重庆大巴山国家级自然保护区森林植物多样性垂直格局
2018-07-19倪东萍邓洪平李宗峰左有为李满婷
倪东萍,邓洪平,顾 梨,何 松,李宗峰,左有为,李满婷
(西南大学 生命科学学院,三峡库区生态环境教育部重点实验室,重庆400715)
物种多样性的分布格局会受到外界环境不同程度的影响[1],其对环境条件的响应是植物生态学研究的热点之一[2-3]。海拔梯度由于包含了多种环境因子的梯度效应,被认为是影响物种多样性格局的决定性因素之一[4-5]。随着海拔升高,在温度、光照、水分和土壤等生态因子的综合作用下,生物群落表现出明显的垂直地带性分布规律[6]。且沿海拔梯度升高,水热组合发生变化,地带性植被以及群落内的建群种和优势种亦将发生改变[7]。
重庆大巴山国家级自然保护区由于区内秦岭山脉阻挡了第四纪冰川对保护区的影响,使之成为第四纪冰期许多生物的“避难所”,保留了大量珍稀濒危及孑遗物种[8-9]。截至目前,大巴山已被《中国生物多样性国情研究报告》、世界自然基金会(WWF)等列入中国生物多样性保护的关键地区和热点生态区,被《中国生物多样性保护行动计划(2011~2030年)》列入中国35个生物多样性优先保护区域之一,具有很高的科学研究和保护价值[10]。
许冬焱等先后对大巴山自然保护区内种子植物区系组成及其植被特征进行了初步研究[11-12],邓洪平等对保护区内植被垂直带谱、群落类型和植物区系特征进行了分析[9],但对其海拔梯度上植物多样性及成分结构的数量变化尚无深入了解。因此,本研究以大巴山保护区海拔梯度上的植物群落调查为基础,对区内森林植物多样性垂直格局进行研究,以期为进一步研究群落的稳定性、恢复机制、演替过程以及群落的结构和功能提供生态学依据,从而为大巴山生物多样性的保护和管理提供科学依据。
1 研究区概况
重庆大巴山国家级自然保护区(N31°37′27″~32°12′15″,E108°27′07″~109°16′40″)位于重庆市东北部的城口县境内[8]。地质构造上属大巴山弧形断褶带的南缘部份,由一系列北西至东西走向的雁列式褶皱和冲断层组成。地貌上属米仓山、大巴山中山区,山脉受地质构造和岩性的控制,诸列山岭均由北西向南东展布。区内最高点光头山,海拔2 685.7 m,最低点龙田乡田湾,海拔754.0 m。整个地势南东偏高,北西偏低。该区属亚热带季风气候,由于山高谷深,高差大,具有山区立体气候的特征。区内年均气温13.8 ℃,年均降水量1 261.4 mm,年均日照时数1 534 h,年无霜期234 d,年平均相对湿度78%。保护区为森林生态系统类型保护区,主要保护对象为亚热带森林生态系统及其生物多样性。
