁灭性破坏。与这小飞蓬、苋属和地肤相比,可能是抗性每周豚草和三裂叶豚草的种子和繁殖力大限制了其扩展速度。
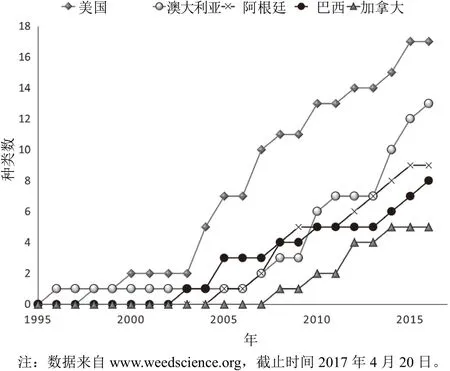
图3 在抗草甘膦杂草品种数量最多的5个国家抗性杂草品种增加情况
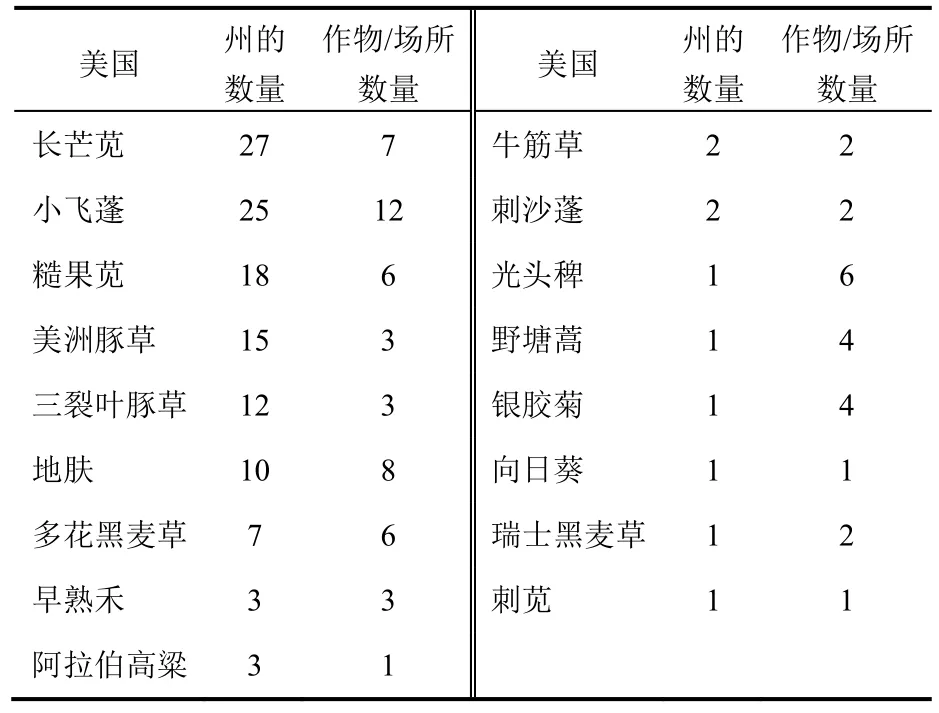
表1 美国抗草甘膦杂草品种发生情况
1.2 澳大利亚
在澳大利亚瑞士黑麦草(Lolium rigidum )是第一个对草甘膦产生抗性的杂草,发生在新威尔士苹果园。和美国不一样,澳大利亚种植抗草甘膦作物的时间不长,草甘膦主要用于非选择性除草。尽管如此,已有13种杂草对草甘膦产生了抗性(表2),仅次于美国。在澳大利亚有这么多抗除草剂杂草的部分原因可能是澳大利亚有相对多的合格的杂草科学家在搜寻抗性杂草。在其他大多数国家,许多抗性事例可能被忽略。例如在澳大利亚在栅栏和路边已确定5种抗草甘膦杂草。毫无疑问,在美国路边许多抗草甘膦杂草未被发现。在澳大利亚抗草甘膦杂草多的另一原因(与美国相比澳大利亚农田相对少)可能是使用的草甘膦剂量低于美国的南部和北部,这样就形成了对比高水平抗性更易发展的低水平抗性机制的选择。
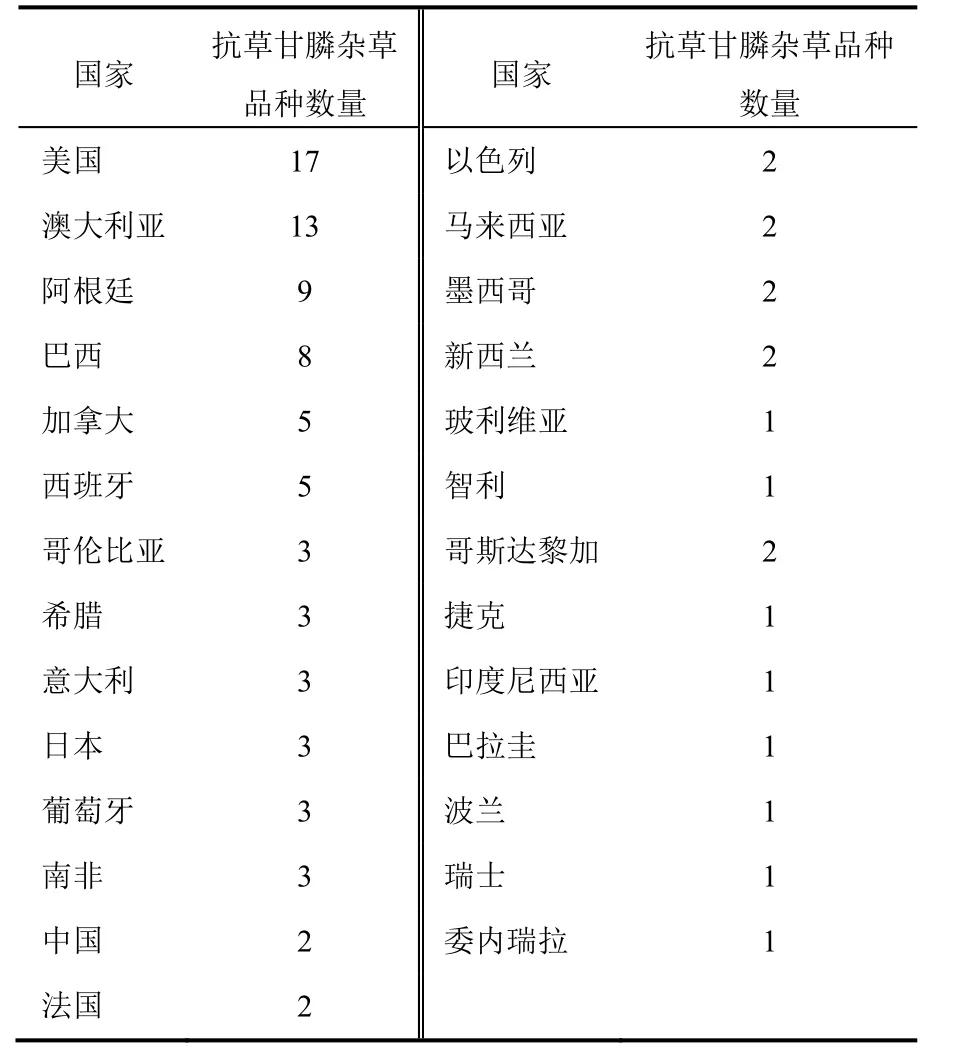
表2 不同国家抗草甘膦杂草品种数量
1.3 阿根廷
在20世纪90年代末阿根廷快速地接纳了抗草甘膦作物,到目前已有9种杂草对草甘膦产生了抗性,主要发生在抗草甘膦大豆田中。抗草甘膦阿拉伯高梁(Sorghum halepense)广泛分布于阿根廷北部大豆种植区。令人惊奇的是阿拉伯高粱是美国南部常见的杂草,报道在美国只有少数抗草甘膦事例。这是否是由于2个国家的农业技术措施的不同导致抗性扩展速度的差异,在阿根廷抗草甘膦阿拉伯高粱是通过污染的大豆种子和/或收割机扩展所致,还需研究。在阿根廷,除了抗草甘膦阿拉伯高粱,其他抗草甘膦禾本科杂草[两耳草(Digitaria insularis)、光头稗(Echinochloa colona)、牛筋草(Eleusine indica)]也是抗草甘膦大豆田中重要的杂草。2个抗草甘膦苋属品种(A. hybridus subsp. quitensis和长芒苋)在抗草甘膦玉米和大豆田越来越普遍,长芒苋被认为是从美国进口的受污染的大豆种子而引入阿根廷大豆田中。
1.4 巴西
和美国和阿根廷一样,在巴西也出现了抗草甘膦杂草,这主要是由于在抗草甘膦大豆田中广泛使用草甘膦所致。在巴西,在1999年就开始种植抗草甘膦大豆(最初是从阿根廷非法进口种子),到目前已确定8个抗草甘膦杂草品种。在抗草甘膦作物田中危害最严重的抗性杂草为白酒草属[小飞蓬(C.canadensis)、野塘蒿(Conyza bonariensis)、苏门白酒草(Conyza sumatrensis)]、两耳草、牛筋草(E. indica)以及最近发生的长芒苋。
