ERF3促根突变体缓解高浓度CO2对粳稻植株氮吸收的负效应
2018-08-04王伟露袁嫚嫚朱建国刘钢
王伟露 ,袁嫚嫚 ,朱建国,刘钢
1. 中国科学院南京土壤研究所/土壤与可持续农业国家重点实验室,江苏 南京 210008;2. 中国科学院大学,北京 100049
大气CO2浓度持续升高,提高了水稻光合能力,有利于增加地上部干物质积累和产量(Kimball,2016;李春华等,2016)。早先FACE(free air CO2enrichment)研究表明,水稻产量对高浓度 CO2的响应程度依赖品种选择和氮水平(Kim et al.,2003;Hasegawa et al.,2013;Yang et al.,2006)。高 CO2浓度条件下,粳稻增产幅度较低,约为10%~15%,是杂交籼稻品种增幅的一半。随着氮水平的增加,粳稻产量对高浓度CO2的响应有所增强(Kim et al.,2003;Yang et al.,2006)。因此,增施氮肥以增强粳稻品种产量对高浓度CO2的响应十分重要。然而,氮肥过量使用会对环境造成诸多负面效应(Peng,2011)。因此,如何在控制氮肥使用量的前提下,进一步提高粳稻产量对高浓度CO2的响应能力,具有重要意义。
前期研究表明,制约粳稻对高浓度 CO2的响应能力的关键在于叶片净光合速率及每穗颖花数的稳定响应(Zhu et al.,2014)。叶片净光合速率和每穗颖花数的发育和建成均受到氮浓度的调控(Chen et al.,2014;Ding et al.,2014)。然而,高浓度 CO2明显降低了水稻各器官的氮浓度(Terashima et al.,2014)。高浓度CO2条件下各器官氮浓度下降的主要原因包括:(1)生物量增加导致的稀释效应(Gifford et al.,2000);(2)蒸腾下降导致氮素吸收下降(Drake et al.,1997);(3)CO2浓度升高导致植物对氮素的需求降低(Li et al.,2003);(4)CO2浓度升高导致的氮素损失(Pang et al.,2006)。Feng et al.(2015)研究表明,高浓度 CO2下氮浓度在没有生物量稀释的情况下,氮浓度依然降低,表明氮浓度下降可能还与植物对氮的吸收密切相关。Finzi et al.(2007)研究表明,在未来全球大气 CO2升高的背景下,增强植物对氮素的吸收相较于提高氮素利用效率更为重要。这表明提高粳稻氮吸收可能可以缓解高浓度 CO2对氮浓度的负效应,是提高粳稻产量对 CO2升高的响应能力的重要途径。
植物氮吸收与根系的形态结构及根系的功能密切相关(Peng,2011)。前人研究表明,高浓度CO2下,粳稻根系生物量显著增加,但其单位根干重的根系活力在拔节后显著下降(Yang et al.,2008)。庞静等(2005)指出 FACE条件下,粳稻单茎的根系活力下降,且无机氮含量呈下降趋势,这说明根系活力同氮素的吸收存在内在的联系。增施氮肥可以提高水稻根系活力,同时根系活力的增加可促进氮素吸收,提高叶片净光合速率、籽粒灌浆强度、结实率等(王余龙等,1992;叶宝兴等,2005;戢林等,2012;陈云风,2015)。通过增强根系活力促进氮的吸收能力能否缓解高浓度 CO2下氮浓度降低的现象目前仍为未知。