2 研究方法
2.1 调查地点的选择与样地设置
为全面反映保护区内森林植物多样性的海拔梯度格局,根据保护区内森林分布现状,参照国家环境保护部生物多样性监测标准,在涉及保护区的13个乡镇(左岚乡、高楠镇、龙田乡、北屏乡、岚天乡、河鱼乡、东安镇、明中乡、高观镇、巴山镇、厚坪乡、蓼子乡、咸宜镇)设置20个样地(表1),形成了一条较为整齐的样带。样带最低点842.0 m,最高点2 217.2 m,以西南为主坡向,坡度范围15°~64°。
在20个样地内,采用相邻格子法和十字分割法,对不同群落类型分别进行典型样方取样,群落层次按乔木层、灌木层和草本层划分。(1)设置20 m×30 m的样方,以10 m×10 m为单元进行每个胸径(DBH)≥5 cm的乔木物种调查;(2)每个10 m×10 m的格子中分别划出1个5 m×5 m的样方作灌木层物种调查;(3)在每个灌木层样方内,设置2个2 m×2 m的样方作草本物种调查。调查记录的指标有调查地点、地形、样地面积、群落类型、森林起源、郁闭度、种名、胸径、树高、株数、盖度、多度等,同时记录各样方的经纬度、海拔、坡向、坡度和坡位等因子。对所有样地采用分时段重复调查方法,自2016~2018年间重复调查4次,共调查乔木样方120个,灌木样方120个,草本样方240个。
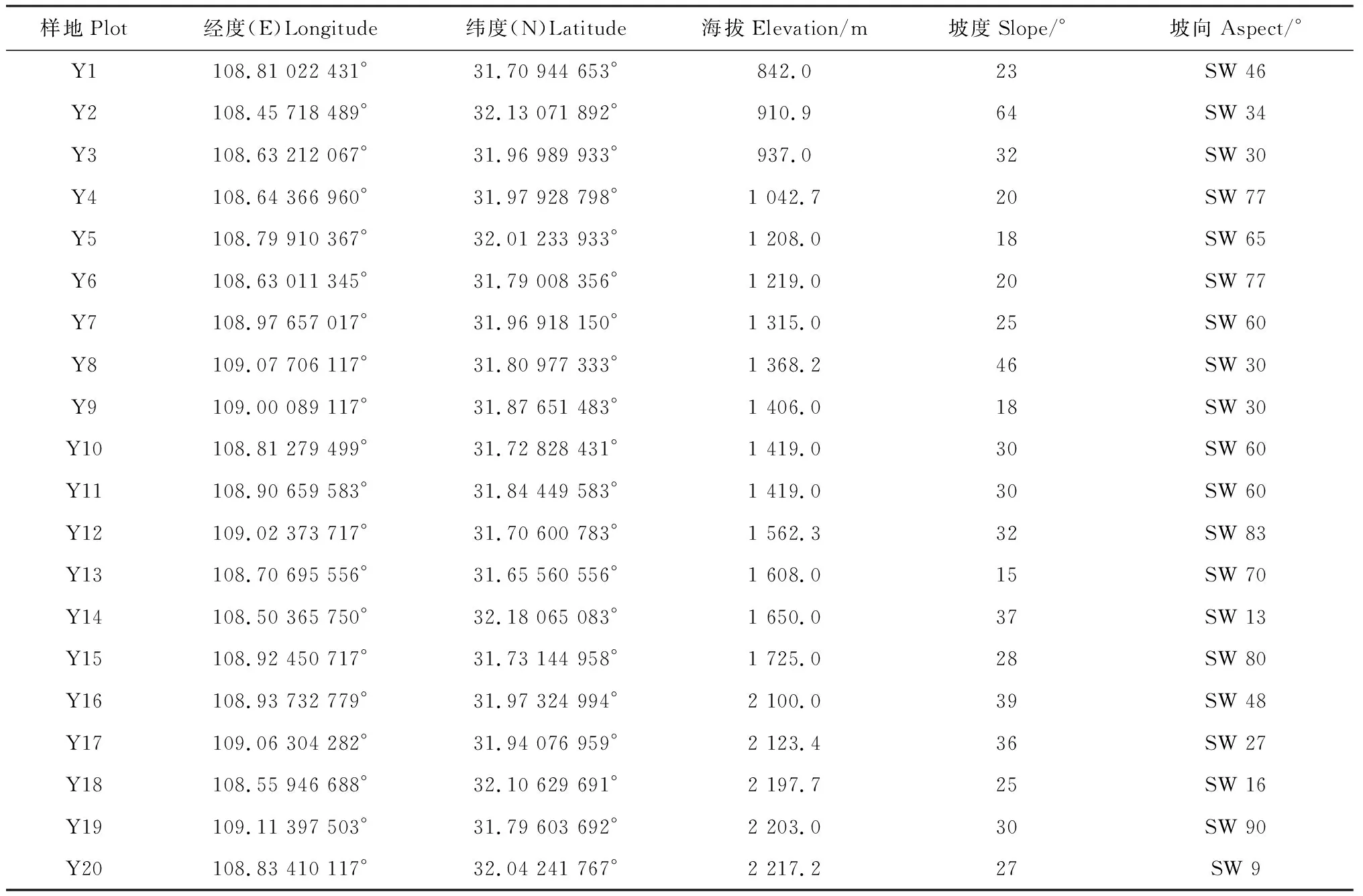
表1 样地概况
2.2 数据处理与计算方法
2.2.1物种丰富度及其结构成分(1)物种丰富度。统计各样方中维管植物的科(F)、属(G)、种数(S),计算各样方内种数/属数(S/G)、种数/科数(S/F)、属数/科数(G/F)。
(2)生态成分。根据乔木层物种的生长型划分不同的生态成分,并计算其比值。
2.2.2群落数量分类采用等级聚类分析(Hierarchical Cluster Analysis)方法[13],依据乔木层优势种重要值对样方进行聚类分析。重要值计算方法为:
IV= (A+D+F)/3
(1)
其中,A为相对多度,即每个种的多度占所有种多度之和的百分比;D为相对优势度,即每个种的胸高断面积之和占所有种个体胸高断面积之和的百分比;F为相对频度,即每个种的频度占所有种频度之和的百分比。
