基于SSR分子标记的延胡索遗传多样性研究
2018-07-19周天华张智强
徐 皓,周天华,张智强,白 桥
(1 陕西理工大学 生物科学与工程学院,陕西汉中 723001;2 陕南秦巴山区资源生物综合开发协同中心,陕西汉中 723001)
延胡索又名元胡,为罂粟科(Papaveraceae)紫堇属(Corydalis)植物延胡索(CorydalisyanhusuoW. T. Wang)的干燥块茎,性味辛温,味苦,归肝、脾经,能行血中气滞,具有理气止痛,活血化瘀的功效[1-2]。临床常用于胸肋、脘腹疼痛、经闭痛经、产后瘀阻、跌打肿痛等症[3]。
延胡索主要分布于浙江、江苏、安徽、陕西等地,以浙江东阳、磐安、缙云和永康等地及江苏南通地区栽培面积最大。近年来,陕西汉中的种植面积由2008年4 355 hm2已发展到2017年的6 700多hm2,面积和产量占到全国的75%左右,已经成为汉中市中药材产业的支柱品种之一。
简单重复序列(SSR,simple sequence repeat)是一种以特异性引物PCR为基础的标记方法,也叫微卫星DNA,通常以1~6个核苷酸为重复单位串联成长达几十个核苷酸的重复序列[4]。1970年左右,人们就已经发现在真核生物基因组中普遍含有简单的重复序列,且不同物种重复单位数和重复序列存在丰富的差异。重复序列的两端一般情况下是趋于保守的单拷贝序列,因此可设计特异性引物来进行 PCR 扩增,该方法同时具备RAPD和RFLP技术的优点,而且克服了它们的不足,因而成为目前分子标记的热点,广泛应用于遗传图谱构建、品种指纹图谱绘制、品种纯度检测及目标性状分子标记筛选等领域[5]。但迄今尚没有利用微卫星分子标记技术对延胡索进行遗传多样性分析的相关报道。
本研究首次应用SSR分子标记技术,对19个居群的360份延胡索样本进行了分析,旨在揭示其遗传多样性及遗传结构,并据此了解其进化潜力和对环境的适应能力,为科学保护和利用延胡索植物资源提供理论依据和技术支持。
1 材料和方法
1.1 实验材料
试验采集了延胡索19个居群,360份样本的新鲜叶片,放入自封袋中加硅胶干燥,带回实验室保存在-80℃超低温冰箱中保存备用,其中汉中城固、汉中南郑和浙江东和的延胡索材料为栽培种,其余的材料均为野生种,居群采样基本数据见表1。
1.2 方 法
