曹妃甸近岸海域大、中型浮游动物优势种空间生态位研究
2018-08-04梁淼姜倩孙丽艳李德鹏陈兆林崔雷路波
梁淼,姜倩,孙丽艳,李德鹏,陈兆林,崔雷,路波
国家海洋环境监测中心/国家海洋局近岸海域生态环境重点实验室,辽宁 大连 116023
生态位表征物种对每个生态位空间点的响应及其在每个生态位空间点中的效应(彭文俊等,2016),体现了群落中某一物种与其他物种在时间、空间及其功能上的相对位置(Whittaker et al.,1973)。生态位研究己在群落结构功能、群落内物种间关系、生物多样性、群落动态演替和种群进化等方面占有重要的地位(张金屯,2004),并且运用生态位宽度和生态位重叠这两个指标进行相应的估算和分析(Grönhaug et al.,1989)。目前,国内关于生态位的研究主要集中在陆地植物(陈丝露等,2018;陈玉凯等,2014;汤景明等,2012)和陆生动物群落(张晶等,2018;苏军虎等,2017;陈顺德等,2016),只有少量涉及海洋生物,而后者该方面的研究主要集中在潮间带(焦海峰等,2011)、红树林(黄相相等,2017)及河口(张敬怀,2014)等区域的大型底栖生物、湖泊的浮游植物(郭坤等,2016)及游泳动物(邓小艳等,2018;李超男等,2017),涉及海洋浮游动物生态位的研究鲜有报道,仅在辽河口海域(张皓等,2016)、北部湾海域(郑挺等,2014)及浙江三门湾(徐晓群等,2013)等局部海域开展研究。
曹妃甸海域位于渤海湾东北部,属于半封闭内海,海水的交换能力较弱(索安宁等,2012)。近年来,该区域内填海造陆、港口开发、工业区建设和矿产开采等一系列开发活动的进行,给海洋生态环境带来了巨大压力,生物资源和环境质量也面临着前所未有的考验。浮游动物在海洋生态系统中起着承上启下的传递作用,对外界环境变化非常敏感,其空间生态位分布在一定程度上能间接反映出海洋生态环境变化现状(张皓等,2016)。鉴于此,本研究从优势种(类)群空间生态位角度出发,对曹妃甸近岸海域的浮游动物群落结构进行研究,以夏、秋两季优势种的生态位宽度和生态位重叠指数为基础,对浮游动物优势种(类)群的空间分布及环境因子的变化进行分析,探讨优势种在不同资源位点的功能地位和适应性,为该海域浮游动物群落结构特征研究和海洋生态环境保护、修复提供基础资料。
1 材料与方法
1.1 采样点设置
曹妃甸海域位于渤海湾湾口北侧,潮汐主要受西支渤海潮波系统控制,为不规则半日潮混合潮,一天出现两次高潮和两次低潮,平均潮差由东向西逐渐增大。潮流基本属于往复流,即涨潮西流,落潮东流,且涨潮流速大于落潮流速。波浪以风浪为主,常浪向为偏S向,强浪向偏E~ENE向,各方向风浪频率的季节变化比较明显。根据曹妃甸海域的地形地貌及海洋水文特征,本研究在该海域由西到东布设了14个调查站位(图1和图2),于2014年6月(夏季)和9月(秋季)对其浮游动物和海水理化因子进行了综合调查研究。
1.2 样品的采集和处理
1.2.1 浮游动物样品

图1 曹妃甸海域地理位置图Fig. 1 The geographical location map of the coastal area of Caofeidian
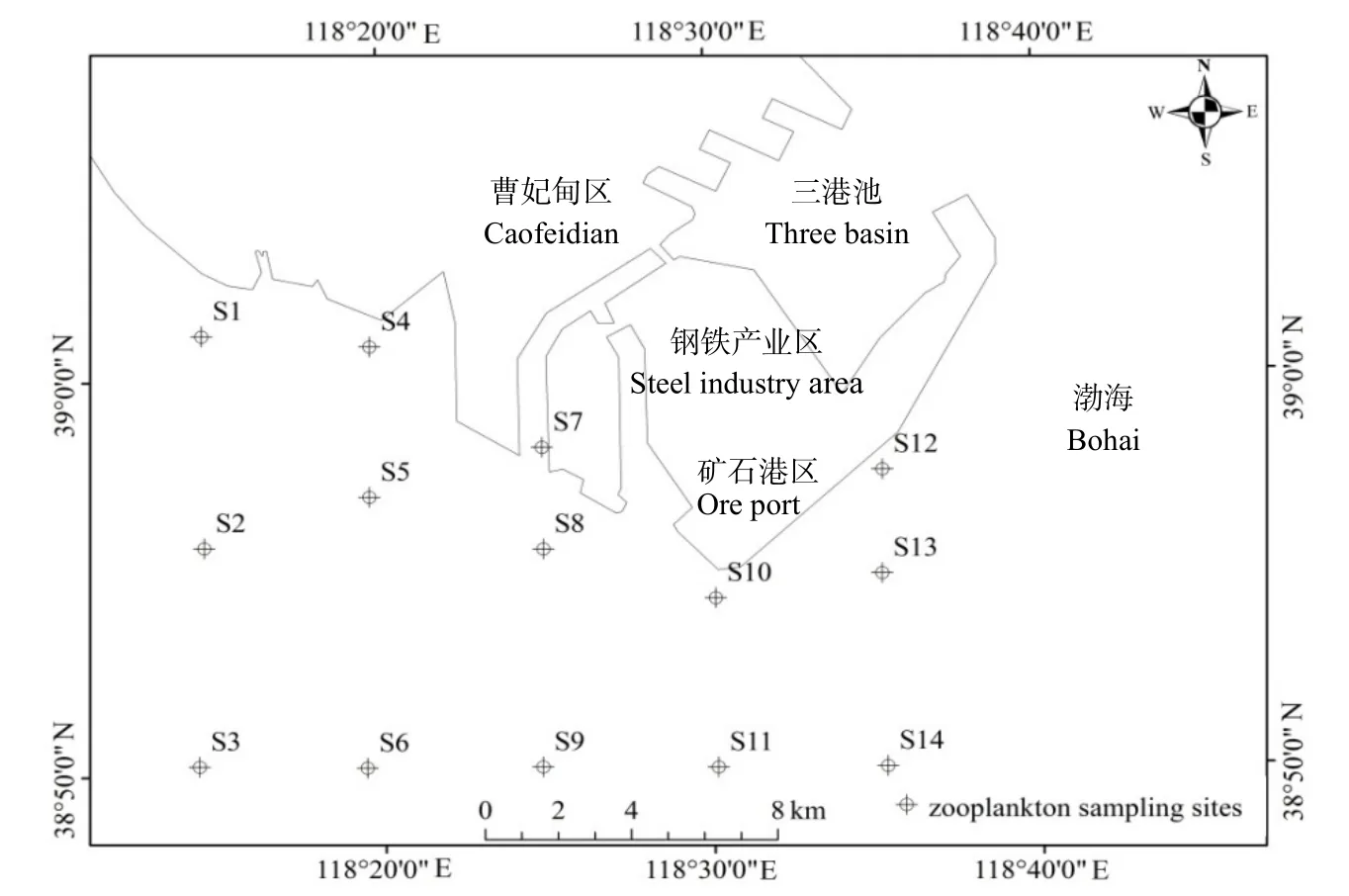