2.2.3多样性测度(1)α多样性测度。本研究选择Shannon-Wiener多样性指数(H)、Pielou均匀度指数(J)和Simpson优势度指数(D)研究物种多样性[7],计算公式如下:
H=∑PilnPi
(2)
J=(-∑PilnPi)/lnS
(3)
D=∑Pi2
(4)
其中,Pi为该物种i的简化重要值,S为物种总数。
(2)β多样性测度。
Cody指数[14]:
βc =[g(H)+l(H)]/2 =(a+b-2c)/2
(5)
其中a、b分别为两群落的物种数,c为两群落的共有物种数,g(H)为沿生境梯度H增加的物种数,l(H)为沿生境梯度H失去的物种数。
所有统计分析均在R软件(R Development CoreTeam,2009)中进行。
3 结果分析
3.1 物种组成与分类
3.1.1群落各层种类组成及沿海拔的变化规律重复调查的20个森林群落样地中共记录到维管植物97科226属335种,其中蕨类10科15属19种,裸子植物4科9属13种,被子植物83科202属303种。
调查样地中重要值≥0.1的物种共有141种,占总种数的42%;属于单种的科有3种,占总科数的3.1%,单属的科12种,占总科数的12.5%;种数大于10的科有蔷薇科(Rosaceae)、菊科(Compositae)、壳斗科(Fagaceae)、 百合科(Liliaceae)、豆科(Leguminosae)5个科。种数最多的科为蔷薇科(31种)。
20个森林群落样地中共记录乔木树种94种,隶属于32科53属,其中有落叶阔叶树种54 种,常绿阔叶树种27种,常绿针叶树种13种,未发现落叶针叶树种。落叶树种和常绿树种分别占乔木树种的57%和43%,灌木物种100种,隶属于37科67 属,草本物种141种,隶属于47科109属。
群落各层次优势种如表2所示,综合分析结果为:沟谷浅丘中乔木层优势种多以壳斗科植物及马尾松(Pinusmassoniana)、杉木(Cunninghamialanceolata)等偏暖性针叶树种为主,壳斗科植物常见的如栲树(Castanopsisfargesii) 、青冈(Cyclobalanopsisglauca)、巴东栎(Quercusengleriana)。随着海拔上升,陆续出现落叶阔叶树种,与常绿阔叶树种形成混交局面。高海拔地段则以偏湿性针叶树种,如华山松(Pinusarmandi)、巴山松(Pinushenryi),及偏寒性针叶树种巴山冷杉(Abiesfargesii)、青杄(Piceawilsonii)为主。