干旱胁迫条件下小麦各器官可溶性碳水化合物积累转运相关性状遗传分析
2018-08-08张家乐栗孟飞杨德龙陈菁菁程宏波柴守玺
张家乐,栗孟飞,杨德龙,刘 媛,陈菁菁,程宏波,常 磊,柴守玺
(1. 甘肃省干旱生境作物学重点实验室/甘肃农业大学生命科学技术学院,甘肃 兰州 730070; 2. 甘肃农业大学农学院, 甘肃 兰州 730070)
干旱已成为限制小麦产量形成的重要非生物胁迫因子,每年世界各地因不同程度的干旱所导致的小麦减产约在30%左右,其危害相当于其他自然灾害的总和[1-3]。因此,只有提高小麦自身的水分利用效率才有可能取得小麦抗旱上的新突破[4]。
小麦产量的形成是源、库、流互作的结果,要保证小麦的稳产高产,不仅要有较强的光合物质生产能力,又要求营养器官中的光合产物能更多地向籽粒库中转运[5]。小麦营养器官中暂贮性可溶性碳水化合物(Water soluble carbohydrates,WSC)不仅是籽粒产量形成的重要碳源,还可以缓解逆境胁迫对小麦的伤害。尤其是在干旱胁迫条件下,小麦光合作用产物无法满足冠层呼吸消耗和保持籽粒正常灌浆需要时,花前贮存于茎秆中的WSC积累与转运对产量的形成具有重要作用[6-9],且对籽粒干物质积累的贡献率可达10%~70%[10-13]。然而,小麦营养器官WSC积累与转运不仅受基因型的控制,而且与环境条件有密切关系[14],进一步研究表明,小麦不同器官对小麦WSC转运及其对籽粒粒重的贡献也有显著影响[7,15]。由此表明,干旱调控小麦营养器官WSC积累转运与产量形成的关系非常复杂。因此,开展不同水分环境下不同基因型小麦各营养器官WSC积累转运与籽粒灌浆和产量形成的遗传关系研究,可为小麦抗旱种质资源的筛选和抗旱遗传育种提供重要的理论依据。
1 材料与方法
1.1 供试材料
以不同生态区196份小麦抗旱种质组成的自然群体为供试材料(表1)。
1.2 试验设计
试验于2014年10月至2015年6月在甘肃省兰州市榆中县金家营小麦试验点(35°51′N,104°07′E,平均海拔1 900 m,平均气温6.7℃,年降雨量450 mm,年蒸发量1 450 mm,无霜期140 d)进行。小麦播前基施K2O 60 kg·hm-2、N 180 kg· hm-2、P2O5150 kg·hm-2,在整个生育期内均不再施肥。水分管理分为干旱胁迫(Drought stress,DS)和正常灌溉(Well-watered,WW)两种处理。播前试验地均统一灌底墒水900 m3·hm-2。正常灌溉处理条件下,在小麦拔节期、抽穗期和开花期补充灌水,每次灌水量为750 m3·hm-2;干旱胁迫处理条件下,仅在小麦拔节期灌水750 m3·hm-2,其后完全依靠自然降水(2014年10月-2015年6月小麦全生育期降水量为130 mm)。在两种水分处理的试验区之间种植18 m宽的小麦隔离带,隔离带水分处理同干旱胁迫处理。整个试验随机区组设计,3次重复,条播,行长1 m,行距0. 2 m,每行点播60粒,12行区,即每个小区2.4 m2。
1.3 目标性状测定与统计分析