1.2.1延胡索DNA的提取采用改良的CTAB法提取24份样品中的基因DNA,使用紫外分光光度计检测DNA浓度和A260/A280吸光比值,并用浓度为1%琼脂糖凝胶检测DNA完整性,凝胶成像仪拍照观察并记录结果。
1.2.2PCR扩增利用开发及筛选的12对引物对24份供试样品的DNA进行扩增。PCR总反应体系为10 μL,DNA 模板1 μL,Mix 5 μL,正反引物各1 μL,ddH2O 2 μL。PCR反应程序:首先94 ℃预变性3 min;94 ℃变性30 s,退火温度下45 s,72 ℃延伸 45 s,共35个循环;最后72 ℃延伸7 min,4 ℃保存。
1.2.3聚丙烯酰氨凝胶电泳检测完成PCR扩增后24 h内,取2.5 μL PCR产物在10%非变性聚丙烯酰胺凝胶上恒电压140~160 V电泳10~15 h(依据目标条带大小而异);用0.2%NaOH银染30 min,用去离子水漂洗3次后,用3%AgNO3进行银染;去离子水漂洗后,用凝胶成像系统采集图像。
1.2.4延胡索遗传多样性分析使用Excel 2007和Bio-rad quantity 软件对电泳后的凝胶图像采用人工读带方式进行条带统计。应用GenALEx 6.3软件进行各居群地理距离和遗传距离相关性分析,用以检测居群间的相互关系。使用POPGENE Version 1.32 软件,分析各引物的等位基因数(Na)、遗传杂合度(He)、Nei’s遗传多样性指数(H)、遗传分化系数(Fst)和基因流(Nm),计算各引物的遗传多样性和近交指数(Fis);还采用该软件计算了各种群的地理距离和遗传距离,并做了两者相关性检验(Mantel),采用MEGA5.0软件基于各居群的遗传距离进行UPGMA聚类分析。此外,为分析延胡索的遗传关系,应用Structrue V2.3.4[6]软件来确定所有样本的聚类分组,并构建了各样本的遗传关系图,使用Arlqequin3.0计算变型间与变型内的遗传变异情况。
2 结果与分析
2.1 DNA提取与引物扩增效果
紫外分光光度计检测结果显示,提取到的延胡索叶片DNA浓度均在100 ng/μL之间,A260/A280吸光比值均在1.8~1.9之间,基本上没有蛋白质杂质污染。琼脂糖凝胶电泳显示,提取到的延胡索DNA长度均大于2 000 bp,能够用于扩增及检测。12对SSR引物扩增结果显示,所有样品均成功扩增出条带(图1),具有较高的稳定性和多态性。
2.2 延胡索遗传多样性与遗传分化
