欧洲卫矛秋冬转色期叶色变化的生理机制
2018-07-19卓启苗丁彦芬朱贵珍
卓启苗,丁彦芬,余 慧,朱贵珍
(南京林业大学 风景园林学院,南京 210000)
欧洲卫矛(Euonymuseuropaea)是卫矛科(Celastraceae)卫矛属(Euonymus)植物,落叶灌木或小乔木,原产欧洲及西亚地区。经过长期的园艺栽培与育种,欧洲卫矛现如今已拥有丰富的园艺品种,是欧洲及北美地区著名的秋季观果及色叶俱佳的植物。欧洲卫矛秋色叶观赏期从9月份开始直到11月份,红叶满枝,极具观赏特色和应用价值。在园林配置时,可点缀于林缘,或与其他小灌木丛植。目前,欧洲卫矛在中国华东、华北地区均有引种,值得进一步推广种植。
通常,秋色叶植物秋季叶色变红是由于叶片中花色素苷大量合成,使得叶片中的色素种类和比例发生了变化[1]。研究认为,可溶性糖与花色素苷含量呈显著性正相关[2],同时花色素苷合成相关酶PAL(苯丙氨酸解氨酶)[3]以及降解相关酶POD(过氧化物酶)、PPO(过氧化物酶)[4]均影响着花色素苷的合成积累。前人对欧洲卫矛的研究主要集中于生态习性[5]、药用价值[6-7]、引种繁殖[8-9]、光合特性[10]等方面,尚未有关于欧洲卫矛叶片秋季变色机理方面的报道。因此,本研究将以欧洲卫矛的3个品种‘矮生’(Euonymuseuropaea‘Pumilis’)、‘八仙花’(E.europaea‘Aldenhamensis’)和‘白果’(E.europaea‘Albus’)为试验材料,测定9月中旬至11月中旬转色期内叶色参数、叶绿素、类胡萝素、花色素苷、可溶性糖含量及PAL、POD、PPO等酶活性的变化,分析各种色素与叶色参数、可溶性糖、相关酶的关系,对欧洲卫矛在秋冬转色期间叶色变化的生理特性进行初步探讨,为欧洲卫矛呈色机理的理论研究提供定量数据分析和理论指导,最终为欧洲卫矛的园林应用提供依据。
1 材料和方法