图2 曹妃甸近岸海域采样站位示意图Fig. 2 The diagrammatic sketch of sampling sites in the coastal area of Caofeidian
根据调查站位的水深(均小于30 m),使用浅水Ⅰ型浮游生物网(孔径0.505 mm,口径50 cm,网长145 cm)采集浮游动物,在每个采样站位,采用自底层至表层垂直拖网的方式对大、中型浮游动物进行定量采集。网口入水后,以不超过1 m∙s-1的速度下沉,直至网具到达距海底2 m后起网,以0.5 m∙s-1左右的速度起网,网具升出水面后,反复冲洗网衣外表面,以保证样品收集完整;开启网底管收集样品,并用体积分数为 5%的甲醛固定与保存。带回实验室,样品静置48 h后,虹吸上清液,在显微镜下进行浮游动物种类鉴定并计数。其中,计数法采用浓缩计数,种类鉴定参考形态学及分类系统等(郑重等,1984;刘瑞玉,2008;黄宗国,2008)。所有操作均按照《海洋调查规范》(GB/T 12763.6—2007)执行(中华人民共和国国家质量监督检验检疫总局,2007)。
1.2.2 水质指标
在每个调查站位,同步进行水样采集。使用多参数水质分析仪(HQ40D,Hach,Loveland,USA)对 pH、溶解氧(DO)、浊度(NTU)和水温(t)进行现场测定;盐度(S)、悬浮物(SS)、化学需氧量(COD)、无机氮(DIN)(氨、亚硝酸盐、硝酸盐)、活性磷酸盐(DIP)、叶绿素a(Chla)的分析测定于实验室内完成。水样采集和理化因子分析均参照《海洋调查规范》(GB/T12763.4—2007)(中华人民共和国国家质量监督检验检疫总局,2007)和《海洋监测规范》(GB17378.4—2007)(国家海洋局,2007)进行。
1.3 数据分析
1.3.1 优势度分析
优势度(Y)计算公式(Lampitt et al.,1993):

式中,N为该海域出现所有浮游动物种类的总个体数;ni为第i种的个体数;fi为第i种在各站位出现的频率。当Y>0.02时,即为该海域浮游动物优势种。
优势种更替率(R)(郑白雯等,2014)分析优势种的季节演替,其计算公式:
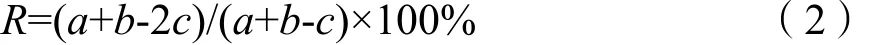
式中,a、b为相邻两季优势种数;c为相邻两季共同优势种的种类数。
1.3.2 生态位宽度
本研究采用的Levins(1968)生态位宽度公式进行计算比较:

式中,Bi为物种i的生态位宽度,取值范围为[0, 1];Pij为物种i在第j个资源状态下的个体数占该种所有个体数的比例;r为资源总数,本研究为采样站位数。
1.3.3 生态位重叠指数
本研究采用Pianka生态位重叠指数(Pianka,1973)对种群间的生态位重叠程度进行计算,其公式为:
式中,Oik为生态位重叠指数;Pij和 Pkj为种 i和k在资源j上的优势度;r为资源总数,本研究中r为采样站位数。
1.3.4 数据统计分析
