西宁盆地黄土区典型草本植物单根抗拉力学特性试验
2018-08-21刘亚斌李淑霞余冬梅胡夏嵩杨幼清
刘亚斌,李淑霞,余冬梅,胡夏嵩,杨幼清
(1.中国科学院青海盐湖研究所中国科学院盐湖资源综合高效利用重点实验室,西宁810008;2.青海省盐湖地质与环境重点实验室,西宁810008;3.中国科学院大学,北京100049;4.青海大学地质工程系,西宁810016)
0 引 言
近年来随着西宁盆地及其周边地区经济快速发展,区内基础设施建设力度亦不断加大。随着大规模工程建设活动的开展,诸如边坡开挖等工程活动会在一定程度上造成原有斜坡植被的破坏,并形成大量裸露边坡,其结果是进一步加剧了区内水土流失、滑坡、泥石流等地质灾害现象的发生,同时也对区内地质环境与生态环境产生了直接影响和破坏作用[1-4]。据已有国内外相关学者研究成果表明,通过种植植被的方法不仅可起到边坡生态恢复的作用,同时还能有效地控制水土流失、浅层滑坡及泥石流等地质灾害现象的发生[5-9]。植物通过其根系力学效应可实现有效增强边坡土体抗剪强度和提高边坡稳定性作用,主要归因于根系具有较强的抗拉特性,而土体具有抗压能力强而抗拉能力弱的特性,故当含根系土体受到剪切作用时,通过根-土界面摩擦力的作用,根系可将土体内部剪应力转化为自身所承受的拉应力,起到增强土体抗剪强度的作用[10]。植物根系抗拉特性是表征其增强土体抗剪强度能力大小的重要力学特性[10-13]。
有关植物根系抗拉力学特性的研究,国内外学者开展了大量相关试验工作。Stokes等[14]指出根的抗拉强度愈高则愈具有抗拔出和增强土体抗剪强度的能力,故在根面积比一定的条件下,数量相对愈多的细根较数量相对较少的粗根,表现出具有相对更为显著的固土护坡能力。Baets等[10]研究了生长于地中海地区的25种植物(含灌木、草本和小型乔木)的单根抗拉强度,研究结果认为植物单根抗拉强度随着根径增加呈降低的趋势,且单根抗拉强度与根径之间的函数关系取决于植物的种类。钟荣华等[15]通过室内单根拉伸试验对三峡库区消落带牛鞭草、扁穗牛鞭草、双穗雀稗及狗牙根 4种草本植物进行单根拉伸试验研究,结果表明 4种草本植物单根平均抗拉力由大至小依次为狗牙根、牛鞭草、扁穗牛鞭草和双穗雀稗;而单根平均抗拉强度由大至小依次为狗牙根、扁穗牛鞭草、牛鞭草和双穗雀稗;4种草本植物单根抗拉力和单根抗拉强度,随着根径增加分别呈幂函数增大和幂函数降低趋势。此外,Tosi等[11,16-22]等学者亦对不同草本、灌木及木本植物的单根抗拉力、单根抗拉强度与根径之间的关系进行了研究,并得出了相类似的研究结果。
有关植物根系应力-应变关系研究方面,刘国彬等[16]通过单根拉伸试验对黄土高原常见的沙打旺等12种草本植物根径为0.1~1.0 mm的毛根应力-应变关系、弹性模量等力学特性进行了系统研究,认为不同草本植物的本构方程和弹性模量均存在差异性,且毛根应力-应变关系对应变率的敏感程度,随根径的增加表现出呈降低趋势。刘秀萍等[23]通过分析油松单根拉伸试验过程中表现出的应力与应变关系特征,指出当油松单根应力达到抗拉强度极限的 50%~70%时,所对应点即为单根的弹性极限点。朱海丽等[1]通过对种植于西宁盆地生长期为18个月的柠条锦鸡儿、霸王等4种灌木进行室内单根拉伸试验,结果表明4种灌木单根在受拉初期其应力-应变曲线呈线性关系,表现出弹性材料的特征,当载荷超过弹性极限时,4种灌木应力-应变关系表现出弹塑性材料的特征。