1.1 试验材料
试验地点位于江苏句容市乾景天园苗木有限公司基地,以欧洲卫矛3个品种‘矮生’(Euonymuseuropaea‘Pumilis’)、‘八仙花’(E.europaea‘Aldenhamensis’)和‘白果’(E.europaea‘Albus’)为试材,各品种均为3年生扦插苗,株高2.5 m,各品种长势一致,无病虫害。
1.2 试验方法
每个品种选取15株作为样品,每5株为1次重复,共3个重复。各材料于2017年9月下旬至11月下旬,每隔10 d取样1次。取样于每天上午10:00前后进行,每次取树冠中部向阳面相同叶位的叶片。
叶片取回后,立即洗净,擦干,去掉主脉后剪碎,混匀,用于叶绿素、花色素苷、可溶性糖含量及酶活性的测定,每个处理设3次重复。叶色参数的测定参照郭欢欢等[11]的方法;叶绿素和类胡萝素含量测定采用丙酮浸提法[12];花色素苷含量测定采用盐酸甲醇浸提法[13];可溶性糖含量测定采用蒽酮比色法[12];苯丙氨酸解氨酶(PAL)、过氧化物酶(POD)活性测定参照Lister等[14]和李合生[15]的方法;多酚氧化酶(PPO)活性测定参照李明等[16]、杨暖[17]的方法。
1.3 数据分析
采用Excel 2010和SPSS 24.0软件对数据进行统计分析,使用Excel 2010作图。
2 结果与分析
2.1 欧洲卫矛叶色参数变化特征
叶色参数是利用色差计来定量表征色泽在三维空间中变化的变量值。L*用来表示光泽明亮度,其值越大明亮度越高。a*值是红/绿的比值,正值表示偏红,负值表示偏绿。b*值是黄/蓝程度,b*值越大,呈现黄色越深;反之,蓝色越深。欧洲卫矛在整个转色期过程中,品种‘八仙花’和‘白果’叶色参数L*值的变化趋势相同,均先上升后下降,而品种‘矮生’的L*值先下降后上升;3个品种L*值总体上变化幅度不大,‘矮生’品种叶色明亮度相较于‘八仙花’、‘白果’品种偏暗(图1,A)。