利用 CANOCO 4.5统计软件对曹妃甸海域浮游动物优势种与环境因子进行多元统计分析。在进行统计分析前,对浮游动物优势种密度和环境因子(除 pH)的数据矩阵进行 log(x+1)转换,以使它们的分布更接近正态分布。在对数据进行空间排序之前,先对曹妃甸海域夏、秋季浮游动物优势种密度数据进行去趋势对应分析(DCA),结果表明,夏季排序轴最大梯度长度为1.401,秋季为0.689,均小于 3,表明浮游动物优势种群落对生态梯度的响应是线性的,因此选用冗余分析(RDA)线性模型进行排序分析,再对排序结果进行999次蒙特卡罗(Monte Carlo)置换检验,分析环境因子对浮游动物优势种分布的影响是否具有统计学意义(傅明珠等,2014)。
利用 SPSS 13.0对变量进行方差齐次性检验(Levene检验)后,再对不同季节的理化环境因子进行两独立样本t检验。
2 结果与分析
2.1 浮游动物优势种(类)组成、季节演替及空间分布特征
曹妃甸近岸海域2014年夏季、秋季共鉴定出3大类9种浮游动物优势种和4种浮游幼虫,其中夏季8种,秋季12种(表1)。夏、秋季共同出现的优势种有 6种,分别为小拟哲水蚤(Paracalanus parvus)、双毛纺锤水蚤(Acartia bifilosa)、拟长腹剑水蚤(Oithona similes)、多毛类幼虫(Polychaeta larva)、蔓足类无节幼虫(Cirripedia larva)和强壮箭虫(Sagitta crassa),优势种更替率达 57.14%,可见夏、秋季浮游动物优势种季节性更替较大。另外,克氏纺锤水蚤(Acartia clausi)为夏季第一优势种,优势度为0.246,出现频率为92.86%,对浮游动物总丰度的贡献率达26.46%;而其未在秋季出现。秋季的主要优势种为小拟哲水蚤和双毛纺锤水蚤,出现频率均为 100%,二者对浮游动物总丰度的贡献率达到31.26%,这两种优势种在夏季分别为第二、第三优势种,出现频率分别为 92.86%和64.29%。

表1 曹妃甸近岸海域浮游动物优势种统计情况Table 1 The zooplankton dominant species in the coastal area of Caofeidian
夏、秋季调查海域主要优势种的空间分布特征如图3所示。由图3可知,夏季,克氏纺锤水蚤和小拟哲水蚤的空间分布具有明显差异,克氏纺锤水蚤的高值区主要分布于调查海域东侧区,而小拟哲水蚤的高值区主要分布于调查海域西侧近岸海区。秋季,小拟哲水蚤和双毛纺锤水蚤分布区域较一致,高值区均分布在研究海域西侧近岸海域。曹妃甸海域的主要优势种的空间变化特征与整个渤海的分布特征是一致的,全年大部分时间,小拟哲水蚤和双毛纺锤水蚤的高生物密度区主要集中在近岸水域。
2.2 浮游动物优势种群的生态位宽度
利用Levins公式计算了夏、秋两季的浮游动物优势种生态位宽度,计算结果分别见表2和表3。根据生态位宽度值,将曹妃甸近岸海域浮游动物优势种群分为3大类:第一类为广生态位种类,其生态位宽度值Bi≥0.70;第二类为中生态位种类,其生态位宽度值0.50<Bi<0.70;第三类为窄生态位种类,其生态位宽度值为Bi≤0.50。
夏季,浮游动物群落中生态位宽度值变化范围为 0.28~0.89(表 2)。其中,广生态位种类包括强壮箭虫(0.89)和克氏纺锤水蚤(0.71)共 2种;中生态位种类包括小拟哲水蚤(0.67)、多毛类幼虫(0.59)、双毛纺锤水蚤(0.53)和拟长腹剑水蚤(0.52)共4种;窄生态位种类包括无节幼虫(0.39)和蔓足类无节幼虫(0.28)共2种。