随机选择小麦自然群体内各品种开花期长势一致主茎做标记,每个处理分别在小麦开花初期、灌浆中期(花后14 d)和成熟期剪取主茎15株,取旗叶(FL)、穗下节(包括茎鞘,PedI)、倒二叶(SL)、倒二节(包括茎鞘PenI)、倒三叶(TL)和倒三节(包括茎鞘,ThiI),在105℃杀青30 min,80℃烘干至恒重,并称取主穗粒重(SGW)。采用蒽酮比色法[16]测定不同处理的WSC含量,各器官开花期、灌浆期和成熟期的WSC含量分别记为WSCf,WSCg和WSCm,所有测定均3次重复。
WSC花前转运率(RRpr)=(WSCf绝对含量-WSCm绝对含量)/WSCf绝对含量×100%;WSC花后转运率(RRps)= (WSCg绝对含量-WSCf绝对含量)/WSCg绝对含量×100%;WSC花前贡献率(CRpr)= (WSCf绝对含量-WSCm绝对含量)/(1000×SGW)×100%;WSC花后贡献率(CRps)= (WSCg绝对含量-WSCf绝对含量)/(1000×SGW)×100%;WSC绝对含量=WSC浓度×干物质量。
1.4 数据处理
采用DPS V7.0.5 统计软件进行不同处理间各目标性状方差分析(ANVOA)、多重比较、相关分析和聚类分析。聚类分析采用系统聚类中的类平均法(UPGMA),聚类距离为欧式距离。
性状表型多样性指数采用Shannon和Weaver[17]的方法计算。多样性指数划级方法:先计算参试材料总体平均数(X)和标准差(d),然后划分为10个等级,从第1级[Xi<(X-2d)]到第10级[Xi>(X+2d)],每0.5d为1个等级,每一级的相对频率用于计算多样性指数。多样性指数的计算公式为:H=-∑PilnPi,式中,Pi为某性状第i级别中材料份数占总份数的百分比。

表1 供试小麦材料Table 1 Wheat accessions
2 结果与分析
2.1 小麦自然群体各性状表型多因素方差分析
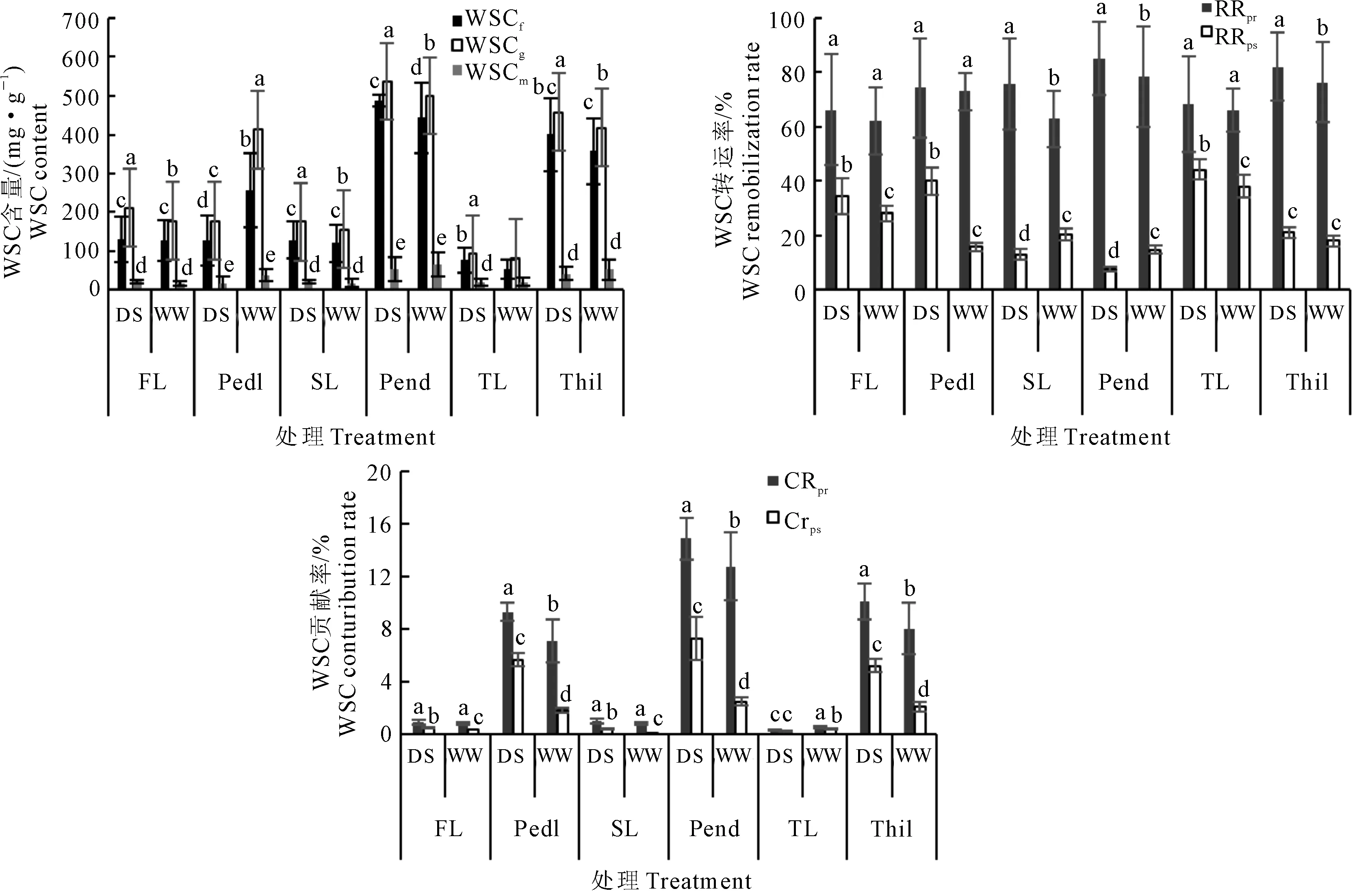
小麦自然群体WSC含量受到水分、发育阶段、器官和基因型及其各因子之间互作效应的显著(P<0.05)或极显著(P<0.01)的影响(表2)。从F值看出,发育阶段(F=32 775.74**)和器官(F=11 223.18**)是影响该群体WSC含量表型变异的主要因素;器官(F=390.15**~1 482.96**)和水分(F=72.16**~274.61**)是影响WSC花前、花后转运率及其对籽粒贡献率表型变异的主要因子;而其他因子及其互作对目标性状表型变异的相对贡献率较小。WSC各目标性状遗传力较低,在0.24~0.53。
2.2 不同水分条件下小麦自然群体不同器官相关性状表型分析
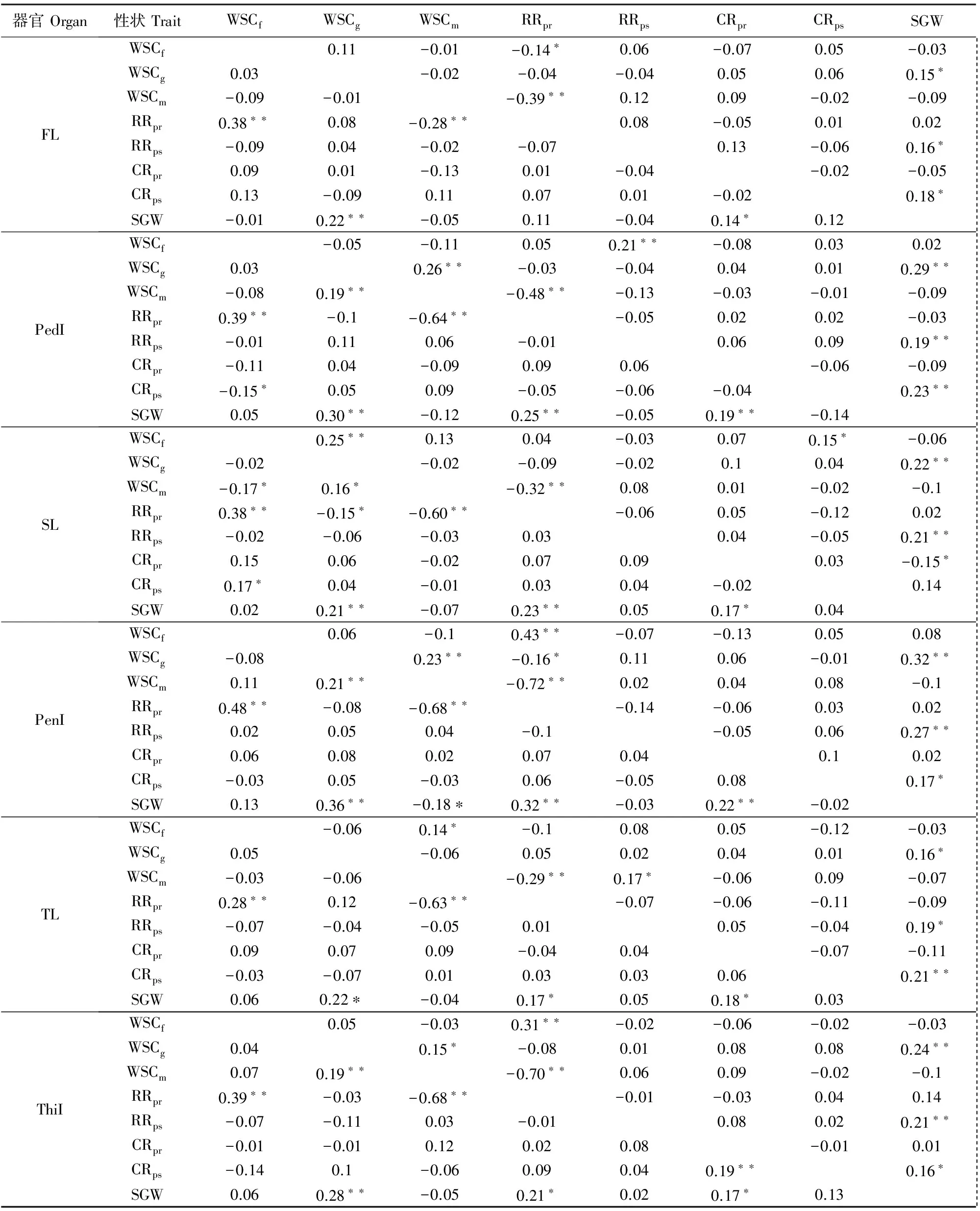