灌木层主要优势种随海拔上升主要有盐肤木(Rhuschinensis)、棣棠(Kerriajaponica)、三桠乌药(Linderaobtusiloba)、小果蔷薇(Rosacymosa)、箭竹(Fargesiaspathacea)等,蔷薇科中灌木型植物在各海拔区段均有分布。草本层优势种主要为菊科植物,如橐吾(Ligulariasibirica)、白酒草(Conyzajaponica),蕨类植物在低海拔区段略占优势,高海拔区段莎草科(Cyperaceae)苔草属(Carex)植物相对丰富。
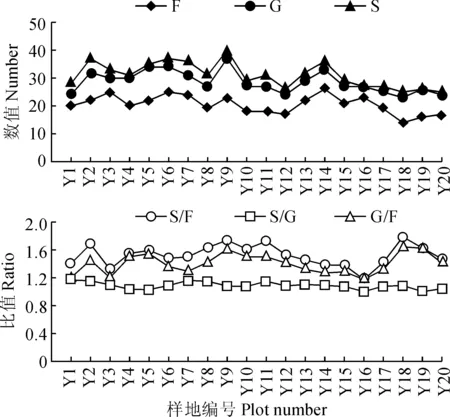
F.科数;G.属数;S.种数;S/F.种数/科数;S/G.种数/属数;G/F.属数/科数图1 不同系统等级分类单元的多样性梯度F. Family; G. Genus; S. Species; S/F. Species/Family; S/G. Species/Genus; G/F. Genus/FamilyFig.1 Altitudinal gradient of diversity of units at different taxonomic levels
3.1.2物种丰富度不同分类等级之间的数量比(图1) 基本反映了不同林型的物种丰富度沿海拔梯度的变化规律。20个森林群落样地中,科(F)、属(G)、种(S)总体上随着海拔的升高呈先增加后降低的趋势,在Y10~Y12和Y15~Y20区段有较小值,在Y9和Y14段出现较高峰值。初步推测,这些变化趋势与该样地林型分布范围相关。
S/F、S/G、G/F的比值是对植物区系分化强度的反映,其值越高,则表示属、科的丰富度越低,区系的分化程度越低[15],20个样地的计算结果表明:S/F、G/F有相同的变化趋势,在高海拔区段(Y17、Y18、Y19、Y20)出现较大值,表明该区域区系分化程度相对较低。S/G的值变化不大,说明种的多样性变化时,属的多样性随之变化。
3.1.3群落生态成分图2反映了乔木层常绿阔叶、落叶阔叶和常绿针叶3种类型树种的重要值百分比沿海拔梯度的变化,整个调查样地中未发现落叶针叶树种的存在。在海拔1 000 m以下(Y1-Y3)的样地中,常绿树种的重要值为32%~100%,其中偏暖性常绿针叶树种的重要值为20%~35%,说明该海拔区段包含常绿阔叶林和偏暖性针阔混交林两个林带;海拔1 000~1 600 m (Y4~Y12)段,常绿阔叶和落叶阔叶树种的重要值百分比值较高,仅有少量针叶树种出现;海拔1 600 m~2 100 m (Y13~Y16)段,偏湿性针叶树种,如巴山松、华山松等的比例增大,落叶阔叶树种的重要值迅速降低,有少部分常绿大灌木或小乔木重新出现;海拔2 100 m(Y17~Y20)以上,偏寒性暗针叶林占主导地位,期间出现少量红桦、锐齿槲栎等落叶树种。