秋季,浮游动物群落中生态位宽度值变化范围为 0.44~0.93(表 3)。其中,广生态位种类包括小拟哲水蚤(0.93)、异体住囊虫(0.91)、近缘大眼剑水蚤(0.87)、双毛纺锤水蚤(0.87)、蔓足类无节幼虫(0.83)、多毛类幼虫(0.82)、真刺唇角水蚤(0.81)和强壮箭虫(0.72)共 8种;中生态位种类包括腹足类幼虫(0.63)和太平洋纺锤水蚤(0.52)共2种;窄生态位种类包括中华哲水蚤(0.49)和拟长腹剑水蚤(0.44)共2种。

表2 夏季浮游动物优势种类群间的生态位宽度和生态位重叠指数统计Table 2 Niche breath and niche overlap of the dominant zooplankton species in the coastal area of Caofeidian in summer 2014

图3 曹妃甸近岸海域浮游动物主要优势种分布特征Fig. 3 Distribution of major dominant zooplankton species in the coastal area of Caofeidian
2.3 浮游动物优势种群的生态位重叠
曹妃甸近岸海域夏、秋季主要优势种类群生态位重叠指数的统计结果分别见表2和表3。由表2和表3可知,夏季,浮游动物优势种之间的生态位重叠指数范围为0.24~0.82,秋季生态位重叠指数范围为0.31~0.95,总体上,秋季各优势种之间的生态位重叠指数高于夏季。
夏季,广生态位种类强壮箭虫与克氏纺锤水蚤、双毛纺锤水蚤、拟长腹剑水蚤、小拟哲水蚤和多毛类幼虫的生态位重叠程度均较高,生态位重叠指数均在0.7以上,其中强壮箭虫与多毛类幼虫生态位重叠指数最高,为0.82;中生态位种类多毛幼虫与双毛纺锤水蚤、拟长腹剑水蚤和无节幼虫之间生态位重叠指数也在0.7以上。
秋季,生态位宽度最大的小拟哲水蚤与近缘大眼剑水蚤、双毛纺锤水蚤、真刺唇角水蚤、蔓足类无节幼虫、多毛类幼虫、异体住囊虫、强壮箭虫等有着很高的重叠度,分别为0.95、0.95、0.88、0.89、0.91、0.90、0.86,且这些物种均属于秋季的广生态位种类。窄生态位种类中华哲水蚤和拟长腹剑水蚤均与其他生态位较宽的种类的生态位重叠程度较低,生态位重叠指数范围在0.25~0.67。
2.4 浮游动物优势种群的生态位分化
2.4.1 海域环境因子
曹妃甸近岸海域夏、秋两季各监测站位的主要水环境因子指标见表4。由表4可知,调查海域夏季温度、叶绿素 a和盐度极显著低于秋季(t=-17.458,P=0.000<0.001;t=-3.642,P=0.0005<0.001;t=11.071,P=0.000<0.001),悬浮物浓度夏季显著低于秋季(t=-2.150,P=0.041<0.05)。pH和溶解氧(DO)浓度均相对稳定,差异不显著(t=0.148,P=0.883>0.05;t=0.000,P=1.000>0.05);化学耗氧量(COD)、无机氮(DIN)和活性磷酸盐(DIP)呈现秋季平均值高于夏季,但差异不显著(t=-2.075,P=0.048>0.05;t=0.198,P=0.587>0.05;t=0.477,P=0.880>0.05);浊度(NTU)呈现夏季平均值高于秋季,但差异不显著(t=0.000,P=1.000>0.05)。