同时,3个品种在转色期间的叶色参数a*值均呈逐渐上升趋势,且品种间a*值差异不大,叶片颜色也均逐渐由绿转红;其中,‘矮生’品种a*值在10月30日前逐渐上升的趋势较平稳,10月30日后急剧上升(图1,B)。另外,3个品种b*值的变化趋势与a*值变化规律相似;品种‘八仙花’、‘白果’a*值始终高于同期品种,叶色较于‘矮生’偏黄(图1,C)。
2.2 欧洲卫矛叶片质体色素含量的变化特征
由图2可见,3个欧洲卫矛品种在叶色变红的过程中,质体色素含量整体呈一致的下降趋势;叶绿素a、叶绿素(a+b)含量明显下降,叶绿素b、类胡萝素以及叶绿素a/叶绿素b下降速率变化较为平稳,总体上变化不大。可见,欧洲卫矛叶片叶绿素总含量的下降主要是由于叶绿素a含量的下降,这与崔珂婉的结果相一致,认为是植物对降温、光照时间缩短等不利环境的适应[18]。
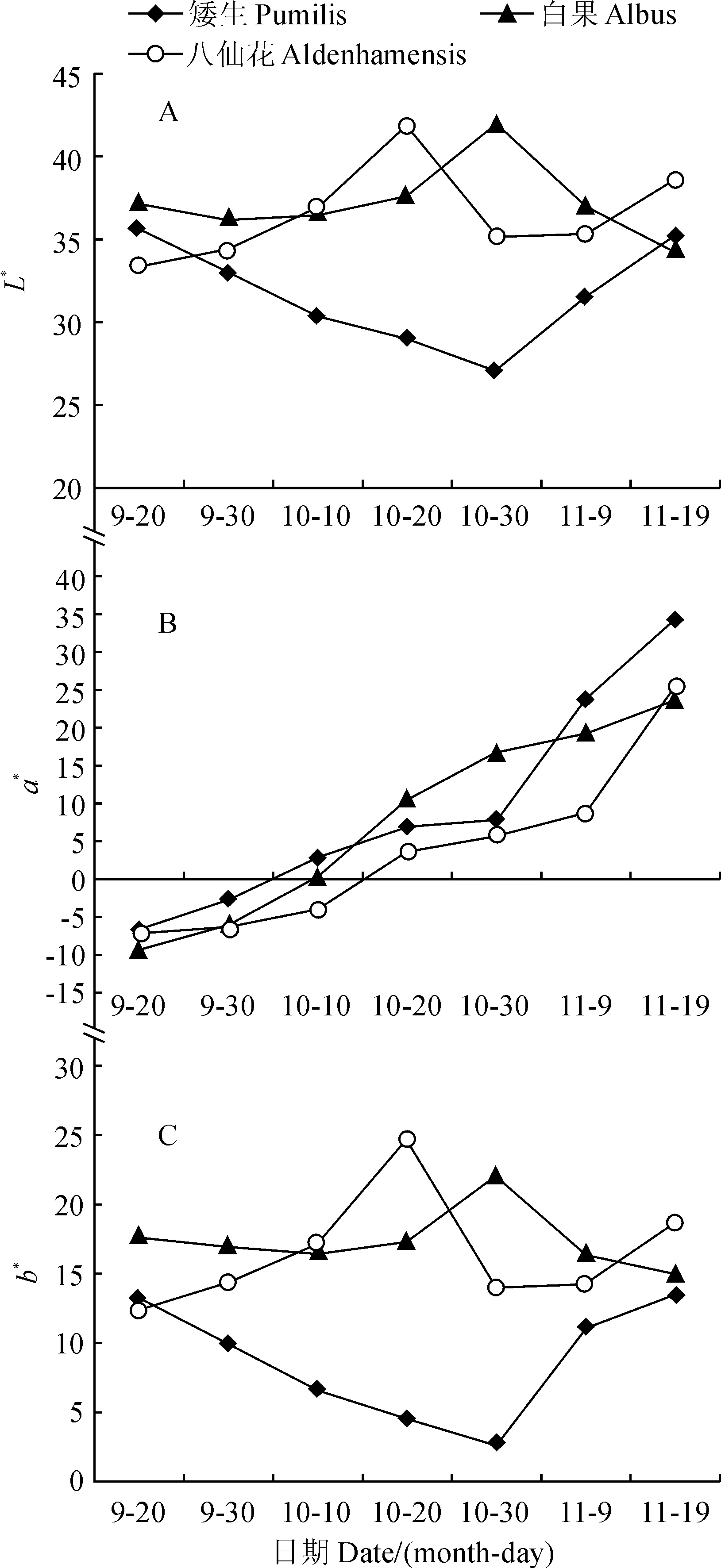
图1 3个欧洲卫矛品种叶色参数L*、a*、b*值的变化Fig.1 The changes of leaf color parameter L*, a* and b* in three E. europaea varieties
2.3 欧洲卫矛叶片花色素苷相对含量及花色素苷/叶绿素、类胡萝素/叶绿素比值的变化
3个欧洲卫矛品种叶片中花色素苷的含量在整个转色期不断升高(图3,A)。其中,花色素苷的含量在前期上升均较为缓慢,10月30日‘八仙花’和‘白果’上升较前期略快,‘矮生’从10月30日起急剧上升,这与叶色参数a*值变化趋势相似(图1,B)。可见,欧洲卫矛转色初期叶色变化程度较小,后期随着花色素苷的大量合成叶色变化较为明显,并因花色素苷含量达到最大值而叶色相对保持不变。同时,在欧洲卫矛叶片整个转色过程中,花色素苷/叶绿素比值的表现为上升趋势(图3,B),这与花色素苷含量变化趋势基本一致,且后期上升速率增大。