1.5 加拿大
和澳大利亚一样,在加拿大少耕系统,很长时间以来在作物种子发芽以前应用草甘膦进行灭生性除草。因此,对草甘膦抗性形成了大的选择压。此外,加拿大已种植抗草甘膦作物,在安大略省主要为抗草甘膦大豆和玉米,在草原三省为抗草甘膦油菜。目前,在加拿大已报道有5种杂草对草甘膦产生了抗性,第1个为在安大略省抗草甘膦作物田中发现的三裂叶豚草(A. trifida)、以及在同一作物系统的小飞蓬、美洲豚草和糙果苋。在草原三省抗草甘膦油菜田中草甘膦抗性发展慢,这可能是种植者经常轮作抗草甘膦、抗咪唑啉酮和抗草铵膦油菜,甚至是抗溴苯腈油菜。在草原三省草甘膦被广泛使用以实现零耕作,也经常用于休闲地以及作物种子发芽前灭生性除草,这导致对抗草甘膦地肤的选择,且其迅速扩展,对加拿大草原三省造成大的经济影响。
1.6 其他国家
以上提及的所有国家中除了澳大利亚都大面积种植抗草甘膦作物。在22个其他国家也发现了抗草甘膦杂草。其中大部分国家没有种植抗草甘膦作物,而剩余国家种植抗草甘膦作物非常少,或/和种植时间非常短(例如玻利维亚)。巴拉圭种植的抗草甘膦作物面积最大,2015年为 370万 hm2。因此在这 22个国家中巴拉圭抗草甘膦杂草品种数量增长最快,与其接壤的国家阿根廷和巴西一直也存在抗草甘膦杂草问题。
这 22个国家的数个抗草甘膦杂草的案例引人注意。例如,第2个报道的抗草甘膦杂草事例为在1997年在马来西亚果园发现的抗草甘膦牛筋草(图1)。马来西亚没有种植抗草甘膦作物,也再没有第2例抗草甘膦杂草的报道。
2 杂草科类
Holm等人发表了“世界恶性杂草”一文,根据此对含有抗草甘膦杂草品种数量最多的杂草科进行列表(表3)。其中也列出了每一杂草科中抗草甘膦杂草品种的数量(表3)。通过对比表格各列可发现4科杂草(禾本科、菊科、苋科、藜科)对草甘膦较易产生抗性(表 3)。
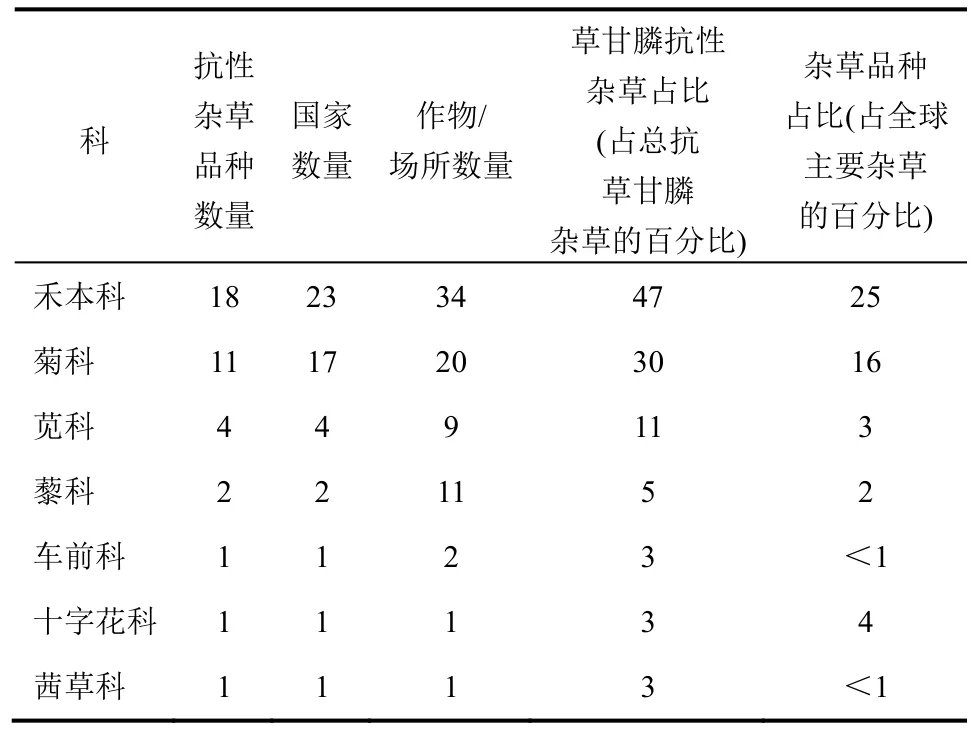
表3 不同科的抗草甘膦杂草情况
2.1 禾本科
草甘膦一般对禾本科杂草的活性要高于其他主要杂草科。有趣的是对草甘膦产生抗性的禾本科杂草比其他科杂草多。禾本科杂草量占世界主要杂草的25%,而抗草甘膦禾本科杂草为所有抗草甘膦杂草的47%。因此,在剂量-反应曲线中禾本科杂草通常接受到的草甘膦剂量要高于非禾本科杂草。抗草甘膦禾本科杂草数量的不成比例性可能是由于禾本科杂草更易对草甘膦产生抗性,暴露于草甘膦的禾本科杂草数量越多,和/或相对高剂量能更有效选择抗草甘膦杂草个体。高剂量能成功地选择抗草甘膦禾本科杂草与低剂量能更有效地选择抗性的观点相反。对于禾本科杂草来说,黑麦草属、虎尾草属和雀麦属杂草易产生草甘膦抗性,在17个抗草甘膦杂草中占8个。抗草甘膦黑麦草属草(Lolium sp.)、两耳草、牛筋草、阿拉伯高粱和光头稗分布最广,经济影响最大。