2.4.2 冗余分析(RDA)排序结果
选取了温度、盐度、pH、溶解氧、悬浮物、化学需氧量、无机氮、活性磷酸盐和叶绿素a 9个主要环境因子作为分析影响浮游动物优势种群生态位分化的环境因子,采用冗余分析(RDA)来定量分析曹妃甸海域浮游动物种群与各个环境因子的关系(表5和表6),根据前2个排序轴绘制排序图(图 4),物种在排序图上的分布基本能反映其空间分布变化格局。
由表5可知,夏季,第一排序轴和第二排序轴的特征值分别为0.305和0.178,分别解释了物种环境方差的 44.9%和 26.1%,达总标准特征值的71.0%。RDA排序中环境因子共解释了物种组成总变异的63.2%,第一排序轴所包含的物种与环境相关系数为 0.936,第二排序轴所包含的物种与环境相关系数为 0.864。秋季,第一排序轴和第二排序轴的特征值分别为0.274和0.193,分别解释了物种环境方差的 45.1%和 31.8%,达总标准特征值的76.9%。RDA排序中环境因子共解释了物种组成总变异的56.9%,第一排序轴所包含的物种与环境的相关系数为 0.886,第二排序轴所包含的物种与环境的相关系数为 0.963。由此表明,夏、秋季曹妃甸海域环境因子与浮游动物群落组成关系密切。
2.4.3 冗余分析(RDA)排序图分析
RDA相关双序图的解读遵循如下规则:响应变量(物种)与解释变量(环境因子)箭头之间的夹角的余弦值反映它们之间的相关性,夹角越小,相关性越大;响应变量之间、解释变量之间的关系亦如上(Terbraak,1989;Terbtaak,1994)。基于上述规则对图3进行解读。
由表6可知,夏季,物种两排序轴与环境因子两排序轴的相关系数分别为 0,表明排序结果是可信的。由表6和图4(a)可知,与第一物种排序轴呈正相关且影响较大的环境因子依次为无机氮(0.3726)、pH(0.3513)和温度(0.3209),其他环境因子与第一物种排序中轴的相关性系数较小,其中悬浮物、化学耗氧量和叶绿素a与其呈现负相关性。与第二物种排序密切相关的环境因子为无机氮、化学耗氧量和活性磷酸盐,其相关系数分别为0.4496、0.3641和 0.3425,pH、温度、盐度和溶解氧与其呈负相关性。由图4(a)可知,夏季无机氮、pH、温度、化学耗氧量和活性磷酸盐是影响浮游动物优势种生态位分布最大的环境因子。沿第一排序轴,排序图右侧的小拟哲水蚤和双毛纺锤水蚤等类群生态分布受到无机氮、pH和温度的影响;沿第二排序轴,排序图上侧的克氏纺锤水蚤和蔓足类无节幼虫等类群生态分布受到无机氮、化学耗氧量和活性磷酸盐的影响。
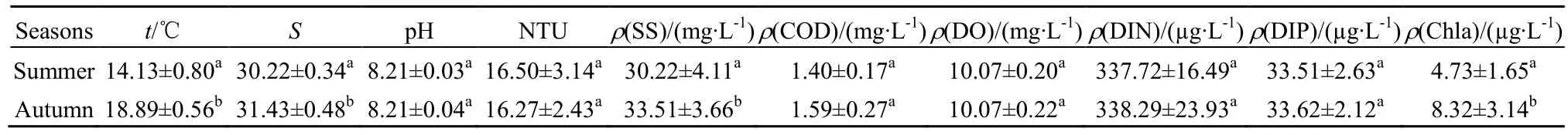
表4 曹妃甸近岸海域夏季和秋季环境因子Table 4 Environmental parameters in the summer and autumn season in the coastal area of Caofeidian
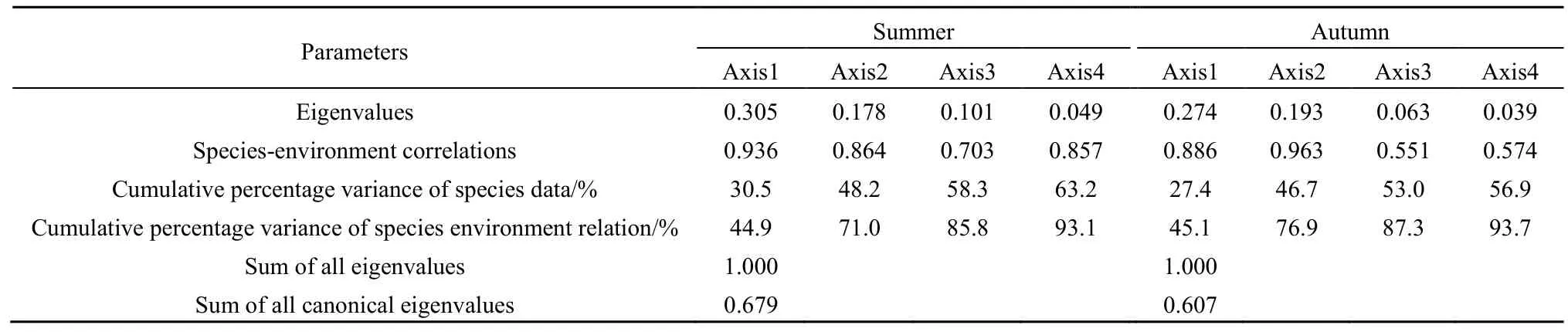
表5 曹妃甸海域浮游动物优势种种类和环境因子间RDA分析的统计结果Table 5 The axes of RDA performed between the dominant zooplankton species and environmental parameters in the coastal area of Caofeidian