乙烯响应因子家族(ERF)是一类调节植物生长发育和胁迫响应的转录因子。早先的研究多集中于 ERF在生物以及非生物胁迫方面的调节作用(Xu et al.,2008;Zhang et al.,2013)。近年来,对ERF响应因子的研究发现了其更多新的功能。Zhao et al.(2015)研究表明,OsERF3过表达植株产生更大的根系统,形成更多冠根,主根变长。理论上ERF3过表达株系单茎根系活力会有所提高。为了探究高浓度 CO2下根系形态的增大及其活力的增强是否有利于缓解大气 CO2浓度升高对粳稻氮吸收的负效应,本研究利用生长箱和 FACE平台,以促根突变体ERF3及其野生型为研究对象,考察二者根系形态及活力、组织氮浓度、氮素吸收和利用效率、叶片净光合速率、地上部生物量对CO2浓度升高的响应,以期为进一步提高粳稻物质生产和产量响应幅度提供理论依据。本研究假设:(1)促根突变体ERF3可以提高根系活力;(2)ERF3突变体在高浓度 CO2下可以提高植株氮吸收,避免组织氮浓度的下降;(3)ERF3突变体可以提高粳稻叶片净光合速率和物质生产对高 CO2浓度的响应幅度。
1 材料与方法
1.1 供试材料和培养条件
水稻品种中花11(粳稻)为野生型供试品种,OsERF3突变体以中花11为遗传背景(Zhao et al.,2015)。
试验A于2016年4月在中国科学院南京土壤所生长箱进行,对照和 CO2处理分别设置为 400 μmol·mol-1和 600 μmol·mol-1。于 4 月 3 日先用 H2O2将水稻种子消毒30 min后浸种,催芽2~3 d后用1/2浓度的培养液育秧12 d后移栽至7 LPVC面包盒中进行试验处理28 d。营养液采用国际水稻所的配方配制(略做修改):1.25 mmol·L-1NH4NO3,0.3 mmol·L-1KH2PO4,0.35 mmol·L-1K2SO4,1 mmol·L-1CaCl2·2H2O,1 mmol·L-1MgSO4·7H2O,0.5 mmol·L-1Na2SiO3·9H2O , 9 μmol·L-1MnCl2·4H2O , 0.39 μmol·L-1Na2MoO4·2H2O,20 μmol·L-1H3BO3,0.37 μmol·L-1ZnSO4·7H2O,0.32 μmol·L-1CuSO4·5H2O,20 μmol·L-1FeSO4·7H2O+Na2-EDTA。每天用 NaOH调节pH为5.5。两个生长箱相对湿度为70%左右,昼夜温度分别为 29 ℃和 21 ℃,光合有效辐射(PAR)为 800~1000 μmol·m-2·s-1。
试验 B于江苏省江都市小纪镇马凌村良种场(119°42′0″E,32°35′5″N)进行,该地区年降雨量约1000 mm,年均温约 15 ℃,年均日照时间大于2000 h。平台共有3个 FACE试验圈和3个对照(Ambient)圈。FACE圈之间以及FACE圈与对照圈之间的间隔大于90 m,以减少CO2择放对其他圈的影响。FACE圈设计为正八角形,直径12 m,平台运行时通过 FACE圈周围的管道向中心喷射纯CO2气体,利用计算机网络对平台的CO2择浓度进行监测和控制,根据大气中CO2浓度、风向、风速、作物冠层高度的CO2浓度及其昼夜变化等因素自动调节CO2气体的释放速度及方向,使水稻主要生育时期FACE圈内CO2浓度保持比大气背景CO2浓度高 200 μmol·mol-1。对照田块没有安装 FACE 管道,其他环境条件与自然状态一致(Yang et al.,2006)。采用盆栽试验,于2016年稻季开展,5月25日播种大田育秧,6月20日移栽至盆栽中。每盆3穴,每穴2苗。