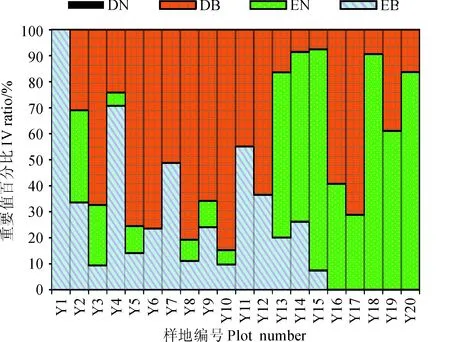
DN.落叶针叶;DB.落叶阔叶;EN.常绿针叶;EB.常绿阔叶图2 乔木层不同生长型植物重要值百分比沿海拔梯度的变化DN. Deciduous needles;DB. Deciduous broadleaf;EN. Evergreen needles;EB. Evergreen broadleafFig.2 Altitudinal patterns of Ⅳ percentage in different life forms of tree layer

3.1.4群落分类等级聚类分析结果(图3)表明,20个样地被明显地分为了4个类型:第Ⅰ类,以常绿树种和偏暖性针叶树种(马尾松、杉木等)为主要优势种的群落,包括 Y1~Y3号样地。第Ⅱ类,以常绿落叶阔叶树种为主要优势种的群落,包括Y4~Y12号样地。第Ⅲ类,以巴山松和华山松为主要优势种的群落,包括Y13~Y15号样地。第Ⅳ类,以巴山冷杉和青杄为主要优势种的群落,包括Y16~Y20号样地。数量分类的结果与样地植被类型的分布格局基本相符,印证了分类结果的合理性。
3.2 群落多样性
3.2.1α多样性随海拔的变化为克服不同层次因面积不同造成的物种数量上的差异,对样地分乔木层、灌木层和草本层3个层次进行物种的Simpson 指数、 Shannon-Wiener 指数、Pielou均匀度指数统计(图4)。结果表明:样地α多样性的垂直梯度变化规律较为复杂,Simpson优势度和Pielou均匀度在-个层次中随海拔梯度的变化较小,说明物种均匀分布在调查区域内,形成最佳搭配模式。乔木层中Shannon-Wiener指数呈现出明显的随海拔升高而逐渐降低的规律,这与被广泛接受的α多样性随海拔升高而降低的格局相吻合[16]。灌木层和草本层的Shannon-Wiener指数曲线在中高海拔区段相对平稳。相对乔木层而言,两者与海拔梯度未呈现明显的变化规律。
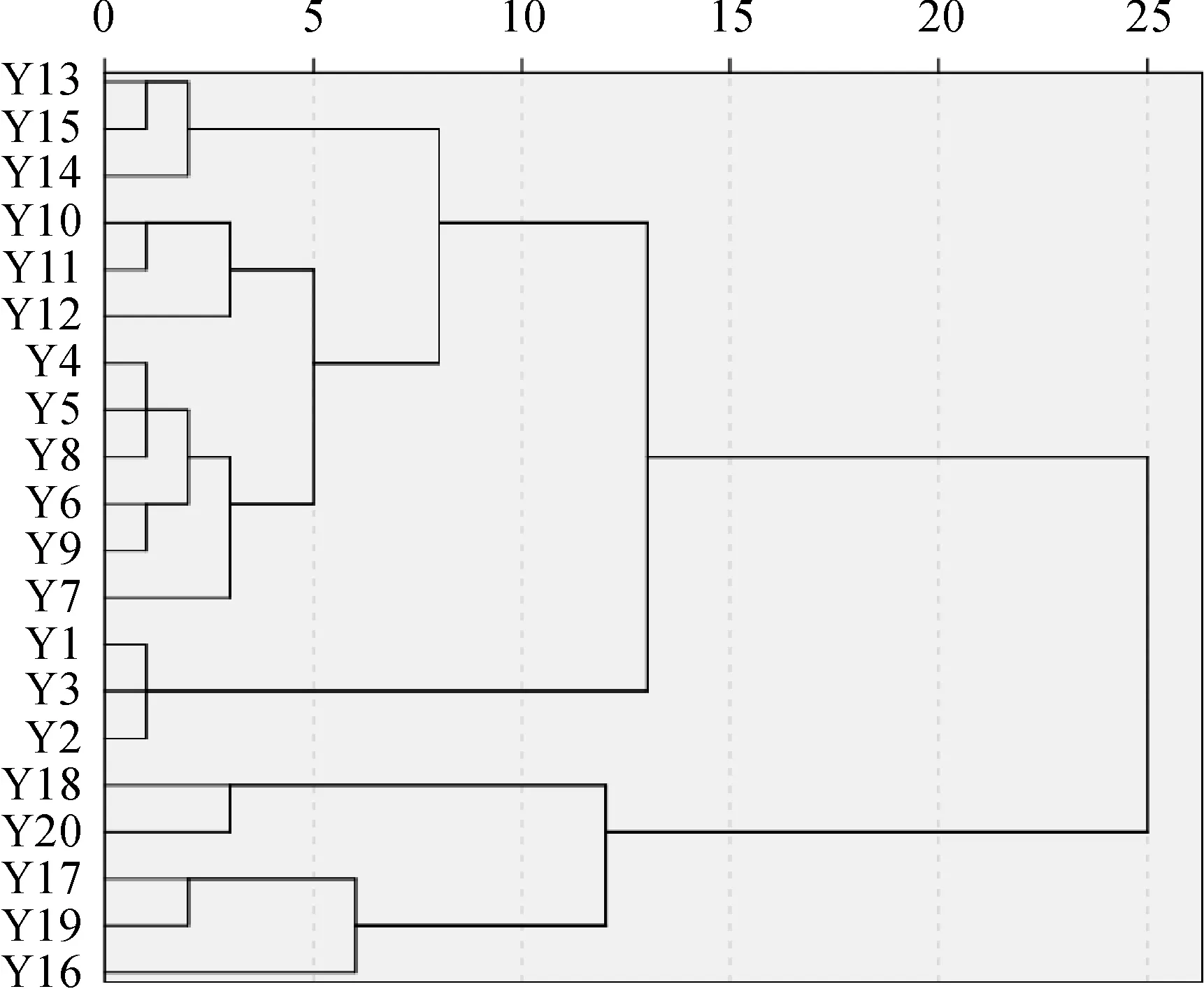