2.2 菊科
抗草甘膦菊科杂草也较多,16%的世界恶性杂草属于菊科(表 3),菊科杂草为抗草甘膦杂草的30%。白酒草属和豚草属(Ambrosia spp.)杂草特容易产生草甘膦抗性,几乎为抗草甘膦菊科杂草的一半。
2.3 苋科
在苋科的163个属中只有苋属对草甘膦产生抗性。4个苋品种杂草已对草甘膦产生抗性[长芒苋、糙果苋、绿穗苋(A. hybridus)和刺苋]。抗草甘膦长芒苋是对经济影响最大的抗草甘膦杂草,糙果苋正快速发展将造成严重的影响,部分原因为这2种杂草已对防治所用的大部分主要作用位点除草剂产生抗性。
2.4 藜科
抗草甘膦地肤和刺沙蓬(Salsola. tragus)是对草甘膦产生抗性的2种藜科杂草。抗草甘膦地肤具有有效的滚风草种子扩散机制而迅速地扩展,目前是主要影响美国大平原和加拿大草原三省经济的杂草。刺沙蓬也是滚风草,最近抗草甘膦刺沙蓬在美国蒙大拿州和俄勒冈州被确定。其扩展机制是否如地肤一样,还有待验证。
3 影响草甘膦抗性的发生和扩展的因素
3.1 抗性杂草的扩展与选择
研究者发现乙酰辅酶A羧化酶(ACCase)和乙酰乳酸合酶(ALS)抑制剂抗性杂草相对常见,部分原因为每一靶标位点发生数个突变,使杂草具有高水平抗性。这2个靶标位点具有许多有效的除草剂作用点,被认为具有可塑性,能够相对快速选择抗性。大多数ACCase和ALS抑制剂抗性杂草是由于一个碱基对突变导致高水平抗性(>10)。相比较,EPSP基因几乎不发生赋予低水平草甘膦抗性的 1 bp突变,使抗草甘膦杂草的最初出现频率很低。草甘膦是仅有的抑制EPSPS的商业化除草剂,其结合点具有非常高的特异性。对于抗草甘膦杂草,在农业生产实践中需要注意的是其抗性的扩展作为新侵染的来源比现有杂草种群中新抗性事例的产生更重要。对于ACCase和ALS抑制剂抗性事例,最初高水平抗性基因频率意味着严格的检疫(清洁机械、货车、牲畜等)不是有效的避免抗性的方法。因为适宜突变发生频率,很可能抗性个体已经在田间天然杂草种群中存在,或不久发生。然而,Bradshaw等人在讨论为什么草甘膦抗性不可能发展中指出,杂草种子卫生学对限制选择抗草甘膦杂草非常重要,因为最初非常低的抗草甘膦基因频率可能意味着天然种群中一定水平的抗性不存在,且在相当长的时间内在任何特定田间不可能发生,甚至在一年一年暴露于草甘膦的情景下。
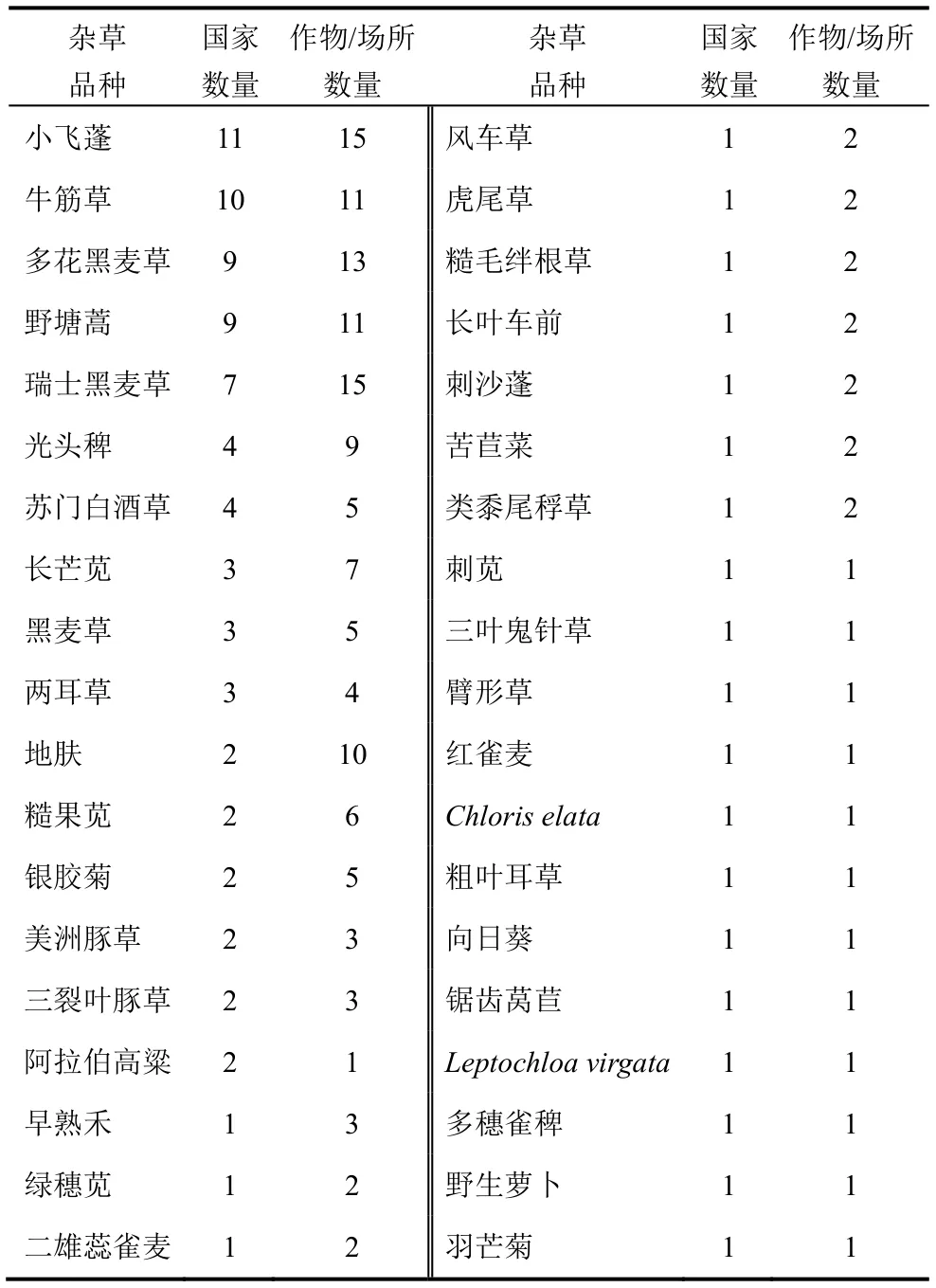
表4 全球抗草甘膦杂草品种发生情况