综上所述,有关植物根系抗拉力学特性研究方面,已有研究成果主要表现在对不同植物单根抗拉力学特性指标差异性评价及其与根径关系,以及单根应力-应变关系分析等方面。其中,有关单根抗拉力、单根抗拉强度等单根抗拉力学特性指标差异性评价时,多数采用的是对不同根径单根,其平均单根抗拉力学特性指标进行对比分析,相应地,以根径级别作为分类依据,对西宁盆地黄土区不同草本植物开展不同根径级别条件下,单根抗拉力、单根抗拉强度等单根抗拉力学特性指标对比分析等方面的研究则相对较少;与此同时,已有的有关西宁盆地黄土区植物单根拉伸应力-应变特征方面的研究,主要以灌木植物为主要研究对象[1,17],而探讨区内不同草本植物单根应力-应变关系反映出的阶段性特征及其差异性方面的研究亦相对较少。基于上述分析,本项研究选取西宁盆地典型草本植物垂穗披碱草(Elymus nutans Griseb.)、细茎冰草(Agropyron trachycaulum Linn.Gaertn.)及紫花苜蓿(Medicago sativa L.)作为试验供试种,通过开展单根拉伸试验对这 3种草本植物单根抗拉力、单根抗拉强度和单根极限延伸率 3个单根抗拉力学特性指标大小及其与根径之间的关系,以及单根拉伸过程中的应力-应变特征进行了试验研究。本项研究结果对于西宁盆地及其周边黄土区边坡水土流失、浅层滑坡等地质灾害现象有效防治,以及保护区域生态地质环境等方面均具有理论研究价值和实际指导意义。
1 研究区概况
研究区为地处青海东北部湟水谷地中部的西宁盆地,属于黄土高原西缘与青藏高原东北部的交接地带。区内为温带大陆性气候,年平均气温为6 ℃,年平均蒸发量为1 400~2 000 mm,年均降水量为250~500 mm且年内分布极不均匀,降雨主要集中分布在6~9月份,约占全年降水量的70%~80%,多以暴雨和阵雨形式出现,具有历时短、强度大、降雨集中等特点[24]。西宁盆地及其周边地区地表植被覆盖率相对较低,黄土高陡边坡发育,地质环境相对脆弱,区内地质灾害类型主要为滑坡、崩塌、泥石流3种类型[25-26]。本项研究的自建试验区位于西宁盆地长岭沟,其地理坐标为东经101°42′00′,北纬36°36′00′,区内海拔为2 315~2 570 m。种植区属于土质边坡,边坡坡度为30°,坡向向南,土质类型为粉土。
2 试验材料与方法
2.1 试验供试种种植方法及供试种选择
本项研究在自建试验区共划分出10块坡长为1 000 cm,坡宽为500 cm的试验种植区,其中的3块种植区(即6号、7号、8号种植区)用于单播种植。3块单播种植区内均设置有11块坡长为1 000 cm,坡宽为45 cm的种植小区。3个单播种植区内草本和灌木植物种植方法采用随机完全区组设计,每个种植区内均种植有10种适宜当地气候条件下生长的草本和灌木植物。每块种植区内草本和灌木植物种植顺序通过随机抽样的方法确定。种植区内种植的草本植物包括垂穗披碱草、细茎冰草、紫花苜蓿、芨芨草(Achnatherum splendens(Trin.)Nevski)、草地早熟禾(Poa pratensis L.)和星星草(Puccinellia tenuiflora(Griseb.)Scribn.)6种,本项研究选取生长期为150 d时,生长情况相对良好的垂穗披碱草、细茎冰草、紫花苜蓿3种草本植物,作为试验供试种进行单根拉伸试验。试验区内草本植物种植方法为条播法,播种量为 3.8~20.1 g/m2。试验区草本和灌木植物种植方案如图1所示。

