施氮及不同根系分隔模式对尾叶桉和降香黄檀幼苗生长及叶片生理特性的影响
2018-07-19许峻模龙佳峰汤文艳田诗韵叶绍明
许峻模,潘 婷,龙佳峰,汤文艳,田诗韵,叶绍明
(广西大学 林学院,南宁530004)
氮素作为植物需求量最大的矿质元素,参与植物体内蛋白质和核酸的合成代谢,是植物正常生长必需的基本元素之一[1-2],影响着植物的光合作用[3-4]、水分吸收利用[5]、抗氧化系统[6]、内源激素[7]以及土壤微生物代谢[8]等。在生产实践中,为了提高人工林产量,施用氮肥是常采用的营林措施之一。但随着氮肥施用量的增加,经常出现氮肥使用过量的现象,造成地下水污染及江河湖泊的富营养化等一系列问题[9]。固氮微生物能将大气中游离的氮素转变成含氮化合物,为植物提供氮素来源,从而可减少氮肥的使用[10]。因此,如何通过有效的方式将生物固氮机制引入非豆科植物尤其是速生树种中,进而建立起非豆科植物的固氮新体系,成为现代林业科学研究中迫切需要又富有挑战性的研究课题。
桉树因其速生丰产、抗逆性强、经济效益高等优点,已成为华南地区的主要商品林之一,为国民经济的发展做出了突出贡献[11-12]。但由于长期以来过分追求短期生产力与经济利益,大面积桉树纯林多代连栽造成的土壤质量退化、生物多样性下降及林分稳定性差等一系列生态环境问题,成为了实现桉树可持续发展的最大障碍[13-14]。为解决人工纯林带来的生态问题,南方地区逐步营造了人工混交林,其构成的混交生态系统优势突显,有效提高了林地土壤肥力和林地生产力。而在优良的混交林生态系统中,具有固氮能力的豆科植物起到了关键性的作用。一些试验指出将桉树与部分固氮树种混交可提高桉树人工林稳定性和可持续性,种间关系协调的混交林不仅可以改善林地生产力,减少水土流失,还可以防止地力衰退,增加单位面积林产品和副产品的产量[15]。目前,树种之间的生物固氮效应已广泛应用于桉树混交林的研究。
降香黄檀为国家二级重点保护濒危树种,因其根部有固氮根瘤菌,常作为改良桉树人工林土壤理化性质的主要混交树种[16],但目前国内外对桉树与降香黄檀混交的研究仅限于桉树与降香黄檀混交的生长特性[17]、土壤呼吸[18]、土壤微生物[19]等方面。且桉树/降香黄檀混交林生态系统只是最近几年时间才引起广泛关注,针对其根系互作对养分吸收利用规律的认识还十分有限,尤其是混交过程中根系互作对氮素吸收的影响研究鲜见报道,而混交系统优势往往又体现在其地下部的根系作用。为此,本文通过盆栽可控试验分析根系分隔和不同施氮水平对尾叶桉和降香黄檀幼苗生长及叶片生理特性的影响,为探讨尾叶桉/降香黄檀混交群体氮素竞争互补关系、利用效率及根系互作机制提供科学依据。
1 材料和方法
1.1 试验材料
实验于2016年2月到8月在广西大学试验大棚(22°13′~23°32′ N,107°45′~108°51′ E)中进行。该区域属于湿润的亚热带季风气候,年平均气温在21.6 ℃左右,年均降雨量达1 304.2 mm,平均相对湿度为79%。供试苗木为3月生的广林9号尾叶桉与1年生的降香黄檀,育苗土壤为南宁市郊区马尾松成熟林下0~40 cm土层土壤,其质地为粘壤土,典型的偏酸性赤红壤。将实验用土经风干、敲碎、过筛后与珍珠岩按8∶2的比例混匀待用。本次试验所施氮肥为尿素,N15的丰度为10.3%。
1.2 试验设计
本试验采用双因素完全随机设计,因素A为不同施氮水平:不施肥(对照)、3 g N /株、6 g N /株;因素B为不同根系间隔模式:不隔(盆钵中间的根系不分隔,种间根系竞争作用和促进作用同时存在)、网隔(盆钵中间采用30 μm孔径的尼龙网分隔,消除种间根系竞争作用)、膜隔(盆钵中间采用塑料膜隔开并保证密封性,消除种间根系相互作用)。试验共组成9个处理组合(表1),每个处理组合4次重复。