花色素苷含量增加在各色素比例中占有绝对优势是欧洲卫矛叶片变红的直接原因。另外,欧洲卫矛叶片类胡萝素含量在整个转色期间总体质量分数降低,但无明显的变化趋势(图1),其类胡萝素/叶绿素比值总体呈下降趋势(图3,C),从而可知类胡萝素对欧洲卫矛叶片变红影响不大。
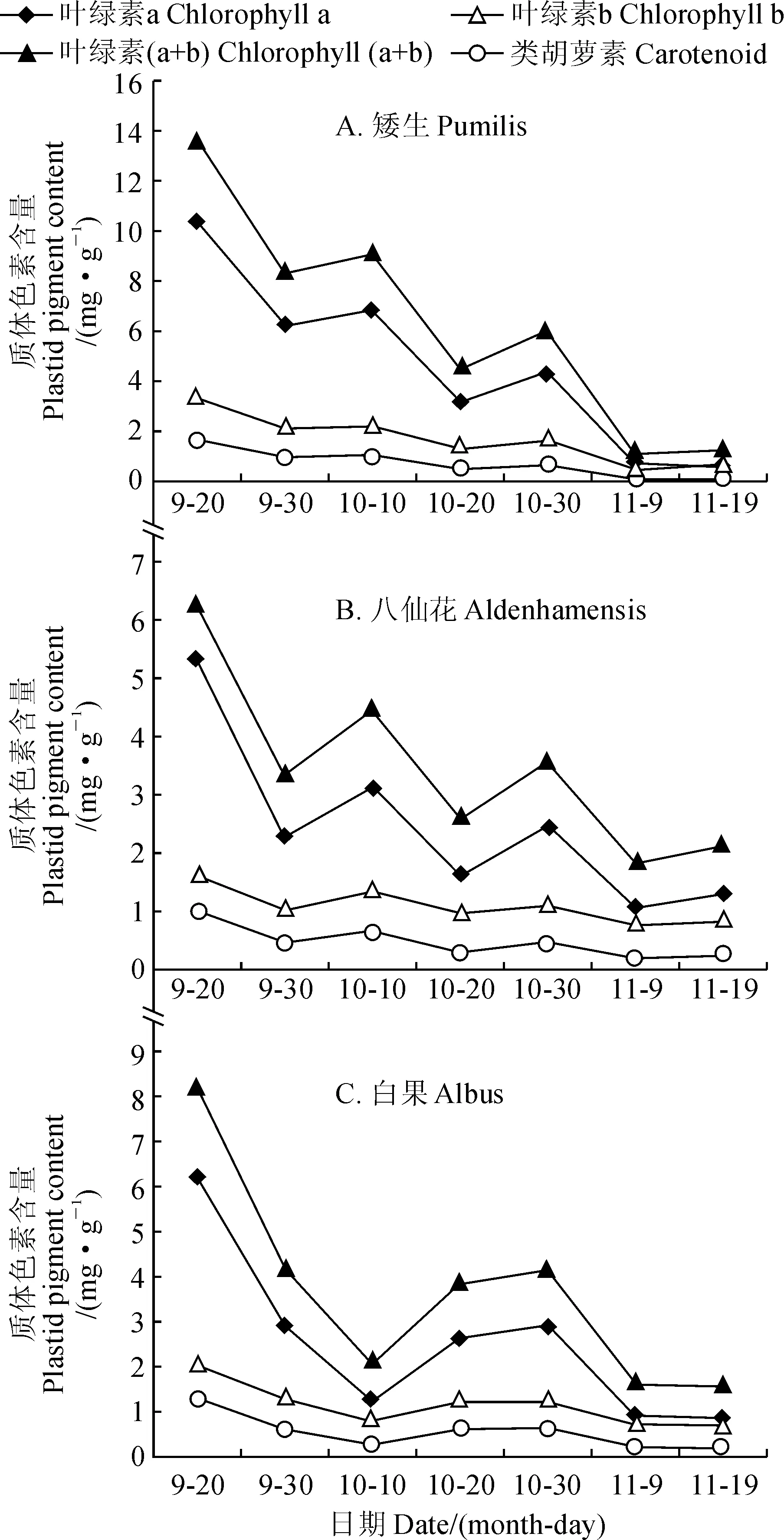
图2 3个欧洲卫矛品种叶片中质体色素含量的变化Fig.2 The changes in plastid pigment content in leaves of three E. europaea varieties
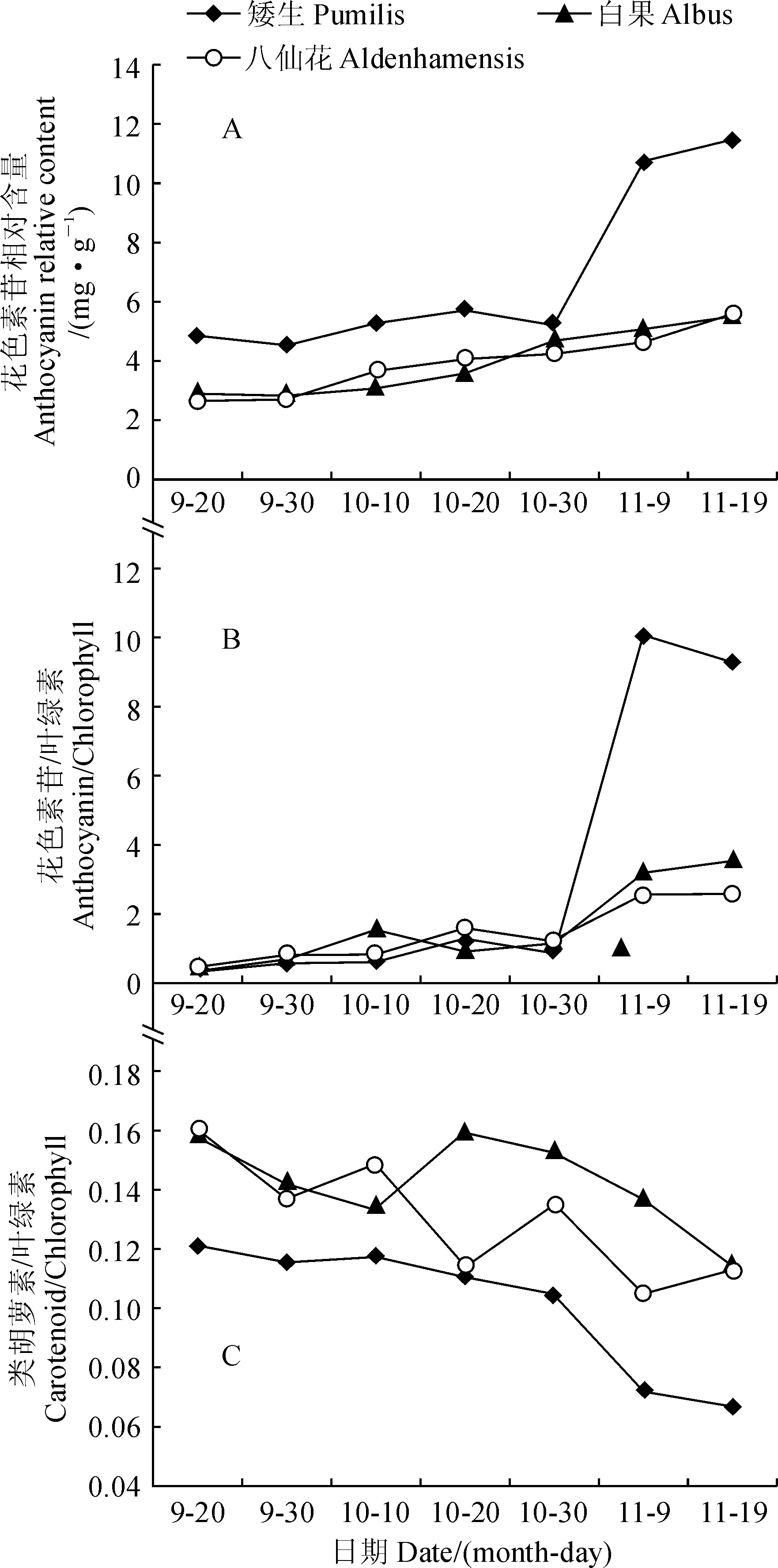
图3 欧洲卫矛叶片花色素苷含量及花色素苷/叶绿素、类胡萝素/叶绿素比值的变化Fig.3 The changes in anthocyanin relative content, anthocyanin/chlorophyll, carotenoid/chlorophyll in leaves of three E. europaea varieties
2.4 欧洲卫矛叶片可溶性糖含量的变化特征
在秋冬叶片转色期,3个欧洲卫矛品种叶片中可溶性糖质量分数呈现先升后降的单峰曲线(图4),这与楚爱香等[19]结果相一致。其中,叶片可溶性糖含量在转色初期增幅明显,推测可能是由于初期气温骤降,诱导可溶性糖含量大幅增加。
在10月20日,各品种欧洲卫矛叶片的可溶性糖质量分数均达到峰值(图4)。相应的‘ 八仙花’、‘白果的’品种叶片花色素苷相对含量在10月20日后上升幅度较之前略大,‘矮生’叶片花色素苷相对含量从10月30日起则急剧上升。可见,各品种欧洲卫矛叶片花色素苷的大量合成出现在其可溶性糖含量达到峰值后,说明可溶性糖的积累可能有利于花色素苷的合成。
2.5 欧洲卫矛品种叶片相关酶活性的变化特征