图1 试验区草本和灌木植物种植方案Fig.1 diagram of planting plots and planting program in test area
本项研究所选取的 3种草本植物中,垂穗披碱草与细茎冰草为禾本科多年生草本植物,紫花苜蓿为豆科多年生草本植物。垂穗披碱草和紫花苜蓿为适宜西宁盆地及周边地区环境条件下生长的常见乡土植物,可在 2 200~4 100 m的海拔范围内正常生长[27-30];细茎冰草为引进至西宁盆地的优良牧草种,该种亦适宜于西宁盆地及其周边地区种植生长,具有较强的抗寒抗旱性和适应性[31]。这 3种草本植物均为研究区人工草地建植、水土保持的理想草种[27,31]。本项研究所选取的垂穗披碱草、细茎冰草和紫花苜蓿3种草本植物生长期均为150 d。3种草本植物中,垂穗披碱草与细茎冰草根系为须根型,紫花苜蓿根系发达且多为直根型[32-33]。生长期为150 d时,垂穗披碱草与细茎冰草根系主要分布在边坡地表以下15~25 cm深度土体中;紫花苜蓿根系可达边坡表层以下50~75 cm深度位置。试验区植物整体生长情况,及3种草本植物生长情况如图2所示。

图2 试验区植物生长情况Fig.2 Growth situation of vegetation in test area
2.2 取样与制样方法
在区内3种草本植物单根拉伸试验之前,首先分别从每种草本植物在6号、7号和8号种植区所对应的种植小区的上部、中部和下部位置(取样位置如图1所示),整株挖掘出试验所需3种草本植物根系各一组,并将原位采集完毕的根系样品放入大号自封袋中随即带回实验室进行试验。在室内将3种草本植物根系用清水洗净后,分别从每种草本植物的3组根系试样中用剪刀各随机剪取70根根长为8~10 cm且未受损伤的单根,即每种草本植物共剪取210根单根进行拉伸试验。因根径<l.00 mm的根系缠绕固结土体、增强土体抗冲性作用相对较为显著[16],同时,鉴于紫花苜蓿单根根径为0~0.10 mm范围内的数量相对较少,且垂穗披碱草和细茎冰草单根根径≥0.6 mm条件下的数量亦相对较少,故本项研究单根拉伸试验所选取的单根根径范围为0.10~0.60 mm。此外,为系统性对比分析区内3种草本植物根系力学指标之间的差异性,本项研究将3种草本植物根径D(mm)依次划分为(0.10,0.20]、(0.20,0.30]、(0.30,0.40]、(0.40,0.50]及(0.50,0.60] mm 5个根径级别进行单根拉伸试验。
2.3 单根拉伸试验方法
区内3种草本植物在开展单根拉伸试验时,首先将单根夹持在上、下拉伸夹具之间,同时保证位于上、下两夹具之间的根段标距为20 mm(夹具间根段保持顺直状态,同时做到避免根段在拉伸试验开始前受拉力作用);试验过程中,上夹具以20 mm/min的速率匀速上行,以使位于上、下夹具间的根段承受拉伸作用,直至该根段被拉断为止。由于单根拉伸试验仪在记录单根应力-应变曲线时,需提前测得待测单根根径,故在每根单根进行拉伸试验前,先用记号笔在其中部位置做出2个标记(标记间距为20 mm),然后在2个标记间的3个不同位置处使用游标卡尺测量其根径,以3次测量的平均值作为该20 mm长根段根径,最后按单根根径级别依次进行相应归类。此外,在夹持单根时,做到准确地将2个标记间的根段夹持在夹具之间。单根拉伸试验进行过程中,单根应力-应变关系曲线则由该试验仪内置程序自动计算生成并记录;待试验结束后该试验仪自动记录单根抗拉力;然后在此基础上计算得到单根抗拉强度、单根极限延伸率等单根抗拉力学特性指标。