采用盆栽试验方式,采用高60 cm 直径40 cm的无纺布育苗袋为种植容器,装入适量土壤,使得土层表面离容器上缘5 cm左右。选取生长良好、长势均匀的尾叶桉与降香黄檀幼苗各1株种植于每个育苗袋中。分别于苗木种植后1个月、3个月各施肥1次,即在2棵植株根系周围挖穴,将肥料溶于纯水均匀施入后用表土覆盖。并人工定期定量对苗木进行浇灌,每盆定量浇灌8 000 mL,保证苗木充足水分供应。
1.3 指标测定与方法
1.3.1形态指标2016年8月对所有幼苗分地上部和根系两部分进行收获。苗高和地径测定分别用直尺和电子游标卡尺测量;生物量测定采用全株收获法,分别测定根、茎以及叶的鲜重后,将各器官带回实验室内用烘箱于80 ℃中烘干至恒重,称量并计算各组分的生物量;将盆栽盆体用水浸泡取出完整根系后采用Epson根系扫描仪进行根系扫描,获取形态结构图像后,根系各形态指标(根系总长度、表面积、体积、平均直径和根尖数)采用根系图像分析软件WinRHIZON Pro 进行测定。
1.3.2生理指标将所有试验植株洗净晾干后带回实验室进行相关生理指标测定。叶片超氧化物歧化酶(SOD)、过氧化物酶(POD)活性分别采用氮蓝四唑法和氧化愈创木酚法测定[20],可溶性蛋白含量测定采用考马斯亮蓝G-250染色法[21]测定,丙二醛(MDA)含量采用硫代巴比妥酸法[22]测定。
1.4 数据处理与分析
所有试验数据用Excel 2010进行整理后,运用SPSS 19.0分别对施氮水平、分隔模式处理进行方差分析(One-way ANOVA),并对差异显著性指标进行多重比较(LSD法);采用Sigmaplot 10.0 软件进行作图。
2 结果与分析
2.1 施氮和根系分隔对尾叶桉和降香黄檀幼苗生长的影响
表1显示,同一分隔模式下,尾叶桉的地径、苗高均随氮素水平的增加而增加,且不同氮素水平间地径的差异均达显著水平(P<0.05)。而同一氮素水平中,分隔模式对尾叶桉地径的影响表现为不隔>网隔>膜隔,除施氮0 g·株-1水平的网隔与膜隔(T4与T7)、3 g·株-1水平的不隔与网隔(T2与T5)差异不显著外,其他氮素水平不同分隔模式间均有显著差异(P<0.05);尾叶桉苗高则在施氮0 g·株-1、6 g·株-1水平下,分隔模式对苗高生长均表现为不隔>网隔>膜隔,而3 g·株-1水平下的苗高生长规律与之相反,除施氮6 g·株-1水平的3种分隔模式以及3 g·株-1水平的不隔与膜隔(T2与T8)苗高存在显著差异外(P<0.05),其他同一氮素水平下不同分隔模式对苗高生长的影响差异不显著。9个对比处理中,尾叶桉地径在T3处理下生长量达到最大值,比最小值T4处理增6.62 mm。苗高则同在T3处理达到最大值,值为119.21 cm,比最小值T7处理增38.83 cm。
氮肥的施入对降香黄檀的地径、苗高生长均有明显的促进作用,并随着氮素水平的增加,降香黄檀地径、苗高均呈递增趋势(表1)。同一分隔模式下,不同氮素水平间降香黄檀的地径存在显著差异(P<0.05),而同一氮素水平中,不同分隔模式下降香黄檀的地径生长规律为膜隔>不隔>网隔,不同分隔模式地径的差异均达显著水平(P<0.05);同一氮素水平下,分隔模式对降香黄檀苗高影响与地径表现规律一致,均为膜隔>不隔>网隔,除施氮6 g·株-1水平的网隔和膜隔(T6与T9)存在显著差异外(P<0.05),其他施氮水平下不同分隔模式间对苗高生长差异不显著。9个对比处理中,降香黄檀地径最大为T9处理,其比最小值T4处理高74.1%,与地径出现最大值的规律一致,降香黄檀苗高在T9处理达到最大值,值为27.88 cm,其比最小值T4处理高54.4%。
2.2 施氮和根系分隔对幼苗叶片生理指标的影响