3.2 抗草甘膦杂草的选择:低剂量和高剂量
曾在多年期间,杂草科学家一直在争论是高剂量(意味高的选择压)还是低剂量(低选择压)导致杂草抗性的快速发展。答案为二者都是。对于赋予非常高水平抗性(常见于对ALS和ACCase抑制剂靶标位点抗性)的抗性机制,当抗性特性显性,高剂量除草剂能够快速杀灭敏感个体,在杂草种群中更快地产生抗性。对于不能赋予高水平抗性的机制,高剂量除草剂将导致携带抗性特性的个体的死亡或生长显著受抑,而低剂量则更可能导致抗性的产生。此外,低剂量除草剂可能使低水平草甘膦抗性特性异型杂交和组合到1个个体,使此个体能够在高剂量草甘膦下存活(creeping resistance)。大部分的抗草甘膦机制为低水平抗性类别,因此避免或延缓草甘膦抗性的策略为在理想的应用条件下使用最大推荐剂量杀灭最多的杂草。这样的话,许多具有低水平抗性机制的个体在通过异型杂交合并不同抗性基因前可能就被杀灭,只剩下较高水平草甘膦抗性个体。如上面讨论一样,此理论显而易见不适用于禾本科杂草,即使所用草甘膦剂量要高于用于大部分其他杂草品种的,抗性禾本科杂草品种数量一直多。
3.3 自花授粉和异型杂交
因为大多数抗草甘膦抗性机制(染色体易位、整合和基因扩增)为低水平抗性。杂草品种具有一定程度的异型杂交特性是其一大优势,这样就具有合并的能力而产生对草甘膦高的抗性。具有单独低水平机制的杂草个体可以存活,但应用草甘膦可能会严重压制其,产生的种子要少于含有2个或多个抗草甘膦机制的个体。由此,如果确定异型杂交植物已产生抗性后继续使用草甘膦防除其他杂草品种,此植物可能继续会发展高水平草甘膦抗性。
3.4 多抗
如果杂草只对草甘膦有抗性,此问题不严重,可用老的除草剂,或老的除草剂与草甘膦混用或轮用进行防除。草甘膦可防治多种杂草,老的除草剂特异性防除抗草甘膦杂草。然而,这种情况比较罕见。种植者快速接纳了抗草甘膦作物,部分原因为防除对其他除草剂产生抗性的杂草,特别是抗ALS、ACCase和三嗪类除草剂杂草。最初,使用新引入的除草剂来防除抗性杂草。在20世纪80年代和90年代混用或直接用 ALS和 ACCase抑制剂防治在 20世纪70年代和80年代产生的抗三嗪和抗二硝基苯胺杂草。相应地,用原卟啉原氧化酶抑制剂,或在抗草甘膦作物田中用草甘膦防治抗ALS和ACCase抑制剂杂草。现在,没有新的作用机制除草剂或很少的新除草剂引入市场,数量不断增加的抗草甘膦杂草的防治就成为难题。
4 抗性机制
4.1 靶标点抗性
已确定数种类型的草甘膦靶标点抗性。顾名思义,靶标点抗性是由于靶标酶(EPSPS)的改变而产生草甘膦抗性。靶标点抗性可由一个碱基对的改变、多个碱基对的改变、密码子缺失或基因的扩增而所致。到目前为止,除密码子缺失外,已发现了其他机制的抗草甘膦杂草。