盆钵上下长、宽分别为20 cm和10 cm,高30 cm,每盆装风干土6.5 kg。土壤质地为沙壤土,氮肥按照大田 250 kg·hm-2纯氮施用,基肥∶分蘖肥∶穗肥比例为 4∶3∶3。其中,基肥为每盆 1.54 g尿素(46% N)、无水磷酸二氢钠1.31 g(26% P),氯化钾0.5 g(52% K)。分蘖肥和穗肥分别于移栽后7 d和倒3.5叶期(抽穗前31 d)一次性水溶施入,共处理 24盆。适时进行病虫草害防治,保证水稻正常生长发育。
1.2 取样和测定
1.2.1 样品采集
试验A和试验B盆栽分别于处理后28 d,拔节后15 d采样。其中试验A取植株根系用于形态结构考察,而后与地上部样本在 80 ℃烘箱中烘干至恒重,粉碎机研磨。选取部分鲜样于液氮中速冻后转移至-80 ℃冰箱,用于根系活力的测定。试验 B地上部和根系样本在 80 ℃烘箱中烘干至恒重,粉碎机研磨。
1.2.2 根系指标测定
考察试验A中根系冠根数及扎根深度。采用根系扫描仪(WinRHIZO;Regent Instruments Inc.,Quebec,ON,Canada)分析根系长度。根系活力采用α-萘胺法测定(李合生,2000),略做修改。称取1 g新鲜根样,转入150 mL 含有20 mg·L-1α-萘胺的三角瓶中。将三角瓶置于摇床上,在室温条件下孵育3 h。过滤,吸取2 mL滤液,加入1 mL亚硝酸钠(NaNO2,1.18 mmol·L-1)和 1 mL 对氨基苯磺酸混匀。用分光光度计测定510 nm吸光度值。余下的根系样本在70 ℃烘干至恒质量。以单位根干质量α-萘胺的氧化量表征根系活力,单位为 µg·g-1·h-1。
1.2.3 光合参数测定
试验B盆栽试验于取样前,釆用Li-Cor 6400光合测定系统测定新完全展开叶的净光合速率(A)。测定时间为 09:30—15:00,对照和处理 CO2浓度分别设置为 390 μmol·mol-1和 590 μmol·mol-1。测定光强设定为 1800 μmol·m-2·s-1,流量为 500 mL·s-1,叶室温度为(30±2) ℃,相对湿度维持在 60%左右。测定后的叶片用液氮速冻后转移至-80 ℃冰箱保存,用于测定 1, 5-二磷酸加氧/羧化酶(Rubisco)。
1.2.4 氮素和Rubisco含量测定
烘干的植株样本研磨粉碎后经浓 H2SO4-H2O2消煮,用全自动定氮仪测定样本氮含量。叶片中Rubisco含量的测定参照Li et al.(2013)方法。取0.5 g叶片加液氮并迅速研磨成粉状,加入5 mL提取液[50 mmol·L-1Tris-HCl(pH=8.0),5 mmol·L-1β-mercaptoethano和12.5%(V/V)glycerol]匀浆,后在1500 g、4 ℃条件下离心15 min。上清液与2%(W/V)SDS、4%(V/V)β-mercaptoethanol和 10%(V/V)glycerol溶液混合,在沸水中煮 5 min。SDS-PAGE凝胶电泳参照Li et al.(2013),电泳后,凝胶用0.25%考马斯亮蓝染色12 h,脱色。切除含有大亚基和小亚基的凝胶,转移至10 mL含有2 mL甲酰胺的试管中,50 ℃水浴8 h。用分光光度计测定595 nm处吸光值。
地上部生物量生产效率(NUEb,mg·mg-1)为地上部生物量与地上部氮素总吸收量之比。
1.3 数据处理