此外,需要说明的是,本次单根拉伸试验过程中,选取位于上、下两夹具中间位置处发生断裂根段的单根抗拉力学特性指标作为试验有效数据。试验采用的仪器为上海衡翼精密仪器有限公司生产的HY-0580型微机控制电子万能试验机。电子万能试验机力学传感器量程为10 kg,精度为0.3%。本项研究中3种草本植物单根抗拉强度计算公式为[1]

3种草本植物单根极限延伸率计算公式为[1]
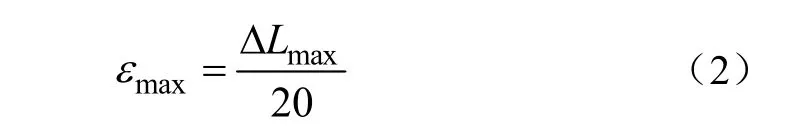
式中Fmax表示单根抗拉力,N;D表示上、下两夹具间根段的平均根径,mm;T表示单根抗拉强度,MPa;ΔLmax表示单根处于极限状态时的根长增长量,mm;εmax表示单根极限延伸率,%。
3 试验结果与分析
3.1 3种草本植物单根抗拉力学特性
3.1.1 单根抗拉力
试验区3种草本植物单根抗拉试验结果如表1所示。由表1可知,区内3种草本植物根径在(0.10,0.20]、(0.20,0.30]、(0.30,0.40]、(0.40,0.50]及(0.50,0.60] mm 5个根径级别条件下,紫花苜蓿单根抗拉力均显著高于垂穗披碱草和细茎冰草。在上述 5个根径级别范围内,紫花苜蓿单根抗拉力为(2.28±0.67)~(12.00±3.89)N,垂穗披碱草和细茎冰草单根抗拉力分别为(1.75±0.43)~(7.66±0.72)N 和(1.82±0.46)~(7.95±0.47)N,紫花苜蓿单根抗拉力较垂穗披碱草和细茎冰草分别平均高1.32倍和1.26倍。此外,区内3种草本植物根径级别为(0.20,0.30] mm时,紫花苜蓿单根抗拉力较垂穗披碱草、细茎冰草的差异性相对较小,即紫花苜蓿单根抗拉力分别为垂穗披碱草和细茎冰草的1.18倍和1.16倍;当根径级别为(0.50,0.60] mm时,紫花苜蓿单根抗拉力较垂穗披碱草、细茎冰草表现出相对较大的差异性,即紫花苜蓿单根抗拉力分别为穗披碱草和细茎冰草的 1.57倍和1.51倍。此外,区内细茎冰草单根抗拉力在 5个根径级别条件下均表现出略大于垂穗披碱草的特性,且细茎冰草单根抗拉力平均高出垂穗披碱草1.05倍,同时在5个根径级别条件下,垂穗披碱草和细茎冰草单根抗拉力的差异性均表现出相对不显著性的特性。

表1 试验区3种草本植物单根抗拉力学特性指标试验结果Table 1 Single root tensile mechanical property indexes of three herbs in test area
3.1.2 单根抗拉强度
由表1可知,区内3种草本植物在5个根径级别条件下,紫花苜蓿的单根抗拉强度显著高于垂穗披碱草和细茎冰草,即具体表现为 5个根径级别条件下,紫花苜蓿单根抗拉强度为(49.21±16.39)~(124.88±46.63)MPa,垂穗披碱草和细茎冰草单根抗拉强度分别为(35.33±1.76)~(84.87±15.93)MPa和(35.96±2.46)~(73.32±19.56)MPa,紫花苜蓿单根抗拉强度分别较垂穗披碱草和细茎冰草平均高出1.30倍和1.31倍。其中,当根径级别为(0.30,0.40] mm时,紫花苜蓿单根抗拉强度与垂穗披碱草、细茎冰草之间的差异性均表现出相对较小的特性,即紫花苜蓿单根抗拉强度分别为垂穗披碱草和细茎冰草的1.18倍和1.12倍;相应地,当根径级别为(0.10,0.20] mm时,紫花苜蓿的单根抗拉强度较垂穗披碱草、细茎冰草表现出具有相对较大的差异性,即紫花苜蓿单根抗拉强度分别为垂穗披碱草和细茎冰草的 1.47倍和1.70倍。