4.1.1 一个碱基对的改变
EPSPS基因中某一碱基对的变化可使此酶对草甘膦有微弱的抗性,使杂草对草甘膦产生低水平抗性(一般2~6×)。然而,不常见的是单一碱基对机制合并非靶标点机制,产生较高倍数的抗性。赋予抗性的 EPSPS的所有单一碱基对的突变导致 Pro106或 Ser、Ala、Thr或 Leu的改变。已报道牛筋草、瑞士黑麦草、 多花黑麦草、两耳草、糙果苋和光头稗发生这些突变。最近报道帚状虎尾草(C. virgata)的Pro 106被Ser和Leu替代。此研究中,发生Pro 106Leu突变的植物的抗性约为 Pro 106Ser突变的3~5倍,和EPSPS离体抗性相一致。没有发现其他抗性机制。相似地,在美国加州的光头稗中发现Pro 106密码子发生突变的2个不同的EPSPS突变体。此杂草品种具有2个EPSPS基因,2个突变发生于不同的基因上。Pro 106Ser突变的抗性要低于 Pro 106Thr突变,这与相对离体抗性一致。但是,此种群可能也有其他抗性机制,因为此研究报道的1个光头稗生物型没有发生EPSPS突变,但仍具有低水平草甘膦抗性。
4.1.2 多碱基对改变
EPSPS基因发生2个碱基对改变,导致EPSPS中2个氨基酸变化,这可能导致的草甘膦抗性比Pro 106突变的高。野生型 EPSPS发生改变的含有Thr102lle和Pro 106Ser转基因EPSPS(TIPS双突变)已被用于开发出商业化的抗草甘膦转基因玉米品种。抗草甘膦牛筋草已具有TIPS形式的EPSPS。双突变的植物具有180倍的抗性,相当于或高于抗草甘膦作物的抗性水平,与ALS和ACCase抑制剂抗性杂草的靶标点抗性可媲美。单独Thr102lle突变产生非常无效的 EPSPS,故先行Pro106Ser突变,然后Thr102lle突变。孟山都认为双突变不可能发生时,它没有想到此类型依序发生的高水平抗性。在草甘膦对单突变抗草甘膦生物型持续的选择压下,TIPS突变将最终在这些抗性种群中发生。
4.1.3 基因扩增或复制
基因扩增或复制增加了基因拷贝数,由此分子靶标或解毒酶的量增加了。EPSPS的增加将需增加能充分抑制EPSPS杀灭植物的草甘膦的量。自从此抗性机制首次在长芒苋中报道后,EPSPS基因的复制已与全美国的数个长芒苋种群对草甘膦的抗性有关。到目前,已报道了糙果苋、刺苋、西部苋(A.rudis)、多花黑麦草、地肤、二雄蕊雀麦(B. diandrus)、牛筋草和C. truncata的抗草甘膦生物型的EPSPS扩增。对于抗草甘膦刺苋,基因扩增可能是由于刺苋与EPSPS扩增的长芒苋杂交所致。EPDPS基因的扩增一般赋予比EPSPS的1 bp突变更高的抗性,虽然额外EPSPS拷贝数量和抗性水平可变,额外拷贝数可小为 3,有时大于 100。EPSPS拷贝数、EPSPS酶的数量和整个植物水平抗性间的一致性较好。没有与EPSPS扩增有关的明显的适合度代价。