运用Microsoft Excel软件整理数据,SPSS 19.0软件进行一般线性模型(GLM)多因素方差分析,样本间差异采用最小显著差数法(Least significant difference test),显著性水平设置为 α=0.05。运用SigmaPlot 12.5绘图。
2 结果与分析
2.1 突变体和野生型粳稻根系的形态及生物量对高浓度CO2响应特征
如图1所示,生长箱对照条件下,野生型和突变体总根长无显著差异(P>0.05)。突变体冠根数和扎根深度略高于野生型,增幅分别为7.6和5.3%,未达显著水平(P>0.05)。在高浓度 CO2下,野生型总根长、冠根数以及扎根深度分别显著增加86.4%、35.9%和35.9%,突变体ERF3总根长、冠根数以及扎根深度分别显著(P<0.05)增加124.0%、79.7%和 42.3%,这表明 ERF3根系形态对高浓度CO2的响应能力更强。由图2可知,对照条件下,生长箱和田间盆栽中ERF3突变体根系生物量与野生型无显著差异(P>0.05)。在高浓度 CO2下,生育前期(生长箱)突变体ERF3根系生物量的增幅大于野生型,生育后期(田间盆栽)ERF3和野生型根系生物量增幅相似。
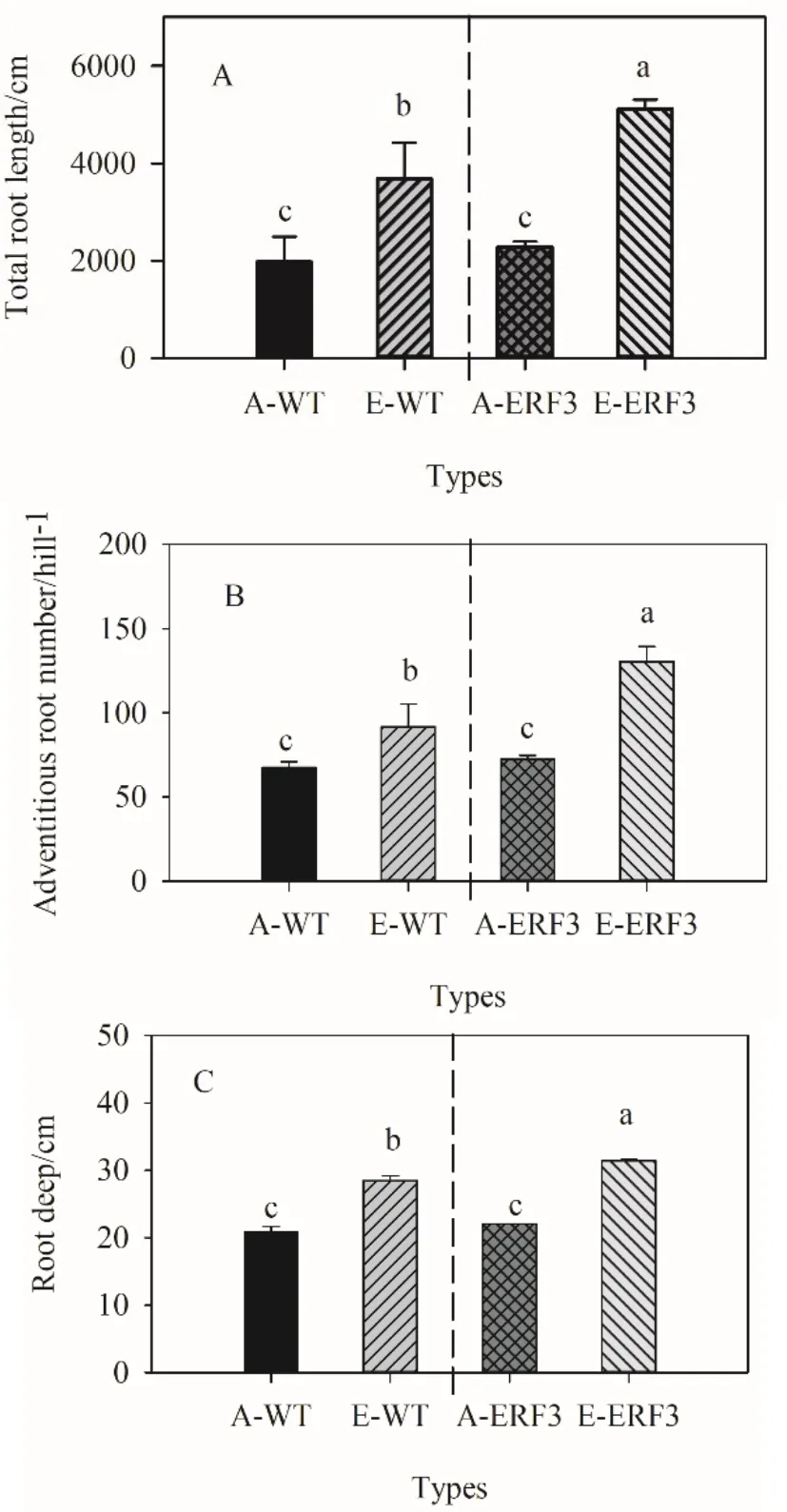
图1 生长箱中CO2浓度升高对野生型和突变体根系形态指标的影响Fig. 1 Root morphology parameters of wild type (WT) and ERF3 mutant under elevated CO2 in growth chamberA:对照;E:CO2+200 μmol·mol-1。WT:野生型;ERF3为 OsERF3突变体。样品重复数n=3。不同字母表示在0.05水平上差异显著。下同A, ambient; E, elevated CO2 treatment (+200 μmol·mol-1). WT, wild type; ERF3, OsERF3 mutant. n=3. Different letters indicate significant difference at P<0.05. The same as below
2.2 突变体和野生型根系活力对高浓度CO2的响应
由图3可知,生长箱对照条件下,突变体单茎根系活力同野生型相比无显著变化(P>0.05)。但是,高浓度CO2条件下,突变体ERF3单茎根系活力显著(P<0.05)提高212.1%,野生型根系活力仅增加16.6%。
2.3 突变体和野生型地上部生物量对高浓度 CO2的响应
由图4A可知,生长箱对照条件下,ERF3突变体地上部生物量略高于野生型。野生型和突变体ERF3地上部生物量对高浓度CO2的响应显著增加(P<0.05),分别为65.1%和156.9%。如图4B所示,田间盆栽对照条件下,野生型和突变体地上部生物量无显著差异(P>0.05)。高浓度 CO2显著(P<0.05)增加野生型和突变体中茎秆生物量,分别增加32.1%和57.7%。突变体ERF3叶片生物量显著增加(P<0.05)。
2.4 突变体和野生型组织氮浓度对高浓度 CO2的响应
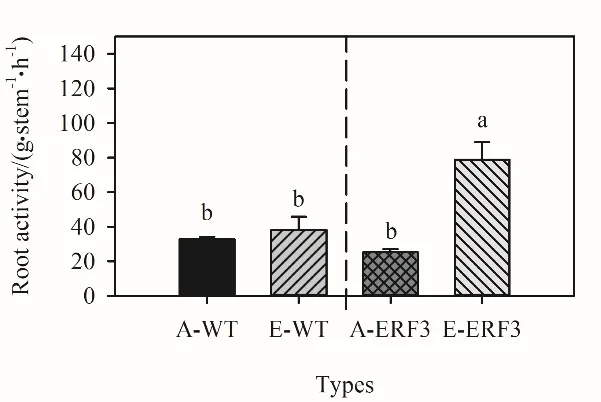
图3 生长箱CO2浓度升高对野生型和突变体根系活力的影响Fig. 3 Amount of a-NA per culm of wild type (WT) and ERF3 mutant under elevated CO2 in growth chamber