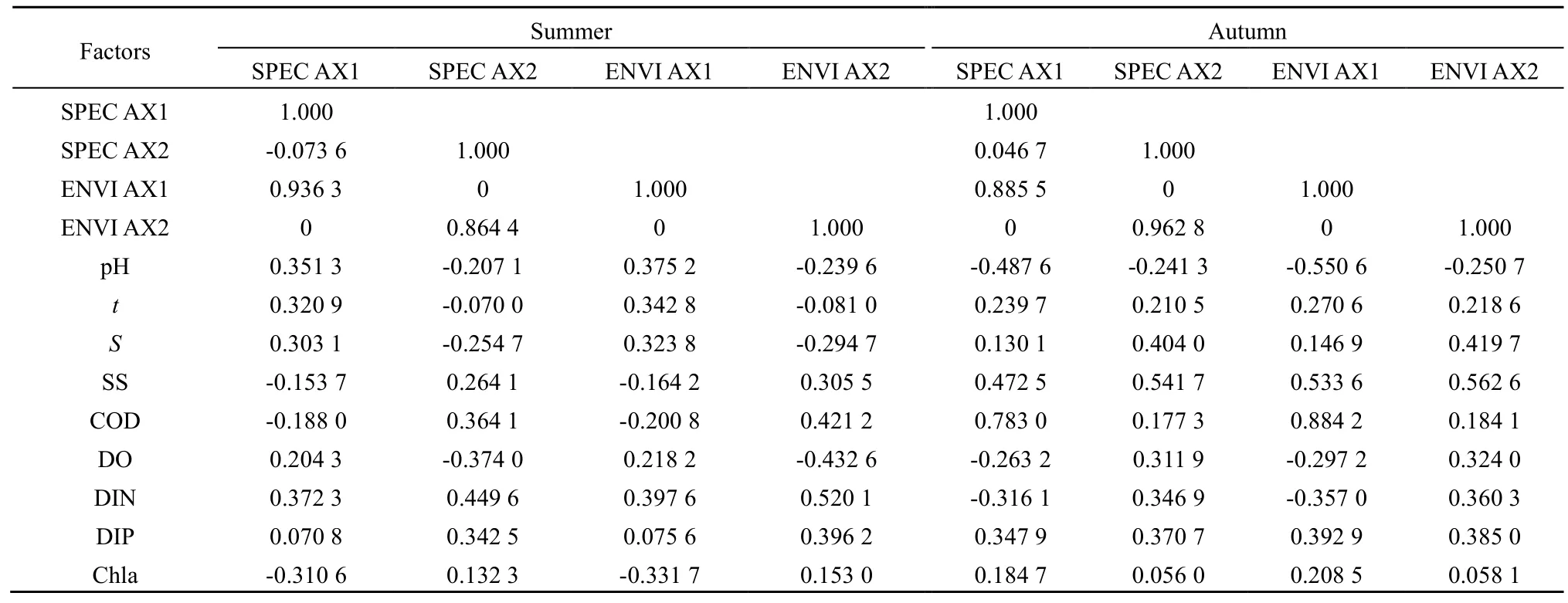
表6 前2个排序轴和环境因子间的相关系数Table 6 Correlation coefficients of environmental factors with the first two axes of RDA
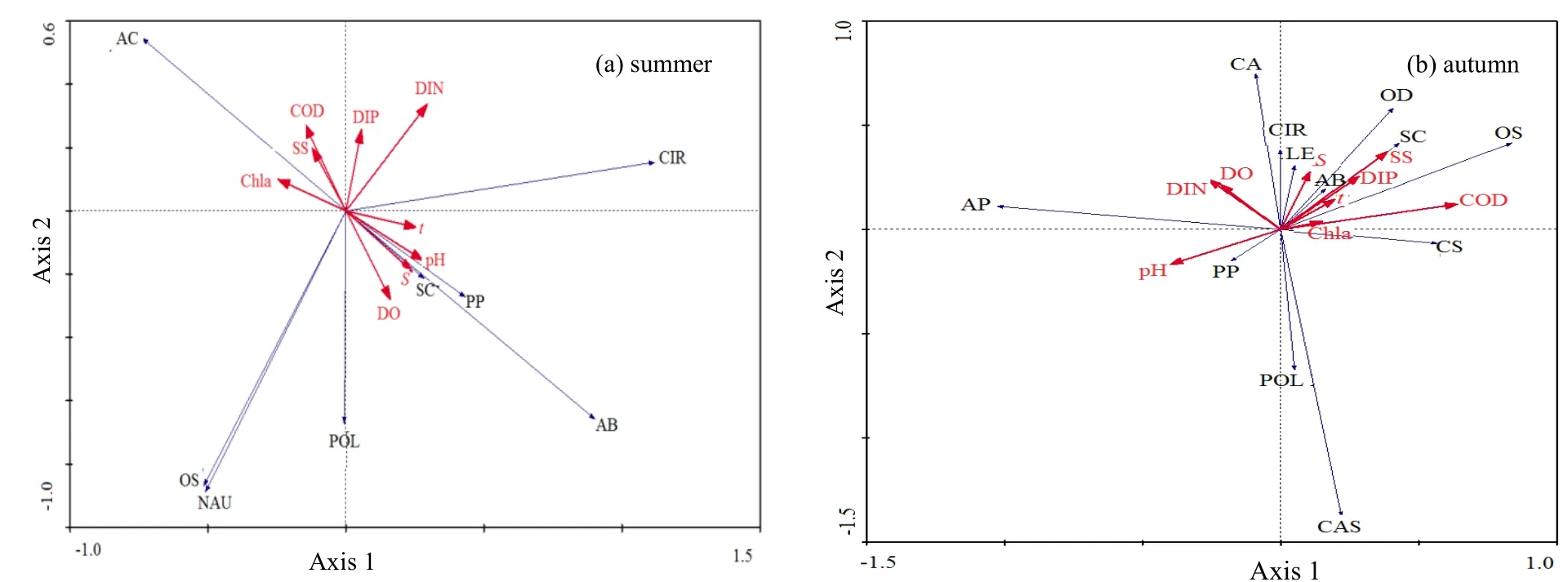
图4 曹妃甸近岸海域浮游动物优势种RDA排序图Fig. 4 RDA ordination graph analysis between the dominant zooplankton species and environment in the coastal area of Caofeidian(a): two-dimensional sort dominant zooplankton species and environment in the summer: AC-Acartia clause, AB-Acartia bifilosa, OS-Oithona similes,PP-Paracalanus parvus, SC-Sagitta crassa, NAU-Nauplius larva, POL-Polychaeta larva, CIR-Cirripedia larva; (b): two-dimensional sort dominant zooplankton species and environment in the autumn: CA-Corycaeus affinis, PP-Paracalanus parvus, CS-Calanus sinicus, AB-Acartia bifilosa, AP-Acartia pacifica,OS-Oithona similes, LE-Labidocera euchaeta, CIR-Cirripedia larva, POL-Polychaeta larva, CAS-Gastropoda larva, OD-Oikopleura dioica, SC-Sagitta crassa
由表6可知,秋季,物种两排序轴与环境因子两排序轴的相关系数分别为 0,表明排序结果是可信的。由表6和图4(b)可知,与第一物种排序轴呈正相关且影响较大的环境因子依次为化学耗氧量(0.7830)、悬浮物(0.4725)和活性磷酸盐(0.3479),其他环境因子与第一物种排序中轴的相关性系数较小,其中 pH、溶解氧和无机氮均与其呈负相关性。与第二物种排序密切相关的环境因子为悬浮物和盐度,其相关系数分别为0.5417和0.4040。由图4(b)可知,秋季化学耗氧量、悬浮物、活性磷酸盐和盐度是影响浮游动物优势种生态位分布最大的环境因子。沿第一排序轴,排序图右侧的拟长腹剑水蚤、异体住囊虫、强壮箭虫等类群生态分布受到化学耗氧量、悬浮物和活性磷酸盐的影响较大;沿第二排序轴,排序图上侧的近缘大眼剑水蚤、异体住囊虫、强壮箭虫等类群生态分布受到悬浮物和盐度的影响。
3 讨论