所有的靶标点机制可发生于或一起发生于同一杂草品种。例如,Chen等人发现牛筋草存在单和双突变EPSPS,以及EPSPS的基因扩增。他们也发现抗草甘膦生物型牛筋草中磷酸果糖激酶的基因扩增,也有EPSPS扩增。
4.2 非靶标点抗性
对于非靶标抗性,EPSPS酶仍然对草甘膦敏感,而是到达EPSPS酶的草甘膦量降低的突变机制导致抗性的产生。许多草甘膦抗性事例是由于非靶标点抗性所致。非靶标点抗性机制通常赋予3~12倍的低水平抗性。
4.2.1 代谢增加
草甘膦被一些杂草和作物代谢为乙醛酸和氨甲基膦酸(AMPA)。一些植物也可把草甘膦代谢为肌氨酸和无机磷酸盐。AMPA的植物毒性非常弱,乙醛酸为天然植物代谢物。因此提高草甘膦代谢为AMPA的量,草甘膦就不会伤害杂草。把草甘膦代谢为AMPA的草甘膦氧化环氧酶(GOX)的基因已被转入油菜,使油菜对草甘膦有耐受性。然而,此转基因作物也含有抗性微生物EPSPS基因(cp4),还不清楚GOX基因对草甘膦抗性的贡献多大。只有cp4的大豆的抗性水平(~50×)约和同时还有 cp4和 gox的油菜一样。在草甘膦应用后一些植物中发现的AMPA的量表明降解可能导致对草甘膦的天然耐受性。但这还没有经过严格的证明。例如,天然抗草甘膦杂草 Ipomoea lacunosa能把草甘膦代谢为AMPA,但草甘膦的代谢量和杂草的耐受性水平间没有相关性。笔者认为还没有确切证据证明提高草甘膦的降解为抗草甘膦杂草的抗性机制。一些植物具有代谢草甘膦的基因以及草甘膦为作用慢的除草剂,但令人惊奇的是增加草甘膦降解不是常见的抗性机制。
4.2.2 降低吸收和/或迁移
降低除草剂的吸收或迁移,使到达作用位点的量不足以杀灭植物,这样就会产生抗性。降低从植物表皮进入植物细胞的草甘膦的移动(除草剂吸收),可能使杂草对草甘膦产生抗性。阿拉伯高粱、多花黑麦草、两耳草、C.eleta和Leptochloa virgata的一些抗草甘膦生物型具有此种机制。吸收降低可能是杂草叶表皮化学和/或形态学改变所致,或叶形状或方位的变化增加了除草剂喷雾液的流失所致。
草甘膦有效的1个原因是其易从叶迁移到分生组织。因此降低此迁移量将会降低草甘膦的防效。一些研究报道抗草甘膦生物型杂草对草甘膦的迁移降低了。然而,最近才开始了解此机制。被处理叶细胞的液泡扣留草甘膦能力的提高是一些抗草甘膦植物对草甘膦迁移降低的1个原因。
4.2.3 滞留