由图5可知,生长箱对照条件下,野生型地上部氮浓度显著(P<0.05)高于促根突变体 ERF3,这与田间盆栽试验结果类似。高浓度CO2下,生长箱中野生型地上部氮浓度显著(P<0.05)降低16.4%,突变体则无显著变化(P>0.05)。田间盆栽结果表明,野生型叶片氮浓度显著(P<0.05)降低16.9%,茎秆氮浓度亦有降低趋势但未达显著水平(P>0.05)。突变体ERF3则表现为叶片氮浓度显著(P<0.05)增加11.7%,茎秆中氮浓度也有增加的趋势,但未达显著水平(P>0.05)。

图2 生长箱(A)和田间盆栽(B)CO2浓度升高对野生型和突变体根系生物量对的影响Fig. 2 Root biomass (dry) of wild (WT) and ERF3 mutant under elevated CO2 in growth chamber (A) and field (B)
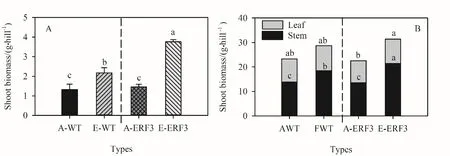
图4 生长箱(A)和田间盆栽(B)CO2浓度升高对野生型和突变体地上部生物量的影响Fig. 4 Shoot dry matter of wild type (WT) and ERF3 mutant under elevated CO2 in growth chamber (A) and field (B)

图5 生长箱(A)和田间盆栽(B,C)CO2浓度升高对野生型和突变体各组织氮质量分数的影响Fig. 5 Nitrogen concentration of wild type (WT) and ERF3 mutant under elevated CO2 in growth chamber (A) and field (B, C)
2.5 突变体和野生型氮素吸收对高浓度CO2的响应
从图6A可知,生长箱对照条件下,野生型和突变体氮素累积无显著差异(P>0.05)。高浓度CO2分别显著(P<0.05)提高了野生型和突变体 ERF3氮吸收量,增幅分别为40.6%和113.5%。由图6B可知,田间盆栽对照条件下,突变体ERF3地上部氮素累积显著(P<0.05)低于野生型,主要归因于叶片中氮素累积的降低。CO2浓度升高显著(P<0.05)增加了突变体叶片和茎秆中氮素累积量,分别达23.8%和77.0%。
2.6 突变体和野生型氮素利用效率对高浓度 CO2的响应
从图7A可知,生长箱中野生型和突变体氮素干物质生产效率分别增加16.7%和21.3%,仅突变体增幅达显著水平(P<0.05)。由图7B可知,田间盆栽试验,野生型氮素干物质生产效率显著(P<0.05)增加20.3%,而突变体ERF3氮素生物量利用率则未发生显著变化(P>0.05)。
2.7 突变体和野生型叶片光合参数对高浓度 CO2的响应
从图8A可知,对照条件下,野生型和突变体叶片净光合速率无显著差异(P>0.05)。FACE条件下,野生型和突变体ERF3叶片净光合速率均显著(P<0.05)增加,增幅分别为13.3%和42.8%。由图8B可知,野生型光合关键酶 Rubisco含量显著(P<0.05)降低24.8%,突变体ERF3则未发生显著(P>0.05)变化。
3 讨论