PAL(苯丙氨酸解氨酶)是花色素苷合成途径中关键性的酶,其活性高低决定花色素苷含量高低,间接决定植物叶色的呈现[20];同时,它在类黄酮类、木质素等的合成过程中也发挥着重要的调节作用[21],能有效抵抗外界环境的干扰。由图5,A可知,3个品种欧洲卫矛叶片PAL活性在转色过程中呈波动上升趋势。其中,‘矮生’和‘白果’的 PAL活性在叶片转色初期增幅明显,转色中期呈下降趋势,转色中后期继续增大,而后保持平稳;而‘八仙花’品种PAL活性在10月20日达到最大值,之后酶活性基本保持平稳。整体上3个品种欧洲卫矛叶片的PAL活性在转色期间均变化不大。
POD和PPO活性与花色素苷的降解有关,花色苷不能直接作为PPO或POD的底物,PPO和 POD 需依赖具邻二酚结构的其他酚类的存在才能氧化、降解花色素苷[4]。由图5,B可知,欧洲卫矛在叶片转色期间POD活性整体呈下降趋势,但3个品种间有所差别,其中,‘矮生’品种POD活性在初期下降明显,中后期基本保持平稳;‘八仙花’、‘白果’品种POD活性在转色前期不断下降,于10月20日后有所上升,最后仍呈下降趋势。同时,由图5,C可知,3个品种叶片的PPO活性在转色期间变化平稳,呈缓慢上升趋势,且3个品种的PPO活性差异不大。
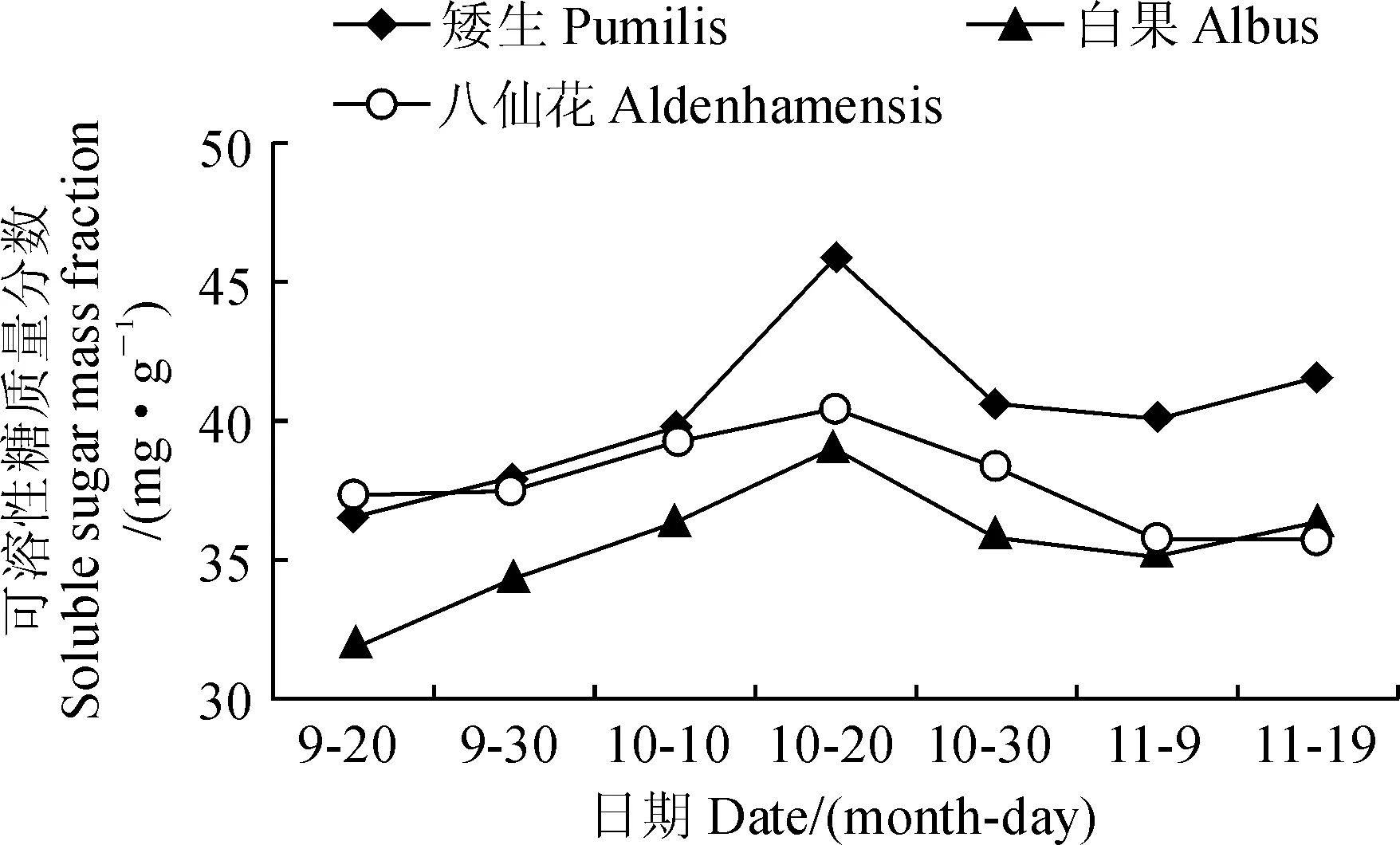
图4 各品种欧洲卫矛叶片可溶性糖质量分数的变化Fig.4 The changes in soluble sugar mass fraction in leaves of three E. europaea varieties
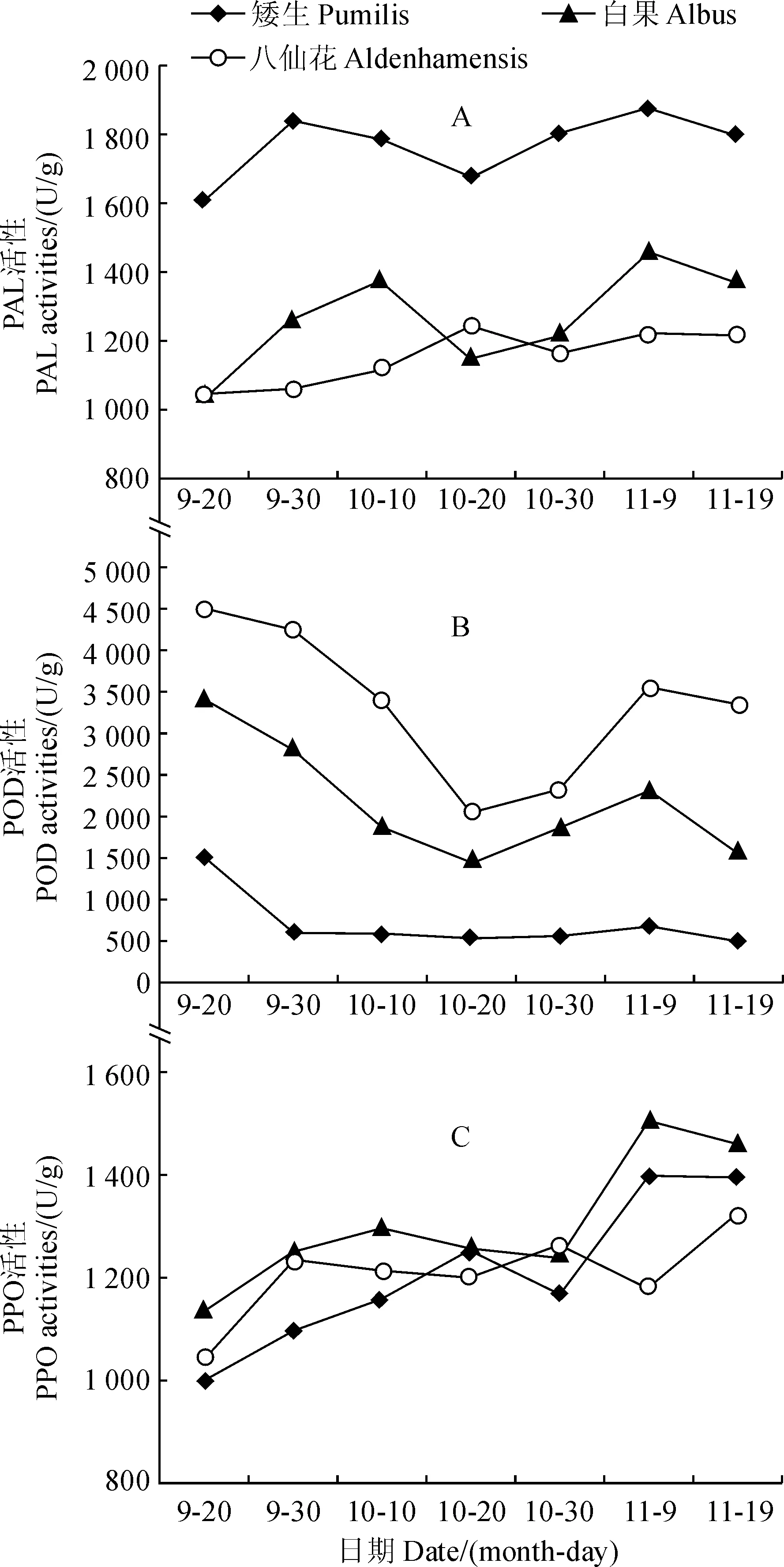
图5 3个欧洲卫矛品种叶片PAL、POD、PPO酶活性的变化Fig.5 The changes in PAL, POD and PPO activities in leaves of three E. europaea varieties