虽然垂穗披碱草和细茎冰草的单根抗拉强度在 5个根径级别条件下均未表现出显著差异性,但在根径级别为(0.10,0.20] mm和(0.20,0.30] mm时,垂穗披碱草单根抗拉强度具有略大于细茎冰草的特性,在上述 2个根径级别条件下,垂穗披碱草单根抗拉强度分别为细茎冰草1.16倍和1.01倍;与此同时,在根径级别为(0.30,0.40]、(0.40,0.50]及(0.50,0.60] mm 3 个根径级别条件下,细茎冰草的单根抗拉强度表现出略高于垂穗披碱草的特性,即分别高出1.05倍、1.09倍和1.02倍。
3.1.3 单根极限延伸率
由表1可知,区内3种草本植物在5个根径级别条件下,垂穗披碱草单根极限延伸率表现出显著高于其他2种草本植物的特性,细茎冰草次之,而紫花苜蓿的单根极限延伸率在 5个根径级别条件下均具有相对较小的特性。区内3种草本植物在5个根径级别条件下,垂穗披碱草的单根极限延伸率为 50.36%±15.52%~76.4%±3.05%,细茎冰草和紫花苜蓿的单根极限延伸率则分 别 为 45.84%±11.56% ~ 73.28%±10.82% 和25.74%±8.05%~36.09%±7.78%,垂穗披碱草单根极限延伸率分别较细茎冰草和紫花苜蓿平均高 1.13倍和 2.16倍。当根径级别为(0.50,0.60] mm时,垂穗披碱草单根极限延伸率较细茎冰草差异性相对不显著,即垂穗披碱草单根极限延伸率仅为细茎冰草的1.02倍;相应地,当根径级别为(0.10,0.20] mm时,垂穗披碱草单根极限延伸率较紫花苜蓿的差异性相对较小,即垂穗披碱草单根极限延伸率为紫花苜蓿的1.75倍;在根径级别为(0.30,0.40] mm时,垂穗披碱草单根极限延伸率分别较细茎冰草和紫花苜蓿表现出相对较大的差异性,即垂穗披碱草单根极限延伸率分别为细茎冰草和紫花苜蓿的1.19倍和2.51倍。此外,在 5个根径级别条件下,细茎冰草单根极限延伸率亦表现出较紫花苜蓿显著的特性,即细茎冰草单根极限延伸率平均高出紫花苜蓿1.91倍,其中根径级别为(0.10,0.20] mm时,细茎冰草单根极限延伸率较紫花苜蓿的差异性相对较小,细茎冰草单根极限延伸率为紫花苜蓿的 1.59倍,相应地,在根径级别为(0.30,0.40] mm时,细茎冰草单根极限延伸率与紫花苜蓿的差异性相对较大,即细茎冰草单根极限延伸率为紫花苜蓿的2.11倍。
通过上述对区内垂穗披碱草、细茎冰草及紫花苜蓿3种草本植物单根根径在(0.10,0.20]、(0.20,0.30]、(0.30,0.40]、(0.40,0.50]及(0.50,0.60] mm 5个根径级别条件下,单根抗拉力、单根抗拉强度和单根极限延伸率 3个单根抗拉力学特性指标对比分析可知,在 5个根径级别条件下,紫花苜蓿单根抗拉力和单根抗拉强度均表现出显著高于垂穗披碱草和细茎冰草的特性,且紫花苜蓿单根极限延伸率均表现出低于垂穗披碱草和细茎冰草的特性。由于本项研究所选取的 3种草本植物单根根径均小于1 mm,表现出主要以加筋作用方式增强边坡土体抗剪强度,即当边坡土体受到剪切力作用时,穿过土体剪切面的根系通过将自身所受拉应力转化为剪切面的法向应力和切向应力,从而起到增强边坡土体抗剪强度的作用[5,13],故在根面积比相同情况下,植物的单根抗拉力和单根抗拉强度愈大,则愈能起到增强边坡土体抗剪强度的作用[14]。相应地,植物单根极限延伸率愈小,当土体发生相对较小剪切位移时,单根即可将其自身的拉应力发挥出来,以起到有效增强土体抗剪强度的作用。由上述可知,在根面积比相同且根径为 0.10~0.60 mm条件下,区内 3种草本植物中紫花苜蓿根系增强边坡土体抗剪强度的作用较区内其他 2种草本植物相对较为显著,其次依次为细茎冰草和垂穗披碱草。