表2和表3为12对SSR引物和19个居群360份样本进行遗传多样性分析,得到的等位基因数、观察杂合度等遗传多样性与遗传分化指数信息。表3显示12对SSR引物中检测到4~9个等位基因,平均等位基因数目为5.25个,这些微卫星引物的观察杂合度为0.178 9~0.465 8,平均值为0.335 5;期望杂合度为0.413 8~0.785 2,平均值为0.635 1。表2显示观测等位基因数总和为43.826 1,有效等位基因数总和33.217 7;Shannon信息指数范围为0.520 1~0.775 0,平均值为0.613 2;遗传多样性指数范围为0.340 2~0.470 0,平均值为0.387 9。一般近交系数(Fis)值越接近于零,说明基因型分布越接近于平衡状态,Fis值为正说明杂合子缺失,Fis值为负说明杂合子过剩。表3显示,Fis数值有正有负,最大为0.454 9,最小为-0.245 3,平均为0.135 1,表明延胡索居群整体基因型表现为杂合子缺失;遗传分化系数(Fst)值平均为0.388 3,大于0.25,说明有38.83%的遗传变异存在于群体间,绝大部分变异(61.17%)位于群体内,群体间有很大的遗传分化。
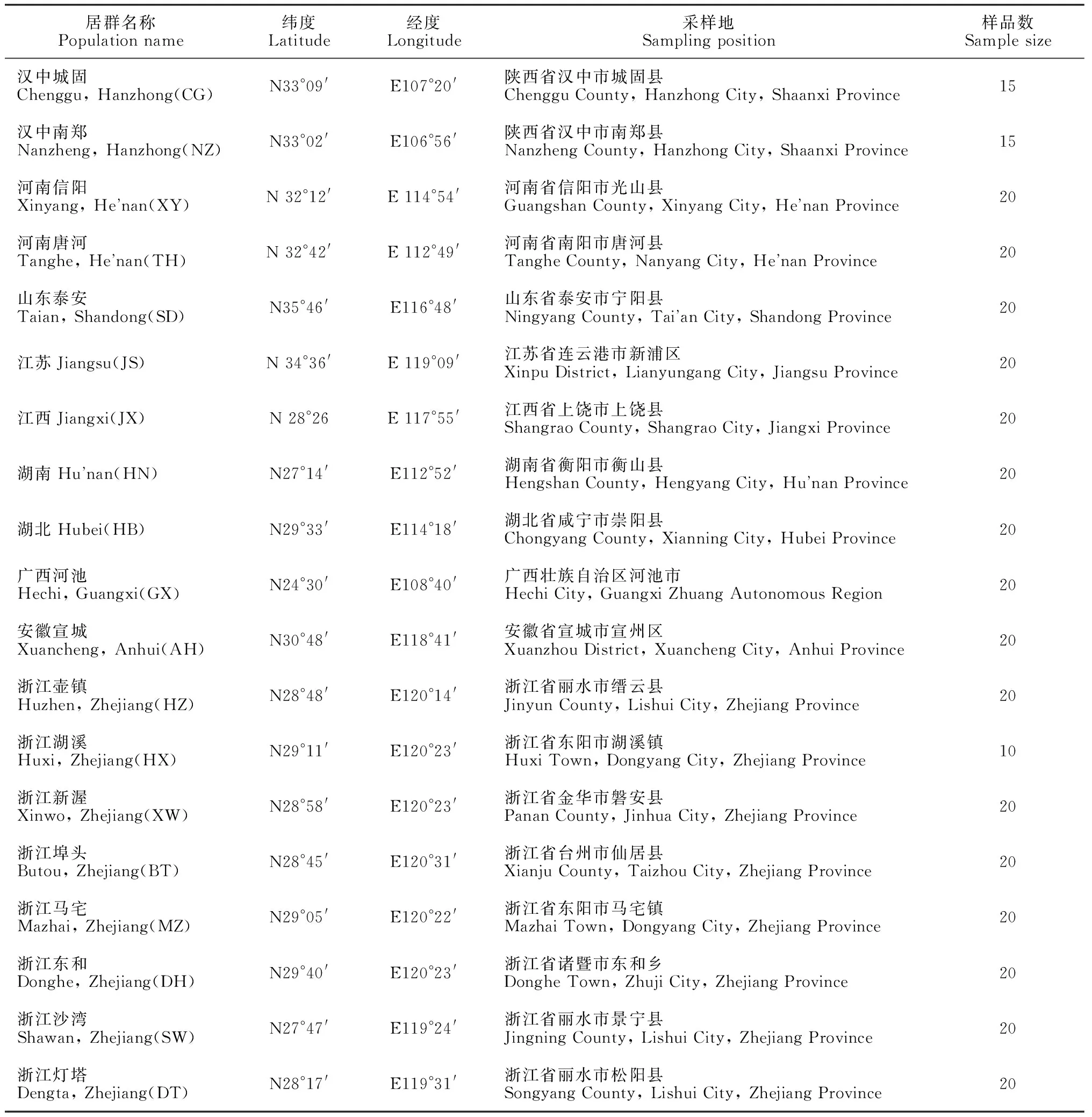
表1 延胡索19个居群采样基本信息
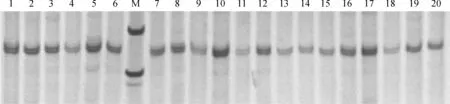
M.PUC-18 DNA marke; 1~20.汉中城固(CG)居群的20个个体图1 引物 Cyuan15 对城固(CG)居群的扩增效果图M. PUC-18 DNA marker; 1-20. 20 samples of population CGFig.1 Amplification pattern of population CG using the primer pair Cyuan15
2.3 居群主相关性分析和聚类分析
基于居群遗传距离进行UPGMA法聚类分析(图2)。可以看出延胡索19个居群明显分为4个组:采自陕西但引种自江西的两个种群(CG和NZ)与华东的3个居群(JX、JS、BT)聚为一小支,采自汉中但引种自浙江的居群(HZ)与浙江的3个居群(DT、SW、XW)聚为一小支,然后这两个小支组成一支;华北3个居群(XY、TH、SD)聚为一支;采集自华东的AH、DH、HX和MZ聚为一支;华中与华南3个居群(HB、HN、GX)聚为一大支。聚类分析的结果表明,当ΔK=4时,K值最大(图3),说明19个延胡索居群分为4支较为合理;此时,19个居群的聚类结果与UPGMA树完全一致。