图6 生长箱(A)和田间盆栽(B)CO2浓度升高对野生型和突变体各组织氮吸收的影响Fig. 6 Nitrogen uptake of wild type (WT) and ERF3 mutant under elevated CO2 in growth chamber (A) and field (B)

图7 生长箱(A)和田间盆栽(B)CO2浓度升高对野生型和突变体地上部氮素干物质生产效率(NUEb)的影响Fig. 7 Nitrogen use efficiency (NUEb) of wild type (WT) and ERF3 mutant under elevated CO2 in growth chamber (A) and field (B)

图8 大田盆栽野生型和突变体叶片净光合速率对CO2浓度升高的响应Fig. 8 Leaf net photosynthesis rate of wild type (WT) and ERF3 mutant in FACE site
水稻拥有复杂的根系统,主要由主胚根、侧根和不定根组成(Yoshida,1981)。根系的形态结构与植物对养分的吸收、干物质生产及产量密切相关(Zhang et al.,2009;杨建昌,2011)。前人研究表明,从分蘖盛期开始,大气CO2浓度升高显著增加粳稻总根长、冠根数、根系体积及根系生物量(Kim et al.,2003;陈改苹等,2006;Yang et al.,2008),这是粳稻物质生产和产量在高CO2浓度下显著提高的原因之一。本研究结果表明,高 CO2浓度显著(P<0.05)增加野生型和突变体总根长、冠根数、扎根深度及根系生物量,这与前人的研究结果一致。此外,突变体ERF3根系形态各指标和根系生物量对高 CO2浓度的响应强度高于野生型,约高出6.4%~191.0%。这表明,高CO2浓度条件下,突变体ERF3根系形态结构较野生型具有更明显的增幅潜力。前人认为地上部和根系的生长对碳水化合物的需求是竞争的关系,活跃的地上部生长可为根系的形态建成和发育提供充足的碳水化合物;根系的活跃生长也可为地上部提供充足的养分(Zhang et al.,2009)。因此,根系生物量的增加与地上部生物量对高CO2浓度的响应存在密切的关系。然而,根系生物量的形成所消耗的碳水化合物约为地上部的2倍,过大的根系会抑制地上部的生长,反过来阻碍根系的生长和发育。这可以解释为什么在试验初期(育秧时期),ERF3在对照条件下发根能力(根长和冠根数)强于野生型,而随着生育期的推进,ERF3与野生型之间根系形态参数的差异逐渐缩小,即促根生长消耗了过多的碳水化合物。
根系氧化能力被认为是表征根系生理活性的重要参数(Yang et al.,2004)。较高的根系活力对于维持根系生物量、根系生长和养分吸收非常重要(Zhang et al.,2009;戢林等,2012;陈云风,2015)。根系活力的强弱多以单位时间内单位根干重对 α-萘胺的氧化量表征,而后者与根系过氧化氢酶的氧化能力有关。根系和地上部充足的碳水化合物一方面为过氧化氢酶的合成提供充足的碳骨架,另一方面为过氧化氢酶的氧化过程提供所需的能量物质ATP。前人研究结果表明,高CO2浓度降低了粳稻单茎根系活力(庞静等,2005;Yang et al.,2008)。本研究并未在野生型结果中发现类似的现象,这可能是由于水培试验周期较短,根系和地上部生物量对高CO2浓度的响应依然较高。突变体ERF3根系和地上部生物量优势明显是其单茎根系活力对高CO2浓度的响应能力强于野生型的重要原因。