除草剂滞留于植物的一些组织中(液泡,或细胞壁的吸附),不能迁移和到达植物细胞的叶绿体作用位点,这样植物就对草甘膦产生抗性。小飞蓬和黑麦草属对草甘膦的抗性是因为草甘膦很快滞留于液泡中。对于这些抗性事例,液泡膜草甘膦运输者的活性和/或水平提升了。此运输者具有 ATP结合盒(ABC)转运子的特性。当小飞蓬抗性生物型暴露于草甘膦后,其2个主要的ABC转运子基因上调了。在较低的温度,抗性小飞蓬的快速液泡扣留机制不能运行,因为在较低温度液泡扣留不会发生。
4.2.4 “长生鸟”现象
草甘膦作为除草剂作用非常缓慢,一些三裂叶豚草生物型的叶子在被喷雾处理后能够对草甘膦迅速做出反应,在处理后数小时内干枯,除草剂不能从死的组织迁移,发育的组织和分生组织能够再生长,产生“类长生鸟”恢复现象。整株植物抗性水平为2.3~7.4倍。草甘膦对此生物类型的快速作用机制未知,虽然外源芳香氨基酸可抑制此种作用,这表明此作用与对莽草酸途径的抑制有关。此外,用草甘膦处理的抗性生物型的叶碟中活性氧迅速积累。此快速反应需要光或外源蔗糖。
4.3 多个机制
即使草甘膦抗性风险相对低,其抗性还是在快速发展。但是,与其他任何类型的除草剂不一样,长期以来草甘膦的选择压存在于大面积的田中。草甘膦抗性进化机制也比其他作用机制的除草剂多样化。
研究较多的抗草甘膦杂草品种一般具有数种抗性机制,同一品种的不同抗性生物型抗性机制组合不同。如Sammons和Gaines指出一样,对ACCase和ALS抑制剂除草剂有靶标位点抗性的仅TIP突变草甘膦抗性杂草具有高水平抗性(>50×),在大多数情况下草甘膦抗性进化是对低剂量抗性机制的选择过程。在足够长的选择期间,抗性机制趋于在1个杂草品种中积累,增加总的抗性因子。随着时间的流逝,由于抗性机制的堆积所致的缓慢发展的抗性可以产生很强的抗性,使杂草免受在经济上或法规上可使用的最高剂量草甘膦的危害。
5 未 来
除草剂已为人类提供了最有效和经济的杂草防治方法约70年,如果不存在抗草甘膦杂草,主要农艺作物田中的杂草防治问题基本解决。草甘膦被迅速用于抗草甘膦作物,部分原因是因为使用的除草剂出现了抗性问题,特别是 ALS、ACCase和光合系统Ⅱ(PSⅡ)抑制剂的抗性。抗草甘膦作物的成功一定程度上导致农业化学工业没有动力来研究和开发新作用位点除草剂。草甘膦的广泛使用导致了抗草甘膦杂草品种不断地产生,危害更大的抗性杂草迅速扩展。此问题对农民来说更严重,因为可代替性除草剂在不断减少,最主要因为这些抗性杂草已经对老的除草剂具有抗性特性且产生新的抗性。多重抗性((草甘膦和其他除草剂)以及新作用位点除草剂的缺乏,给现代农业带来杂草管理危机。此外,瑞士黑麦草等禾本科杂草非靶标点多重抗性(主要为代谢)和大穗看麦娘(Alopecurus myosuroides)给除草剂抗性增加了复杂性。随着此类型多重抗性在阔叶杂草中的扩展预,预期在未来30年此问题的重要性将增加。非靶标位点抗性管理比较困难,因为其经常呈现与不同作用位点除草剂的不可预期的多重抗性,因此不同作用位点除草剂的轮用可能不能有效治理非靶标位点抗性。