在两种水分条件下,小麦自然群体不同发育时期WSC含量,各器官花前、花后WSC转运率及其对籽粒贡献率表型值均差异显著(图1)。各器官WSC含量均呈现出先升高后降低的趋势,即WSCm 从图2可以看出,小麦自然群体WSC相关性状表型多样性丰富,多样性指数为0.61~0.93(DS)和0.62~0.88(WW)。其中,干旱胁迫条件下的WSCf、WSCg、WSCm、RRpr和RRps多样性指数显著高于正常灌溉,但RRpr和CRps相反。WSC含量、WSC花前和花后转运率及其对籽粒贡献率随着主茎节位的下降和生育期的延后呈降低趋势。表明干旱胁迫显著提高了各器官WSC含量多样性指数。 表2 小麦自然群体各性状表型多因素方差分析(F值)Table 2 Multi-factor variance analysis (F value) of target traits in the wheat natural population 注:WSC表示WSC含量;RRpr和RRps分别表示花前WSC转运率和花后WSC转运率;CRpr和CRps分别表示花前WSC贡献率和花后WSC贡献率。*和**分别表示多因素方差分析中显著性水平为P<0.05和P< 0.01。 Note: WSC mean WSC content; RRprand RRpsmean WSC remobilization rate at the pre-anthesis and WSC remobilization rate at the post anthesis, respectively;CRprand CRpsmean WSC contribution rate at the pre-anthesis and WSC contribution rate at the post anthesis , respectively. * and ** mean different significence levels atP<0.05 andP<0.01 by the multi-factor analysis of variation. 注:WSCf:初花期WSC含量;WSCg:花后14d WSC含量;WSCm:成熟期WSC含量;FL:旗叶;PedI:穗下节;SL:倒二叶;PenI:倒二节;TL:倒三叶;ThiI:倒三节;DS:干旱胁迫;WW:正常灌溉。下同。不同小写字母表示同一器官不同处理间差异显著(P<0.05)。Note: WSCf:WSC content at the early anthesis stage; WSCg:WSC content 14 days after anthesis;WSCm:WSC content at the mature stage; FL:flag leaf; PedI:peduncle internode; SL: top second leaf; Penl:the second internode; TL:top third leaf; ThiI:the third internode from top. DS:drought stress; WW:well-watered. The same below. Different small letters mean significant difference between different treatment in the same organ(P<0.05).