3.2 3种草本植物单根抗拉力学特性指标与根径的关系
3.2.1 单根抗拉力与根径关系
如图3所示为试验区3种草本植物单根抗拉力与根径之间的关系曲线。
由图3可知,区内3种草本植物单根抗拉力均表现出随着根径(根径范围为0.10~0.60 mm)的增加呈增大的变化规律,且单根抗拉力与根径间符合幂函数关系。

图3 试验区3种草本植物单根抗拉力与根径关系Fig.3 Relationship between single root tensile force and root diameter of three herbs in test area
垂穗披碱草、细茎冰草和紫花苜蓿 3种草本植物单根抗拉力与根径间的函数关系式分别为 y=17.06x1.25(R2=0.98,P<0.001)、y=20.73x1.40(R2=0.90,P<0.001)和y=21.54x1.27(R2=0.73,P<0.001)。此外,有关植物单根抗拉力与根径之间的关系,与本项研究结果相类似的相关研究主要表现为:陆桂红等[34]通过对寒芒、草地早熟禾等 6种草本植物根系进行室内单根拉伸试验,结果表明根系抗拉力与根径间呈幂函数关系,且抗拉力随根径的增加而增大;Ye等[20]通过对百喜草进行单根拉伸试验,亦得到了单根抗拉力随根径增加呈幂函数增长的变化关系;Lateh等[35]对乔木马占相思进行了单根拉伸试验,也得到了单根抗拉力随着根径增加呈幂函数增长的变化关系。
3.2.2 单根抗拉强度与根径关系
如图4所示为试验区3种草本植物单根抗拉强度与根径之间的关系曲线。由图4可知,本项研究所选取的3种草本植物单根抗拉强度随着根径(根径为0.10~0.60 mm)的增加呈幂函数降低的变化规律。垂穗披碱草、细茎冰草及紫花苜蓿3种草本植物单根抗拉强度与根径之间的函数关系式分别为y=22.09x-0.74(R2=0.94,P<0.001)、y=27.71x-0.60(R2=0.54,P<0.001)和y=25.07x-0.85(R2=0.58,P<0.001)。此外,有关植物单根抗拉强度与根径关系研究方面,赵丽兵等[19]通过室内单根拉伸试验,分析了紫花苜蓿和马唐2种草本植物单根抗拉强度及其与根径之间的关系,试验结果表明紫花苜蓿和马唐其单根抗拉强度随着根径增加均呈幂函数降低的变化关系;李光莹等[36]采用单根拉伸试验,亦得到了小嵩草、紫花针茅等6植物,其单根抗拉强度随着根径的增加呈幂函数降低变化关系的研究结论;Esmaiili等[37]通过单根拉伸试验得出欧洲云杉和欧洲白蜡树单根抗拉强度随着根径的增加亦呈幂函数降低的变化关系。上述研究结果均与本项研究所得到的结果相类似。
相应地,有关草本植物单根抗拉强度随着根径的增加而降低这一特性成因机理研究方面,赵丽兵等[19]的研究结果表明,紫花苜蓿和马唐 2种草本植物根纤维素含量随着根径增加呈线性降低的变化关系,而上述 2种草本植物单根抗拉强度随着根纤维素含量增加呈线性增大变化关系,故使得这 2种草本植物出现单根抗拉强度随着根径增加而降低的特性。上述研究结果对本项研究中,有关垂穗披碱草、细茎冰草及紫花苜蓿 3种草本植物单根抗拉强度随着根径增加而降低的试验结果,亦具有一定程度的参考价值。