表2 19个居群的延胡索遗传多样性分析
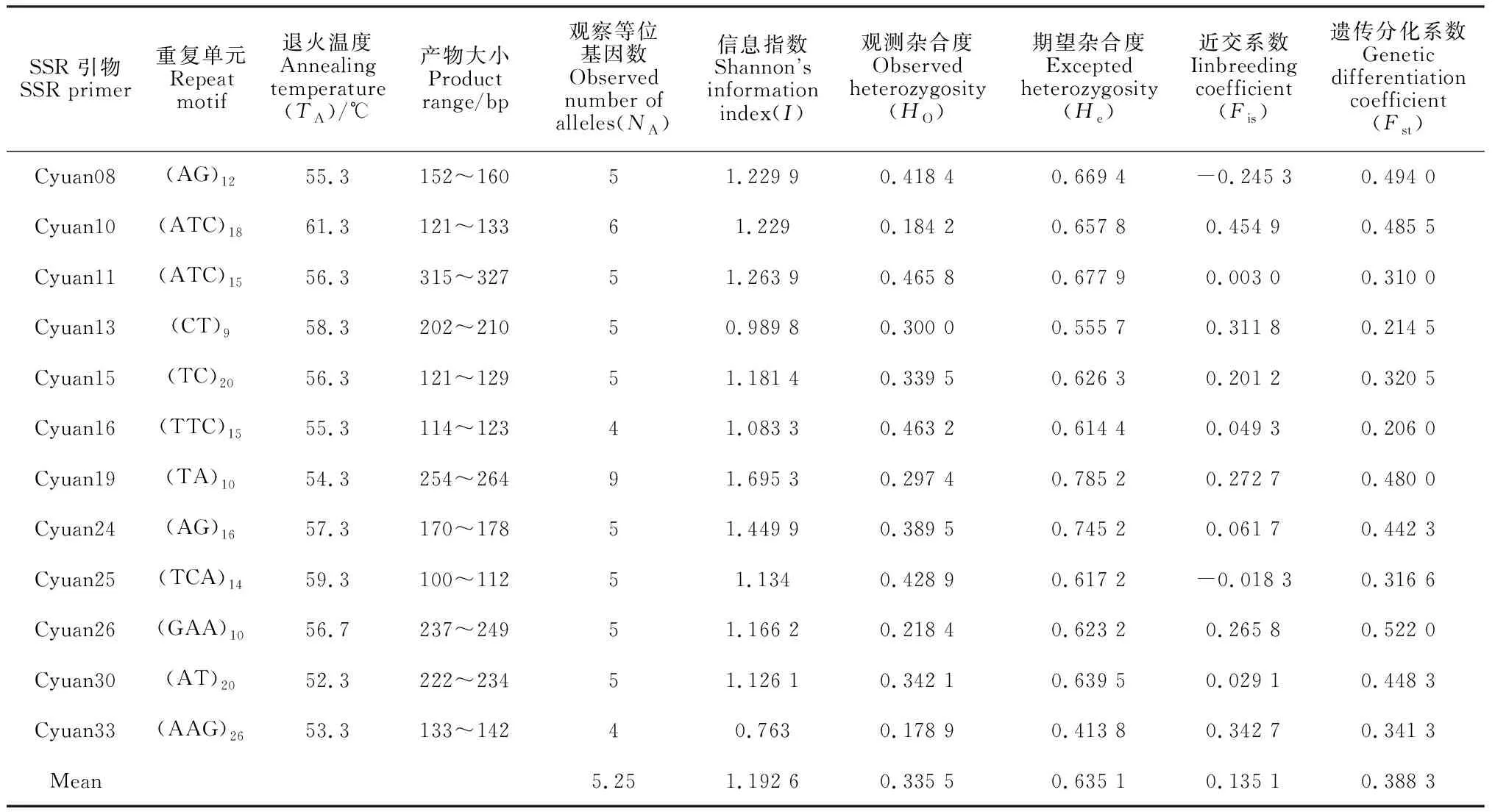
表3 12对延胡索引物的遗传多样性
AMOVA分析表明(表4),47.28%的遗传变异位于居群之间,其中25.94%变异在4个地理单元之间发生,52.72%遗传变异发生在居群内。Nm(基因流)为0.393 8(<1),说明延胡索居群的基因交流较低。Mantel 检测显示,延胡索居群之间的遗传距离与其相应的地理距离呈显著的正相关关系(r= 0.326,P= 0.01)。
3 讨 论
遗传多样性分析是研究物种起源进化、发现新的优异基因、改良现有育种材料的基础工作。因此,延胡索的种质资源及其遗传多样性是延胡索遗传改良和新品种选育的基础。对延胡索遗传多样性的评价有利于了解其遗传背景、保护延胡索种质资源,同时,可以促进有利基因发掘、鉴定延胡索最优亲本组合。通过对19个居群共计360个样本的延胡索研究,12对引物的平均观测杂合度(Ho)为0.335 5,平均期望杂合度(He)为0.635 1,遗传多样性指数为(H)为0.387 9,信息指数(I)为1.192 6,遗传分化指数(Fst)为0.388 3,表现出较高的遗传多样性。
延胡索的遗传结构是该物种自交为主的繁育系统、克隆生长、地理隔离以及居群间有限的基因流共同作用的结果。
繁育系统可以对延胡索居群间遗传分化产生影响。一般来说,自交为主的物种,平均Gst为0.51,即居群间的遗传变异占总变异的一半以上;而异交为主的物种,Gst为0.10,即90%的遗传变异发生在居群内部[7]。本研究中延胡索的Gst为0.388 3,表明延胡索可能是自交为主的繁育系统,这与徐皓[8]等对延胡索传粉生物学的研究结果是一致的。
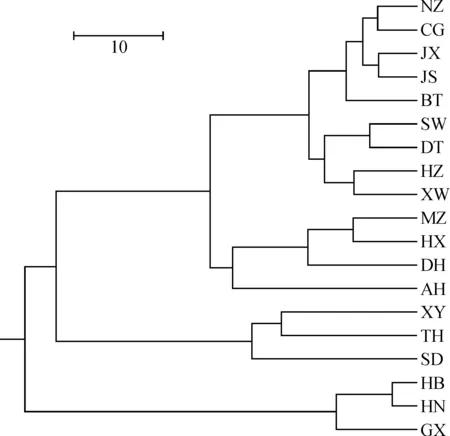
居群编号同表1图2 基于Nei’s无偏差遗传距离的UPGMA 聚类树Population number is the same as Table 1Fig.2 UPGMA dendrogram based on Nei’s unbiased genetic distance