图1 不同水分条件下小麦自然群体WSC积累转运表型Fig.1 WSC accumulation and remobilization in wheat natural population under different water treatment 图2 不同水分条件下小麦自然群体各器官WSC积累转运相关性状表型多样性指数Fig.2 Phenotype diversity index of associated traits related to WSC accumulation and remobilization of different organ in wheat natural population under different water treatment 从表3得出,两种水分条件下的WSCg与SGW间均呈显著或极显著正相关,相关系数(r)0.21**~0.36**(DS)和0.15*~0.32**(WW),倒二节的WSCg与SGW间相关系数达到最高0.36**(DS)和0.32**(WW)。在干旱胁迫条件下,WSCf和RRpr呈极显著正相关(r=0.28**~0.48**),但WSCm相反(r=-0.28**~-0.68**);且WSCg、RRpr、CRpr与SGW均呈不同程度正相关,相关系数(r)为0.11 ~ 0.36**。在正常灌溉条件下,RRps、CRps和SGW呈不同程度正相关,相关系数在0.14~0.27**;而WSCm与RRpr呈极显著的负相关,相关系数为-0.29**~-0.72**。在不同器官中,各因子间呈显著或极显著关系,茎鞘均高于叶片,且在倒二节达到最大值。 在干旱条件下对该自然群体灌浆期196份抗旱种质材料WSC倒二节相关性状表型值进行聚类分析(图略)发现,该群体WSC倒二节含量被分为三类。第Ⅰ类,包括76个株系,各株系倒二节WSC含量较高,其中包括43份甘肃品种、14份山西品种、7份河北品种、3份陕西品种、5份山东品种、1份天津品种和3份北京品种;第Ⅱ类,包括96个株系,其中54份甘肃品种、17份山西品种、4份河北品种、2份山东品种、1份天津品种、6份北京品种和12份美国品种;第Ⅲ类,各株系倒二节WSC含量较低,包括24个株系,其中16份甘肃品种、2份山西品种、1份河南品种、3份北京品种和2份美国品种。该群体在干旱胁迫下WSC转运率表型值被分为三类。第Ⅰ类,包括104个株系,各株系倒二节WSC转运率较高,包含61份甘肃品种、19份山西品种、3份河北品种、2份陕西品种、1份河南品种、2份山东品种、6份北京品种和10份美国品种;第Ⅱ类包括82个株系,其中46份甘肃品种、11份山西品种、7份河北品种、3份陕西品种、5份山东品种、6份北京品种和4份美国品种;第Ⅲ类,包括10个株系,各株系倒二节WSC转运率较低,其中4份甘肃品种、2份山西品种、2份陕西品和2份天津品种。