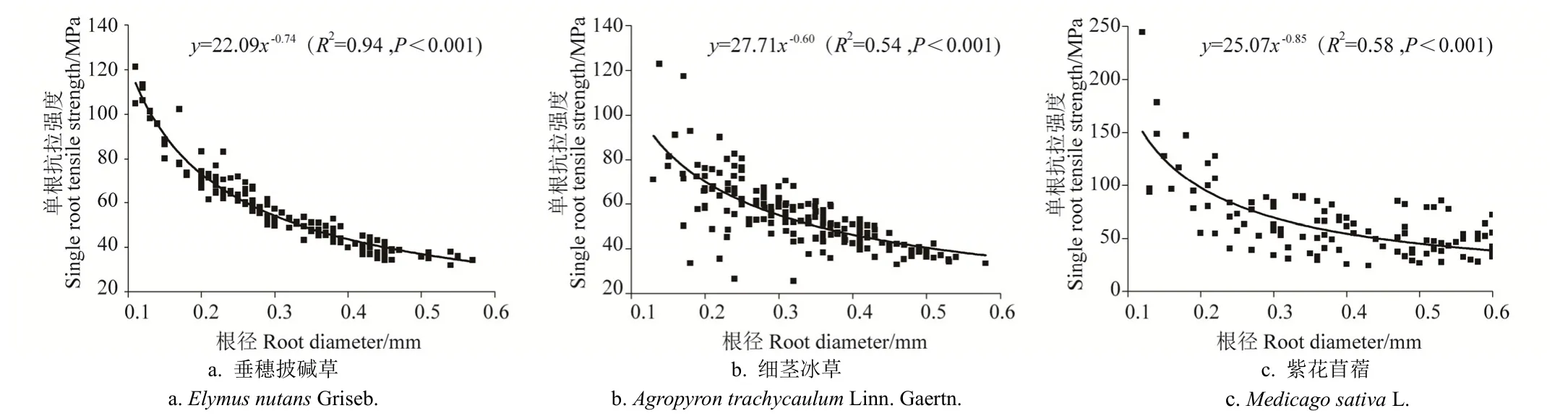
图4 试验区3种草本植物单根抗拉强度与根径关系Fig.4 Relationship between single root tensile strength and root diameter of three herbs in test area
3.2.3 单根极限延伸率与根径关系
如图5所示为试验区3种草本植物单根极限延伸率与根径之间的关系曲线。由图 5可知,垂穗披碱草、细茎冰草及紫花苜蓿 3种草本植物,其单根极限延伸率与根径(根径为0.10~0.60 mm)之间的关系未表现出相对显著性,即未出现决定系数R2大于0.5的函数关系(图5中所示为决定系数R2为最大值时所对应的拟合曲线及函数关系式)。
虽然区内 3种草本植物单根极限延伸率与根径之间关系的回归分析结果反映出两者间相关性相对较低,但由图5可知,区内3种草本植物单根极限延伸率,仍表现出随着根径的增加而逐渐增大的变化规律。
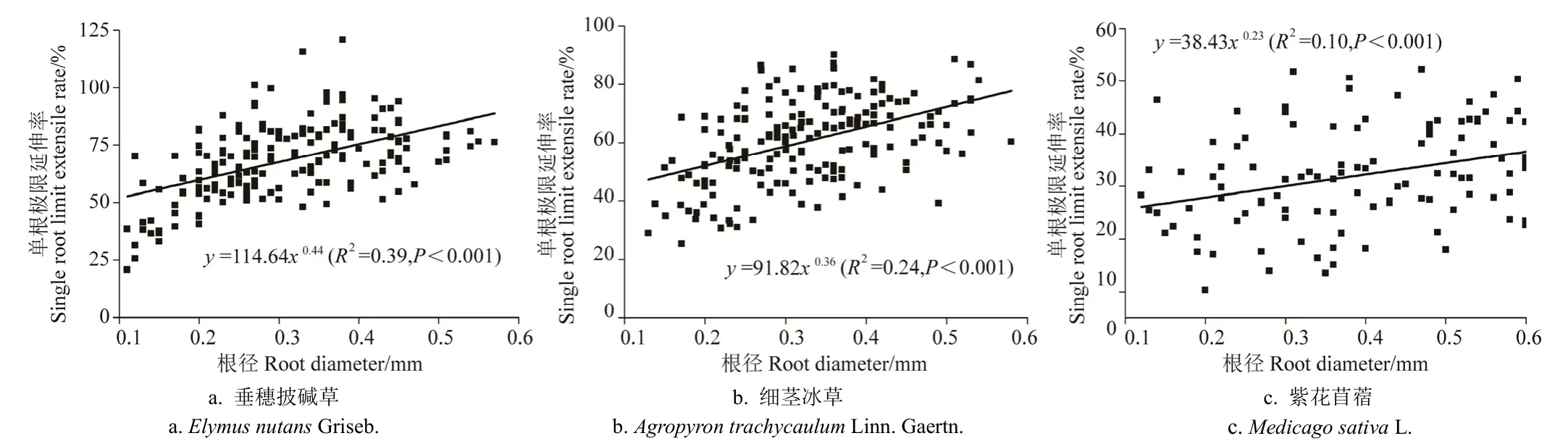
图5 试验区3种草本植物单根极限延伸率与根径关系Fig.5 Relationship between single root limit extensile rate and root diameter of three herbs in test area
3.3 3种草本植物单根拉伸应力-应变关系
本项研究通过对区内 3种草本植物根系进行单根拉伸试验,得到了垂穗披碱草、细茎冰草及紫花苜蓿 3种草本植物单根拉伸全过程应力-应变曲线,如图6所示。由图6可知,区内3种草本植物单根应力-应变关系曲线,存在2种类型的应力-应变关系。第一种类型即应力-应变关系曲线Ⅰ,该类型曲线为 3种草本植物普遍具有的单根拉伸应力-应变关系,其特征具体表现为:①单根受力的初期阶段,即表现在图6a、6b和6c中应力-应变关系曲线Ⅰ的O点至A点处,其应力-应变关系曲线基本呈直线型,反映出单根应力与应变之间呈线性变化关系,该阶段属于单根拉伸过程中的弹性变形阶段。该阶段单根变形可恢复,即卸去拉伸力作用后,单根形态可恢复至其在受拉变形前的初始状态,该阶段单根内部尚未产生残余变形和残余应力;②单根受力的中期阶段,即表现在图6a、6b和6c中应力-应变关系曲线Ⅰ的A点至C点处,单根的应力-应变曲线逐渐呈非线性,说明此阶段单根在拉应力作用下开始屈服,即单根进入弹塑性变形阶段;③单根受力的后期阶段,即表现在图 6a、6b和 6c中应力-应变关系曲线Ⅰ的C点至H点处,应力-应变曲线近似呈直线型,表现为应力与应变之间呈线性增长关系,在该阶段,单根的塑性变形增加了其对继续变形的阻力,故随着单根拉应力增大的同时,单根的屈服点位置亦表现出相应地提高的特性,即出现应变硬化现象;④当应变硬化现象发展至H点时,应力-应变曲线发生突变,即在该点处拉应力迅速下降至0,说明单根此时已被拉断,与此同时,应力-应变关系曲线在H点达峰值点,即H点所对应的极限拉应力即为单根抗拉强度[38]。在此需说明的是,在上述单根拉伸应力-应变关系曲线中,O点位置未选择在坐标原点,其原因在于单根拉伸试验过程中,由于单根其自身存在一定程度的自然弯曲现象,且当单根被夹具夹紧后亦会使单根出现一定的弯曲松弛现象,因此在如图6所示的应力-应变关系曲线中,自坐标原点至O点处的应力-应变关系并非代表单根实际的应力-应变变化关系,故本项研究选择 O点作为单根应力-应变关系曲线的起始点位置。