3.1 曹妃甸近岸海域浮游动物优势种群落特征及其生态位
生态位是种(类)群对多个环境因子生态适应的一个综合结果,它为一个区域内物种的空间分布提供了简单、可靠的生态解释(郑挺等,2014)。生态位宽度反映了种群利用空间资源的多样化程度(郑挺,2014),可反映物种的分布和数量(郭坤等,2016)。强壮箭虫在夏、秋季的出现频率均为 100%,但其平均密度较低,没有成为第一优势种,对浮游动物总丰度贡献较低,分别为2.45%和6.43%。王红等(2015)研究了曹妃甸海域浮游动物种群 1959—2004年的群落结构演变情况,指出了强壮箭虫逐渐失去了优势地位,取而代之的是小拟哲水蚤、拟长腹剑水蚤和双毛纺锤水蚤,本研究结果与其是一致的。但其生态位宽度值在夏、秋两季均较大,属于广生态位种类。由此说明,强壮剪虫更倾向于泛化种,具有更强竞争力(赵惠勋等,1990),故在每个调查站位均有出现。
本研究还发现种(类)群生态位宽度与季节变化有着密切的联系,能够体现浮游动物丰度随季节的动态变化。优势度较高的小拟哲水蚤在秋季是第一优势种,出现频率为100%,其生态位宽度最大,为0.93,属于广生态位种类;而在夏季为第二优势种,出现频率为 92.86%,生态位宽度下降,仅为0.67,为中生态位种类。夏季,强壮箭虫和克氏纺锤水蚤的生态位宽度最大,为广生态位种类。这是由于夏季受降雨影响地表径流增加,海水盐度显著低于秋季,海域内浮游动物种群生态位较宽的强壮箭虫和克氏纺锤水蚤均为低盐种,相比其他浮游动物具有更强的竞争能力,且克氏纺锤水蚤在夏季为第一优势种群进一步验证了生态位宽的物种的竞争能力强这一特性,而小拟哲水蚤优势地位下降,对空间资源的竞争能力下降。秋季,随着降雨的减少,盐度逐渐升高,一些低耐盐物种如克氏纺锤水蚤失去了竞争优势,没有成为秋季优势种,而小拟哲水蚤优势度升高,相应的生态位宽度增大,竞争能力增强,由此说明一个种的生态位越宽,即为广生态位种类,更倾向于泛化种,具有更强竞争力;反之,窄生态位种类更倾向于特化种,在竞争中处于劣势(赵惠勋等,1990),但在生物监测中可被视为指示物种(汪志聪等,2010)。本研究同时印证了生态位宽度是衡量同一生境条件下种群生态优势程度的一个重要指标(徐晓群等,2013)。
生态位重叠反映了两个或多个种(类)群对同一资源因素(食物、营养成分、空间等)的共同利用程度(张金屯,2004),关系到两个种群稳定的生活在一起的生态位相似度(钟章成,1987)。本研究发现,夏季,广生态位强壮箭虫与克氏纺锤水蚤、双毛纺锤水蚤、拟长腹剑水蚤、小拟哲水蚤和多毛类幼虫的生态位重叠程度均较高,生态位重叠指数均在0.7以上,其中强壮箭虫与多毛类幼虫生态位重叠指数最高,为0.82;中生态位种类多毛幼虫与双毛纺锤水蚤、拟长腹剑水蚤和无节幼虫之间生态位重叠指数也在0.7以上。秋季,生态位宽度较大的物种之间有着很高的重叠度,窄生态位的种类与其他生态位较宽的种类的生态位重叠程度较低。这说明,生态位宽度较大的种类之间的生态位重叠程度也较高。Wathne et al.(2000)认为,生态位重叠值大于0.7时,种间生态位重叠较显著,且这些种出现的环境位点重合度较高。而强壮箭虫与生态位宽度较小物种的生态位重叠值则较低,可能是由于物种占据的环境位点重合度较低。同一类群生物往往有着相似的资源利用习性,因此各优势种群生态位必须高度分化才能不形成种间竞争关系,这与它们的生态位重叠度较低的结果相一致。