在干旱条件下该群体倒二节WSC贡献率被分为三类。第Ⅰ类,倒二节WSC贡献率较高,包括71个株系,其中36份甘肃品种、13份山西品种、6份河北品种、1份河南品种、2份山东品种、1份天津品种、5份北京品种和7份美国品种;第Ⅱ类,包括77个株系,其中44份甘肃品种、12份山西品种、4份陕西品种、5份山东品种、1份天津品种、4份北京品种和3份美国品种;第Ⅲ类,包括48个株系,各株系倒二节WSC贡献率较低,其中包含31份甘肃品种、7份山西品种、2份河北品种、1份陕西品种、3份北京品种和4份美国品种。本研究中的196份不同地理来源的小麦种质,因其遗传背景的复杂性和多样性,以及各品种株系对干旱胁迫环境适应能力的差异,导致其在灌浆期可溶性碳水化合物在倒二节中的积累和转运具有显著差异,而倒二节WSC的含量及其转运能力与产量在一定程度上呈正相关,从而使小麦种质表现出不同的抗旱能力,最终表现为产量的差异。此外,通过对该自然群体灌浆期倒二节WSC含量、转运率及其贡献率的表型值聚类分析得到,具有较高WSC含量、转运率和贡献率的品种株系15个,分别为21-30、运旱22-33、冬03-07、鲁麦15、石麦13、B61-2-3-2-1、Dd52-17-3-2、临旱51241、陇鉴127、京双2号、B160-2-2-1-1、A80-3-2-2-1、清农3、石家庄8号、临丰3号。 表3 干旱胁迫和灌溉条件下小麦自然群体不同器官WSC积累转运与穗粒重的相关系数Table 3 Correlation coefficients between WSC accumulation and remobilization of different organ and grain weight per spike in wheat natural population under drought stress and well-watered conditions 注:表中左和右两个三角区域分别表示干旱胁迫和灌溉条件下性状的相关系数。 Note: Figures at the lower left segment are the correlation coefficients under rain-fed conditions (DS);those in the upper right under well-watered conditions (WW). 小麦的抗旱性是复杂的生物学性状,往往通过一系列生理反应来应对不同程度的干旱胁迫[18]。小麦叶片(尤其是旗叶)作为光合作用的主要器官,直接决定着植株WSC的积累量,在干旱胁迫条件下,小麦叶片卷曲,气孔关闭,在灌浆期小麦叶片光合产物无法满足冠层呼吸消耗时,小麦的籽粒灌浆和最终产量的形成更加依赖于茎秆WSC的积累和转运[6,19]。研究表明,适度的干旱胁迫,可以促进小麦暂贮性WSC含量的增加,并且调控其向籽粒的转运,同时提高小麦水分利用效率,从而提高小麦御旱能力,保证小麦籽粒的产量[20-24]。