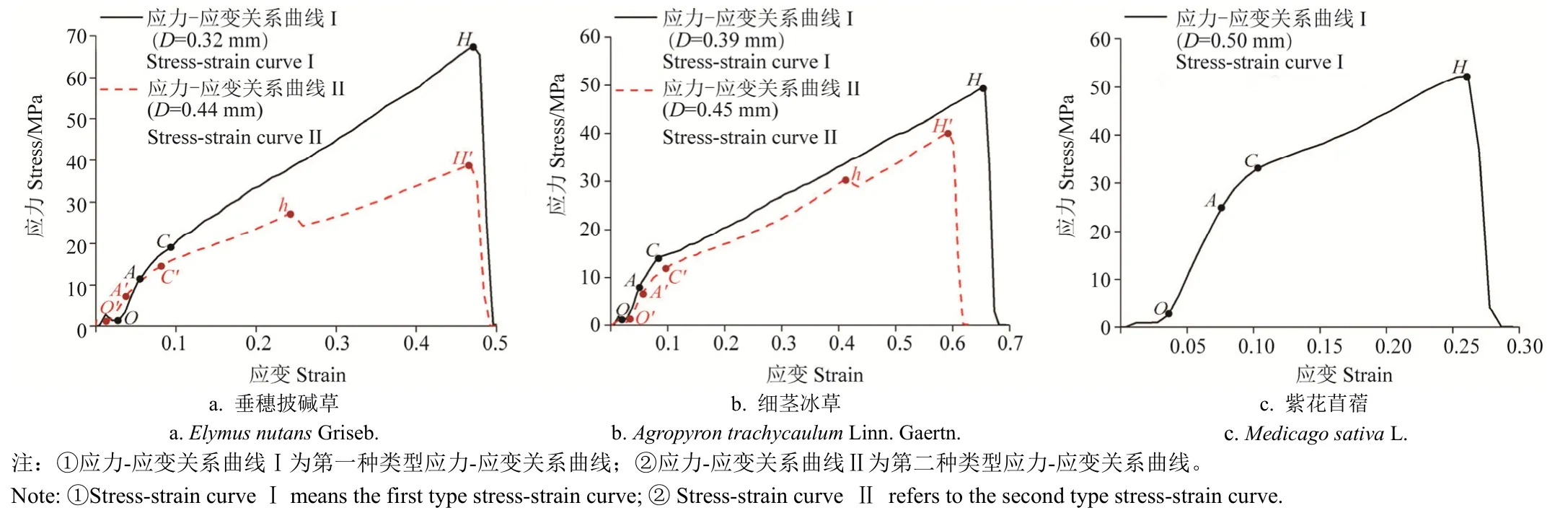
图6 试验区3种草本植物单根应力-应变关系曲线Fig.6 Stress-strain curves of three herbs in test area
区内3种草本植物单根拉伸过程中第二种应力-应变关系类型为应力-应变关系曲线Ⅱ,该类型曲线仅存在于垂穗披碱草和细茎冰草 2种草本植物单根的拉伸试验过程中。由图 6a、6b可知,应力-应变关系曲线Ⅱ与应力-应变关系曲线Ⅰ相比,二者表现出一定的相似性,即反映在应力-应变关系曲线Ⅱ中O'A'段、A'C'段及C'H'段,对应地与应力-应变关系曲线Ⅰ中的OA段、AC段及CH段具有相同的单根拉伸应力-应变关系阶段性变化特征。相应地,应力-应变关系曲线Ⅱ与应力-应变关系曲线Ⅰ的差异性则表现在:当单根应力-应变关系曲线Ⅱ处于应变硬化阶段(即图6a、6b中C'点至H'点位置)时,拉应力在未达到单根抗拉强度时(即应力-应变关系曲线未到达H'点位置时),即当应变为0.24(如图6a)和0.42(如图6b所示)时出现了拉应力峰值点 h,且其对应的拉应力分别为26.5和29.8 MPa,在峰值点h出现之后应力-应变关系曲线Ⅱ呈一定程度下降后,仍处于应变硬化阶段,直至单根应力-应变关系曲线到达H'点。通过对单根拉伸过程中单根表面形态和结构变化特征详细观察,发现峰值点 h出现的原因与垂穗披碱草和细茎冰草单根表皮先于表皮内部组织发生断裂现象关系密切。在单根拉伸过程中,当垂穗披碱草和细茎冰草单根表皮发生断裂后,其结果使得应力-应变关系曲线Ⅱ在C'H'段出现了峰值点h,此后其单根内部组织继续承受拉应力,直至单根应力-应变关系曲线到达极限拉应力点H'。因此,应力-应变关系曲线Ⅱ在整体上呈现出了双峰值的特征。值的一提的是,应力-应变关系曲线Ⅱ未出现在紫花苜蓿单根拉伸试验过程中,说明根径为0.10~0.60 mm的紫花苜蓿单根表皮与表皮内部组织的抗拉力学特性的差异性相对较为不显著,使得紫花苜蓿单根表皮与表皮内部组织的应力-应变过程,在单根拉伸试验过程中表现出具有相对较为显著的同步性。
4 结 论
1)西宁盆地黄土区内3种草本植物在根径为(0.10,0.20]、(0.20,0.30]、(0.30,0.40]、(0.40,0.50]及(0.50,0.60] mm 5个根径级别条件下,紫花苜蓿单根抗拉力和单根抗拉强度均表现出显著高于垂穗披碱草和细茎冰草的特性,且紫花苜蓿单根极限延伸率均表现出低于垂穗披碱草和细茎冰草的特性,体现出紫花苜蓿根系增强边坡土体抗剪强度的作用较细茎冰草和垂穗披碱草相对较为显著。
2)区内3种草本植物根径为0.10~0.60 mm范围内单根抗拉力随着根径的增加而增大,且二者间呈幂函数关系;单根抗拉强度随着根径的增加而减小,二者间亦呈幂函数关系;单根极限延伸率表现出具有随着根径增加而增大的变化趋势,但二者间并未表现出相对显著的函数关系。
3)区内3种草本植物在单根拉伸过程中表现出了2类应力-应变关系曲线,其中应力-应变关系曲线Ⅰ和应力-应变关系曲线Ⅱ均表现出了弹性变形、弹塑性变形、应变硬化及断裂破坏4个阶段;此外,应力-应变关系曲线Ⅱ在应变硬化阶段表现出双峰值现象;应力-应变关系曲线Ⅱ未出现在紫花苜蓿单根拉伸试验过程中,说明紫花苜蓿其单根表皮与表皮内部组织的应力-应变过程,在单根拉伸试验过程中表现出具有相对较为显著的同步性。
该项研究成果对于研究区坡面水土流失、浅层滑坡等地质灾害现象的生态防护工作,具有理论研究价值和实际指导意义。