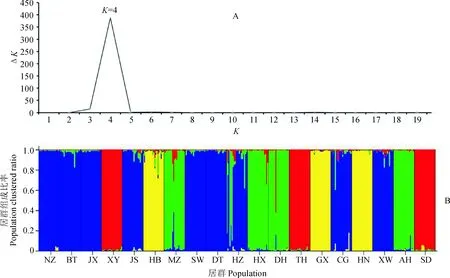
A.基于ΔK值的最适K值计算;B. K=4时Structure 分组结果,图中同一颜色表示属于同一分支图3 19个延胡索种群的贝叶斯聚类图A. Inference of best K using the ΔK method;B. Bar plots showing assignment probabilities from structure analysis when K = 4, the same color in the figure represents the same clusterFig.3 Cluster dendrogram 19 populations of C. yanhusuo based on Bayes analysis
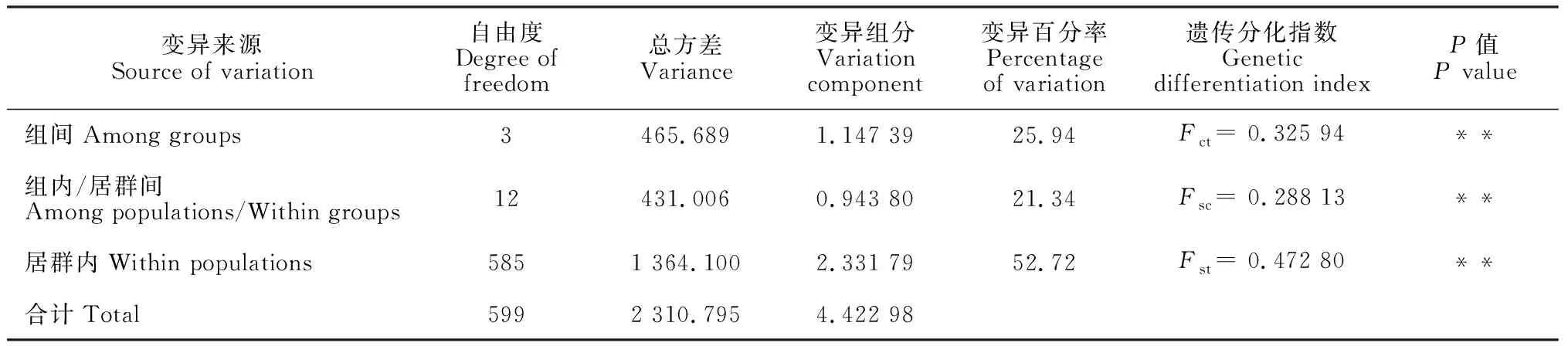
表4 延胡索居群的AMOVA分析
注:Fct.由哈温平衡预期值而来的地区间的遗传差异程度;Fsc.由哈温平衡预期值而来的地区内居群间的遗传变异程度;Fst.由哈温平衡预期值而来的地区间居群间的遗传变异程度Note:Fct. The degree of genetic difference between regions derived from the expected values of Havin equilibrium;Fsc. The degree of genetic difference among populations in the region derived from the expected values of Havin equilibrium;Fst. The degree of genetic difference between populations in the region derived from the expected values of Havin equilibrium
克隆生长也是产生显著遗传分化的一个重要因素。通过野外观察发现,延胡索可以通过其块茎进行无性繁殖。Pluess及 Erickson 等指出[9-10]物种的克隆繁殖会影响有性繁殖的资源供给,最终会导致该物种居群内及其种群间的遗传分化。Cook也指出[11]植物的克隆生长能够增强该物种对资源的利用能力,进而可提高其对环境的适应能力。另外,植物的克隆繁殖还会导致近交衰退和杂合度的降低,进一步影响该植物的适合度,最终导致种群间的遗传分化[12]。因此,延胡索的克隆生长也是其具有显著遗传分化的因素之一。