在本研究中,小麦自然群体灌浆期不同发育阶段WSC含量,花前、花后转运率及其对籽粒的贡献率普遍表现为干旱胁迫高于正常灌溉,WSCg高于WSCf和WSCm,倒二节和倒三节WSC含量显著高于其他器官并在倒二节中达到最大值,与赵万春等[25]研究结果一致,并且小麦籽粒灌浆更多依赖于茎鞘中贮藏的WSC。由此说明,小麦自然群体WSC积累转运具有明显的水分环境、器官和发育阶段效应,不同性状对以上诸因子响应不同。小麦各器官WSC含量受发育时期影响较大,且各器官WSC含量差异显著,小麦WSC转运率受水分和器官的共同作用,而WSC转运对籽粒的贡献具有显著的水分和器官的主导效应,表现出小麦WSC积累转运的复杂数量性状的特点及其在小麦抗旱种质研究中的重要作用。 小麦作为C3植物,要求具有较强的光合能力和有效的转运与再分配能力,贮存更多的光合产物,可促进WSC向籽粒的运输,为籽粒产量的形成提高充足的底物条件[19]。前人利用小麦DH群体和RIL群体研究发现,小麦茎秆WSC含量和WSC积累转运量与产量和穗粒重呈显著或极显著正相关,且干旱胁迫条件下相关系数显著高于正常灌溉[26-28]。在本研究中,两种水分条件下,小麦自然群体各器官WSCg与SGW均呈显著或极显著正相关,且在倒二节中WSCg与SGW的相关系数达到最大值。在干旱胁迫条件下,WSCg、RRpr、CRpr与SGW均呈不同程度正相关,而在正常灌溉条件下,RRps、CRps和SGW呈不同程度正相关,并且相关程度干旱胁迫普遍高于正常灌溉,茎鞘显著高于叶片,倒二节相关系数最高。表明灌浆中期小麦暂贮性WSC对小麦WSC积累转运及其对籽粒灌浆的贡献具有重要作用,尤其是在干旱胁迫条件下,小麦可能通过自身的生理调节作用,动员茎鞘(尤其是倒二节)中暂贮性WSC向籽粒转运,从而补偿籽粒产量损失,缓解干旱胁迫引起的环境压力。 本研究发现,小麦自然群体WSC含量,花前、花后转运率及其对籽粒的贡献率表型变异丰富,在不同水分处理下,不同发育阶段WSC含量以及不同器官中的WSC花前、花后转运率和贡献率多样性指数较高。杨德龙等[28-29]利用DH群体和RIL群体对小麦灌浆期WSC积累转运相关性状进行遗传分析,发现群体内株系各性状表型变异广泛,介于双亲之间,且表现出超亲分离,遗传力较低,多样性指数高。说明小麦的遗传背景、水分、发育阶段和器官等差异,导致了小麦WSC积累转运丰富的表型多样性。因此,加速开展复杂遗传背景的小麦种质在不同水分环境下的WSC积累转运的数量遗传研究,将有利于发掘WSC积累转运丰富的抗旱遗传信息[30]。在本研究中,干旱胁迫显著地提高了小麦自然群体WSC积累转运表型的变异,丰富了表型多样性。这可能是水分胁迫诱导的一些在非水分胁迫条件下沉默的基因表达,以应对干旱胁迫环境对植株的伤害,而这也可能是导致这些性状表型值在干旱胁迫条件下普遍增高的重要遗传基础。2.3 小麦自然群体WSC相关性状表型遗传多样性分析


2.4 小麦自然群体各目标性状之间的相关性分析
2.5 小麦自然群体表型聚类分析
3 讨 论
3.1 小麦自然群体WSC积累转运与抗旱性
3.2 小麦自然群体WSC积累转运与穗粒重的关系
3.3 小麦自然群体WSC积累转运的遗传特性