Y1~Y20同表1图3 群落样方的聚类分析The meaning of Y1-Y20 is shown in Table 1Fig.3 Cluster analysis classification of the community
3.2.2β多样性随海拔的变化β多样性指数的曲线图反映了不同海拔段物种交替的速度,图5为沿海拔梯度相邻样地间的β多样性变化趋势图,结果表明:相邻样地乔木层、灌木层、草本层间种类组成随海拔梯度变化差异比较明显。乔木层的β多样性指数的曲线变化幅度较小,Cody指数在Y2~Y3、Y12~Y13中出现较大值,结合图2分析,这些区段属于不同类型混交林过渡区域,相邻两个群落的生境差异较大,物种异质性相对较高,物种交替速度快。
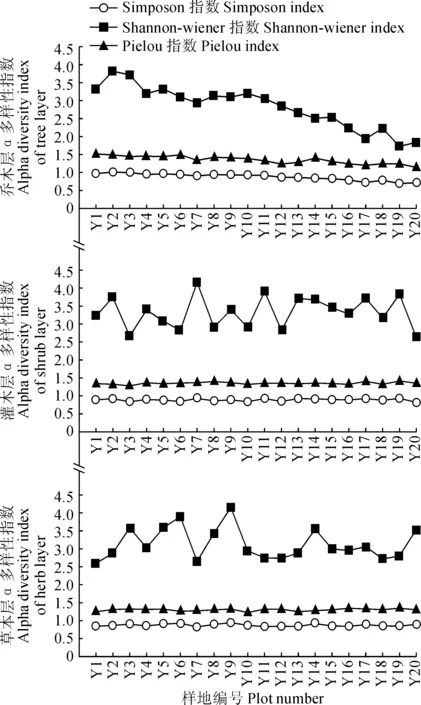
图4 α多样性指数随海拔梯度变化趋势Fig.4 α species diversity along an altitudinal gradient

图5 相邻样地β多样性Fig.5 Beta diversity of each two adjacent quadrats
高海拔区段属于暗针叶林区域,相邻两个群落的生境比较接近,乔木层Cody指数出现最低值。灌木层Cody指数曲线变化幅度较大,Y1~Y2、Y7~Y8的Cody指数值明显高于其他群落。相邻样地间草本层的Cody指数值与灌木层指数值大体呈负相关,在中海拔区段有较低值。以上所有分析的结果与α多样性的结果相一致。
4 讨 论
由于低山地段受水分限制,高海拔地区受热量限制[17],多数植物多样性垂直格局研究结果符合“中间高度膨胀”理论,即物种多样性在中间海拔地区最高[18]。但在本研究中,重庆大巴山国家级自然保护区森林植物多样性格局呈现出较为复杂的变化规律, “中间高度膨胀”多样性理论表现不明显,这可能与样地选址的海拔范围没有完全涵盖整个保护区的海拔范围有关。乔木层中Shannon-Wiener指数呈现出明显的随海拔升高而逐渐降低的规律,原因可能是随着海拔升高,针叶树种大量出现。由于针叶林存在枯落物难分解、自肥能力差等缺陷,其涵养水源和保持水土的能力相对较差,加上高海拔区段生境的异质性和特殊性,仅有特定物种适合生存,从而导致物种的多样性降低。就林下物种多样性而言,研究发现,不同森林群落林下灌木层和草本层具有显著差异。落叶阔叶树种较多的低中海拔区段,由于林地郁闭度大、林下落叶积累较多等原因,不利于林下物种的更新和生长,从而导致灌木层物种多样性较低。与此同时,分布在低海拔区段的林下草本,由于人为活动的干扰,一定程度上增加了种子的传播几率,加上该区段喜阴草本较多,导致物种多样性相对较高。