2.6 欧洲卫矛品种叶色参数与色素的相关性分析
由表1可知,各欧洲卫矛品种叶色参数L*、b*值与叶绿素、类胡萝素、花色素苷含量无明显相关性。各品种叶色参数a*值与叶绿素含量均无明显相关性;‘矮生’品种a*值与类胡萝素含量呈极显著负相关,‘八仙花’、‘白果’品种的a*值与类胡萝素无明显相关性;各品种a*值与花色素苷含量均呈极显著正相关。以上结果说明花色素苷是叶色变红的主导因素。
3 讨 论
高等植物叶片中的色素主要是叶绿素(叶绿素a、叶绿素b)、类胡萝素和花色素苷[22]。叶片的呈色与叶片细胞内色素的种类、含量以及在叶片中的分布有关[21]。叶片色素含量的变化和比例直接影响了秋色叶树种秋季叶色的表达。通常植物叶片呈现为绿色是由于叶绿素含量较高而其他的色素含量较低。随着秋季日照时间的缩短与温度的下降,植物叶片开始自然衰老,从而启动叶绿素降解机制,而类胡萝卜素含量相对较稳定,所以秋季叶色多呈黄色。秋季昼夜温差的增大,日照时间的缩短以及温度的下降等环境因素,促进了部分树木花青素的合成和积累,导致其叶片变成红色[23-24]。前人研究结果表明,火炬树[19]、五叶地锦[25]、红枫[26]、落羽杉[27]、榉树[28]等植物秋季叶片中花色素苷相对含量明显增加,在各色素比例中占有绝对优势,是叶片变红的直接原因。本试验结果认为,转色期间温度降低,欧洲卫矛叶片光合作用受阻,使得叶绿素含量不断下降,主要是叶绿素a含量的减少,类胡萝素含量无明显变化,花色素苷含量逐渐升高,且花色素苷/叶绿素的比值不断上升,直接影响欧洲卫矛叶色转红。此外,3个品种叶色参数a*值不断上升,a*值与花色素苷呈极显著正相关,也说明花色素苷显著影响着叶片的变红。
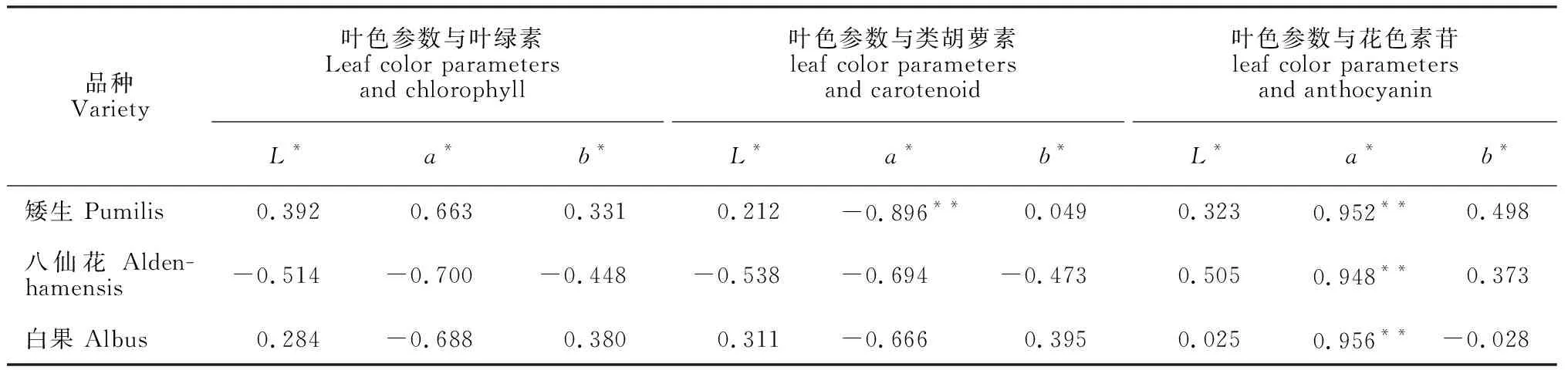
表1 叶色参数与叶绿素、类胡萝素、花色素苷间的相关系数