前人研究结果均表明,高CO2浓度降低了粳稻各器官氮浓度(Kimball,2016),这与本研究结果一致。氮浓度的下降减缓了粳稻物质生产和产量对高CO2浓度的进一步响应(Easlon et al.,2013)。前人针对水稻植株组织氮浓度降低这一现象提出了不同的假设,如稀释效应,氮需求下降,氮素吸收能力降低(Drake et al.,1997;Gifford et al.,2000;Li et al.,2003)。Finzi et al.(2007)指出在氮素供应充足的条件下氮素的利用率增加,降低了植物对氮素的需求。Feng et al.(2015)采用荟萃分析指出氮素的吸收才是作物氮浓度下降的主要原因。植物对氮素的吸收不仅与根系形态有关,而且与根系活力密切相关(Kiba et al.,2016)。当根氧化力增大时,根的有氧呼吸较旺盛,吸收养分也较多(Zhang et al.,2009;杨建昌,2011)。中国FACE平台早先的研究结果表明,在氮肥供应充足的情况下,尽管粳稻总根长、冠根数以及根系生物量显著增加,但是单茎根系活力(活跃吸收面积以及 α-萘胺氧化量)显著降低(庞静等,2005;Yang et al.,2008),这导致了粳稻后期氮素吸收能力的降低。本研究结果表明,高CO2浓度条件下,促根突变体ERF3根系形态和单茎根系活力均显著增加(P<0.05),增幅高于野生型。突变体ERF3根系活力对高CO2浓度的响应较野生型具有更明显的优势。这一根系特性使得突变体ERF3在生长箱和田间盆栽试验中的氮素吸收较野生型显著增加(P<0.05),并且ERF3在生长箱中的氮素利用效率对高CO2浓度的响应能力高于野生型,从而避免了各器官氮浓度的降低。这表明大根系以及根系活力增强的突变体ERF3确实可以缓解高CO2浓度对粳稻氮吸收的负效应。
众所周知,高CO2浓度显著提高水稻叶片净光合速率和物质生产,这与本试验结果一致(Chen et al.,2014)。然而,粳稻在生育后期叶片净光合速率对高浓度CO2的响应幅度的降低制约了干物质和产量的进一步提高(Zhu et al.,2014;Kimball,2016)。水稻叶片净光合速率同氮素存在密切关系,尤其是光合关键酶Rubisco(Chen et al.,2014)。在本试验中,促根突变体ERF3较高的氮素吸收和分配使得叶片Rubisco含量保持稳定,这确保了叶片净光合速率和地上部生物量对高浓度CO2的持续高响应。有研究指出,氮素同细胞分裂素协作调控着叶片发育和光合特性(Kurakawa et al.,2007)。外施细胞分裂素可以提高氮素向光合关键酶 Rubisco的分配(Ookawa et al.,2004),根系是细胞分裂素合成的主要器官。因此,高CO2浓度条件下促根突变体ERF3细胞分裂素与氮素在调节叶片净光合速率响应能力的作用方面还有待进一步研究。此外,粳稻氮素营养状况的改善也可能有助于缓解每穗颖花数的下降,提高稻米中蛋白质的含量,改善稻米营养品质。以上结果表明,高浓度CO2条件下,突变体ERF3较大的根系和增强的根系活力可以缓解粳稻氮吸收的负效应,有利于进一步提高粳稻物质生产对高浓度CO2的响应幅度。
4 结论
高浓度CO2条件下促根突变体ERF3大根系和根系活力增强的特性,促进了根系对氮素的吸收,避免地上部器官中氮浓度的降低。充足的氮素吸收,维持了突变体ERF3叶片氮素和Rubisco的含量,从而提高叶片净光合速率和物质生产的响应能力。在未来的育种中,可以考虑通过促根增强粳稻对未来气候变化的适应能力。