地理隔离是植物遗传分化显著的重要原因。植物遗传学普遍关注地理环境和遗传变异之间的关系,许多研究表明,地理环境的分布与遗传变异具有一定的关联性。Mantle检测表明,本研究中居群间的遗传距离和地理距离存在着显著相关性(r= 0.326,P= 0.01),说明地理隔离在延胡索目前居群的遗传结构中起着重要作用。本研究中主相关性分析和UPGMA聚类保持一致,将19个居群的延胡索样本分为4个组,其中华北聚为一支,华东聚为一支,华中、华南地区聚为一支,浙江的及引种自浙江的几个居群聚为了一支,这也验证了Mantle检测的分析结果,表明地理隔离在遗传分化中起着作用。
群体的遗传变异是基因流和选择作用的综合结果。同种植物各个群体空间上的隔离、突变、环境因子造成的选择差、随机遗传漂变、基因流的隔离都能导致群体基因结构的空间异质性,从而促进群体分化[13]。Slatkin[14]认为当基因流大于 1 时,能发挥其均质化作用,维持遗传变异的多样性;基因流小于 1 时,遗传分化可以由遗传漂变引起,进而对居群遗传结构造成影响。通常情况下,基因流受限的植物比基因流广泛的植物遗传分化大。本研究延胡索居群间基因流Nm为0.393 8,小于1,表明受限的基因流也是其具有显著遗传分化的因素之一。
野生延胡索的繁殖能力强,但适应能力弱,加之人为过度采挖造成野生资源的急剧减少。近几年在陕西、四川和云南等地陆续开始延胡索的大面积种植,在一定程度上缓解了需求增长和资源危机的矛盾。本研究中延胡索的遗传特征,不仅提供了有关延胡索遗传结构的重要见解,而且对延胡索的养护管理也至关重要。实地考察表明,人类活动造成了延胡索居群的急剧缩小和居群的碎片化。这种条件可能使延胡索对遗传漂变的耐受性很低,可能导致其资源灭绝。因此,维持有效的居群规模和减少人为干扰,是延胡索保育工作的先决条件。基于延胡索在居群水平上遗传多样性低,物种水平上的遗传多样性丰富,居群间遗传分化显著,我们提出以就地保护为主的保护策略,并建立多个保护点,从而保护自然居群及其周围的生境,扩大延胡索居群的规模,让其进行自然更新,从而对延胡索的遗传多样性进行保护。