注:**表示相关系数达到0.01的显著水平Note:**denote correlation coefficients is significant at 0.01 level
研究表明,花色素苷的合成与碳水化合物的代谢有关[29-30],特别是与可溶性糖呈显著正相关[30],可溶性糖还可以作为花色素苷代谢过程中的前体物质或信号分子,促进花色素苷的形成。本试验中,欧洲卫矛各品种花色素苷的大量合成出现在可溶性糖达到峰值后,说明可溶性糖的积累可能有利于花色素苷的合成。转色初期可能由于气温骤降,促使可溶性糖大量积累,后期花色素苷的大量合成消耗部分可溶性糖,致使可溶性糖呈现先升后降的单峰曲线。这与楚爱香等[19, 29]在火炬树、小叶鸡爪槭、三角枫等植物上得到的结果一致。本试验结果同时表明欧洲卫矛3个品种的花色素苷与可溶性糖无显著相关性,姜琳等[31]、许鑫科等[32]分别对栎属植物、紫叶加拿大紫荆的研究也得出了相同的结论。而胡静静[2]、荣立苹等[33]对黄连木、三角枫的研究结果则显示花色素苷与可溶性糖呈显著正相关。由此可知可溶性糖对于花色素苷合成的调控较为复杂,可溶性糖对于花色素苷的合成可能并不是决定性的因素,今后可以在分子水平对于欧洲卫矛可溶性糖是否作为信号分子方面进行进一步的探究。
花色素苷的合成是在一系列酶的催化下形成的,而苯丙氨酸解氨酶(PAL)是花青素合成的第一个关键酶[3]。前人对红栌[3]、红花檵木[34]、鹅耳枥[35]、紫叶稠李[36]等植物研究发现,PAL活性与花色素苷含量呈正相关性。而Lister等[14]、王惠聪等[37]发现PAL活性与荔枝果皮中的花色素苷合成无关。本试验中,欧洲卫矛3个品种的PAL活性在转色期间缓慢波动上升,花色素苷的含量也在上升;相关分析表明‘八仙花’品种PAL活性与花色素苷含量呈显著正相关,‘矮生’、‘白果’品种PAL活性与花色素苷含量呈中等水平正相关,但未达到显著水平,可见PAL活性升高有利于欧洲卫矛叶片变红。