通过植被类型间的比较表明,无论是群落中植物科、属、种数,还是物种多样性及区系分化强度,混交林类型的值均比较大,其结果与沈泽昊等研究的神农架南坡植物群落多样性的海拔梯度格局相符[19]。形成原因可能有以下几个方面:首先,区系的过渡性导致不同林型群落的物种多样性和区系分化强度产生差异;其次,α多样性会受限于生境异质性的水平,混交林型群落的冠层结构复杂,林下光照的异质性水平高于其他林型。最后,多样性数值较低的海拔段的优势种可能为成立保护区前栽种的人工林,其物种组成相对单一。
β多样性,表示在一个梯度上从一个生境到另一个生境所发生的种的多样性变化速率和范围[13],Cody 指数作为其中一个指数,通过对新增加和失去的物种数目进行比较,从而获得十分直观的物种更替概念,能清楚地表明多样性含义[14]。本研究中,低海拔区段3个层次的β多样性指数曲线均具有波动性,表明群落间的物种更替速率在低海拔区段无明显的递变规律。乔木层的波动幅度相对较小,其Cody指数峰值点很好地反映了植被带的分界线。随着海拔的上升,群落间的生境差异性减小,海拔梯度上热量的递减导致物种丰富度减少,高海拔地区的植物分布范围较广,进一步引起群落间物种交换量(Cody指数)减少,植物群落间的物种更替速率降低,并达到相对稳定的状态,该β多样性变化格局符合随海拔升高而减小的常见格局[7,20]。草本层的β多样性格局相对复杂,很大程度上与草本植物易受群落盖度、光照和微地形等因素的影响有关[16]。
综合群落生长型和等级聚类分析结果,大巴山森林植物群落的20个样地被分为4个类型:第一类是在海拔1 000 m以下的沟谷地带,光照时间较短,湿度条件较好,包含常绿阔叶林和偏暖性针阔混交林两个林带;第二类,海拔1 000~1 600 m的低中山地带,因太阳直射时间较长,湿度条件不如沟谷地带,属常绿、落叶阔叶混交林带;第三类是海拔1 600 m~2 100 m的中山地段,属偏暖湿性针叶阔叶林带;第四类,是海拔2 100 m以上的亚高山地带,环境条件相对恶劣,群落主要优势种为巴山冷杉和青杄等植物,属偏寒性暗针叶林带。因此,重庆大巴山自然保护区植被在保护和可持续发展时,要根据植物群落物种丰富度的海拔梯度变化规律,科学、合理地进行立地类型规划,制订相应技术和管理措施,开展高效的植物群落营建和封育管护,才能达到费省效宏、持续久远的效果。