尽管有以上情况,除草剂仍然是发达国家杂草防除的中流砥柱,且在被新的技术逐步淘汰前也会如此。但需要以持续的方式使用除草剂,不仅要合理混用和轮用,而且要作为含有机械和栽培措施的集约化综合杂草管理项目的一部分。抗草甘膦杂草的品种和侵染的作物面积将继续增加,但抗性管理可延缓其增加。
即使除草剂的发现研究已停止,但目前多抗杂草的严重和缺乏有效替代防治产品给农化公司带来了开发新除草剂的强大的经济刺激。未被开发的作用机制的除草剂可能将吸引较多投资研究。新转基因、诱变育种和基因编辑抗除草剂作物特性的开发和引入可能继续使老的除草剂以新的方式使用。作物中抗除草剂堆积形状将标准化,即意味这些作物的种植可使种植者根据情况决定除草剂的混用,延缓多重抗性的出现。RNAi技术可能有助于管理抗性杂草,然而,所面临的技术和经济问题可能使其商业化还遥遥无期。杂草防治技术的多样化可减少抗性直接进化,延长有效除草剂的生命货架,可给研究人员足够时间开发新的杂草防治技术。
最后,除草剂将可能被新技术取代。转基因微生物防治剂和作物化感作用使这些杂草管理方法更有效、经济和有选择性。随着计算机技术的快速发展,可通过图像分析增加确定杂草的速度和准确度,再随着机器人技术的快速发展可能在未来 20年中实现精确地应用除草剂和/或机器人经济地防除许多作物田中的杂草。此外,随着以上各个方法的进步,将这些技术结合现有杂草防除方法,包括除草剂,使杂草防除多样化,使杂草防除法的抗性进化最慢。
6 结 论
除草剂曾经被视为主要作物杂草防除的主要解决方案,但由于抗性问题和环境问题,其生命期有限。抗草甘膦作物在短期内被种植者迅速接受,抛弃了复杂的杂草防治策略,因为用草甘膦防除杂草操作简单,费用低且有效,使种植者节约了数以亿计的杂草管理费用。然而过分依赖草甘膦造成草甘膦抗性杂草的产生,以及杂草防除技术缺乏多样化和没有开发出新作用位点的除草剂,目前杂草防除正处于广泛的失败边缘。随着多重抗性杂草的增加,种植者将不得不采用更复杂的综合杂草管理项目来防除杂草。他们应用复杂杂草防除项目的程度将决定延迟除草剂防除杂草彻底失败到来的时间。笔者希望实用的、新的杂草防除技术出现且在除草剂防治失败成为灾难前能有效应用,但这并不是说,一旦新技术被应用,就可以放低警惕,因为不论应用何种杂草防除策略,杂草都将会对其发展抗性,这也就是为何有必要实施综合杂草管理项目的原因。
2023年全球杀幼虫剂市场有望达到9.5亿美元
根据某调研机构的报告,2017年全球杀幼虫剂市场价值为7.2亿美元,2018年预计为7.5亿美元,预计将以4.86%的复合年增长率增长,至2023年达9.5亿美元。