研究表明,POD在过氧化氢(H2O2) 存在时能氧化多种酚类成褐色产物[38],促进叶片呈色,因此POD活性越高,叶片红色素积累越多[3, 25]。吴驭帆等[35]研究发现鹅耳枥花色素苷含量与POD活性呈极显著正相关,且红叶植株的花色素苷含量、POD活性高于绿叶植株,认为POD可能参与并促进了叶片花色素苷的积累,进而影响鹅耳枥叶片的呈色。而在本试验中欧洲卫矛3个品种的POD活性在叶片转色期间呈下降趋势,且初期下降明显,转色后期略有上升,后趋于平稳;相关分析表明,POD活性与花色素苷含量呈负相关,但不显著,推测POD参与了花色素苷的降解,但影响不大。相反,POD活性与叶绿素含量在转色期间均呈下降趋势,推测POD活性与叶绿素含量有一定的关系,这与文国琴等[39]、李力等[40]的研究结果相似,认为POD在叶片转色期间,不仅影响叶片中多酚含量的变化,同时参与了叶绿素的降解。另外,张元慧等[41]对李果皮研究发现,PPO活性与花色素苷含量具有明显的正相关,认为PPO可能将果皮中的酚类物质氧化成醌,促进了果皮呈紫色。本试验中,在叶片转色期间,欧洲卫矛3个品种的PPO活性呈缓慢上升趋势,‘矮生’、‘白果’PPO与花色素苷含量呈显著正相关,‘八仙花’PPO活性与花色素苷含量呈不显著正相关,这与张元慧的研究结果相似。说明PPO有利于欧洲卫矛叶片花色素苷的合成与积累。
综上所述,通过对3个欧洲卫矛品种在秋冬转色期间的叶色参数及各项生理指标含量的比较分析,欧洲卫矛秋季叶色转红主要是由于花色素苷相对含量大量增加,在色素中所占比值占优势。可溶性糖有利于花色素苷在转色后期的大量合成。PAL、PPO有利于花色素苷的合成与积累,POD与花色素苷无显著相关性,可能与叶绿素的降解存在一定的关系。今后可在可溶性糖及各类酶的分子水平上进一步探讨欧洲卫矛叶片呈色机理。