3.2 曹妃甸近岸海域浮游动物优势种生态位与环境因子的联系
海洋生态系统与环境之间的相互作用是基于多重的时间和空间尺度,相对于海洋中其他的生物群落,浮游动物自主运动能力较弱,具有随波逐流的特点,因此,环境因素对浮游动物的生态分布起着主导作用。浮游动物在不同资源维度下发生生态位分化可以通过排序来表现。排序也叫梯度分析(gradient analysis),是将样方或物种排列在一定的空间,使得排序轴能够反映一定的生态梯度,从而解释物种的分布与环境因子间的关系(Austin,1985;张硕等,2017)。本研究RDA分析从环境因子的角度通过排序手段展示种(类)群分布与生态环境的关系,很好地解释了浮游动物在不同资源维度(环境因子)上的生态位分化。夏季,沿第一排序轴,浮游动物优势种(类)大部分在无机氮、pH、温度、化学耗氧量和活性磷酸盐5个梯度上生态位分化,秋季时浮游动物优势种(类)群的分布发生分散,多数的浮游动物优势种(类)群在化学耗氧量、悬浮物、活性磷酸盐和盐度4个梯度上发生生态位分化,因此,由RDA综合分析得出,无机氮、pH、温度、盐度、化学耗氧量、悬浮物和活性磷酸盐度对浮游动物时空生态分化有直接影响,叶绿素a和溶解氧等因素对其分化的影响次之。叶绿素 a和溶解氧的浓度与浮游植物的分布密切相关,水体理化因子通过控制浮游植物的生长(Abrantes et al.,2006),间接影响浮游动物的动态。
另外,RDA不仅可以从环境因子的角度通过排序手段展示种(类)群分布与生态环境的关系,揭示浮游动物在生境中的生态分化现象,还可以对生态位宽度以及重叠程度计测得到的一些不能合理解释的结果做出补充说明。如夏季,蔓足类无节幼虫与克氏纺锤水蚤的生态位重叠值较低,两者在无机氮、化学耗氧量和活性磷酸盐3个梯度上的分化十分明显,克氏纺锤水蚤是曹妃甸近岸海域典型的近岸低盐种类(王红等,2015),蔓足类无节幼虫主要是仔鱼摄食的饵料生物,生态适应性的不同导致时空分布的差异从而造成两者间重叠度很低。
本研究的监测和分析只能初步反映曹妃甸浮游动物群落的分布与演替,仅从优势种在各个季度样点的分布、生态位宽度和生态位重叠及其与环境因子的变化探讨了优势种群落的时空变化,要详细了解该区域浮游动物的演替尚需进一步展开调查与分析;同时种间竞争和水体中其他高等生物如滤食性鱼类对浮游动物生长的影响还有待深入研究。
4 结论
(1)曹妃甸近岸海域2014年夏季、秋季共鉴定出3大类9种浮游动物优势种和4种浮游幼虫,其中克氏纺锤水蚤和小拟哲水蚤为夏季主要优势种,小拟哲水蚤和双毛纺锤水蚤为秋季主要优势种。
(2)夏季浮游动物群落中生态位宽度值变化范围为0.28~0.89,生态位重叠指数范围为0.24~0.82;秋季浮游动物群落中生态位宽度值变化范围为0.44~0.93,生态位重叠指数范围为0.31~0.95。两季均呈现出广生态位种类之间的生态位重叠程度均较高,生态位重叠指数均在0.7以上;窄生态位的种类与其他生态位较宽的种类的生态位重叠程度较低的特征。
(3)RDA排序结果表明,夏季无机氮、pH、温度、化学耗氧量和活性磷酸盐是影响浮游动物优势种(类)群生态位分布的主要环境因子;秋季主要环境影响因子为化学耗氧量、悬浮物、活性磷酸盐和盐度。