2.2.1施氮和根系分隔对尾叶桉叶片生理指标的影响由表2可知,施氮能够使尾叶桉叶片SOD、POD活性、可溶性蛋白含量均显著升高。其中,施氮3、6 g·株-1状态下,不隔模式尾叶桉叶片SOD活性分别为不施氮的1.12和1.43倍,网隔模式分别为1.15和1.25倍,而膜隔模式分别为1.21和1.26倍;不隔模式叶片POD活性分别为不施氮的1.19和1.46倍,网隔模式分别为1.22和1.34倍,而膜隔模式分别为1.25和1.43倍;不隔模式可溶性蛋白含量则分别为不施氮的1.04和1.17倍,网隔模式分别为1.02和1.09倍,而膜隔模式分别为1.02和1.21倍。这说明施氮能够使尾叶桉维持较高的SOD、POD活性,以减轻脂膜过氧化作用对细胞膜的伤害。同时,施氮促使尾叶桉叶片MDA含量显著降低(表2)。其中不隔模式MDA含量降幅分别为不施氮的35.29%和52.10%,网隔模式分别为11.90%和50.00%,而膜隔模式分别为30.88%和52.21%。
2.2.2施氮和根系分隔对降香黄檀叶片生理指标的影响表2表明,施氮对降香黄檀叶片SOD、POD活性、可溶性蛋白含量均有显著影响(P<0.05),

表1 不同处理下尾叶桉和降香黄檀幼苗地上部分生长情况
注:表内数据为平均值±标准差;同列不同小写字母表示处理间差异显著(P< 0.05);下同Note: Values in the table are given as mean±SD; Different letters in the same column show significant difference among treatments at 0.05 level(P< 0.05); The same as below

表2 不同处理对尾叶桉和降香黄檀叶片生理指标的影响
且均随施氮量的增加而升高。其中,在施氮3、6 g·株-1状态下,降香黄檀叶片SOD活性增幅最大的是膜隔模式,增幅分别达25.33%和42.89%,随后依次是不隔模式和网隔模式;不隔模式叶片POD活性分别较不施氮处理提高了8.38%和21.24%,网隔模式分别为4.58%和15.52%,而膜隔模式分别为8.79%和25.49%;同时,施氮也能使降香黄檀叶片可溶性蛋白含量显著升高,且施氮6 g·株-1增幅均大于3 g·株-1,其中在6 g·株-1水平下,可溶性蛋白含量增幅最大的是膜隔模式(63.24%),最小的是网隔模式(61.55%),而在3 g·株-1水平下,可溶性蛋白含量增幅最大的则为网隔模式(32.33%),最小的为膜隔模式(10.88%);与不施氮处理相比,2个施氮环境下(3、6 g·株-1)降香黄檀叶片MDA含量显著降低,其中不隔模式叶片MDA含量分别降低34.46%和58.11%,网隔模式分别为35.48%和57.26%,而膜隔模式分别为34.23%和58.56%。表明降香黄檀适量施氮能够降低其体内丙二醛的积累量,从而减少对其的伤害,提高自身耐受性。
2.3 施氮和根系分隔对幼苗根系生长的影响
2.3.1施氮和根系分隔对尾叶桉根系生长的影响从表3可以看出,同一分隔模式下,施氮量的增加显著增加了尾叶桉根系的总长度、总表面积、总体积、平均直径、根尖数和比根长,且随氮素水平的增加呈增加趋势,各氮素水平间的差异均达显著水平(P<0.05);同一氮素水平下,不同分隔模式下尾叶桉的各项根系指标总体表现为不隔>网隔>膜隔(除施氮0 g·株-1、3 g·株-1水平下根总长度及0 g·株-1水平下根总表面积外),且不同分隔模式间根系生长基本存在显著差异(P<0.05)。T3的根系生长在9个处理中达最大值,而除T4的根尖数、比根长为最小值外,其他根系形态生长则均是T7为最小,最大值分别是最小值的1.91、2.02、2.79、3.13、2.07、1.85倍。
2.3.2施氮和根系分隔对降香黄檀根系生长的影响表4结果显示了不同处理对降香黄檀根系生长的影响,同一分隔模式下,降香黄檀的根系生长量在氮素水平上均表现为6 g·株-1>3 g·株-1>0 g·株-1,表明氮肥的施入显著促进了降香黄檀根系的生长。同一分隔模式下,不同氮素水平的降香黄檀根总长度、根总表面积、根总体积、平均直径以及根尖数存在显著差异(P<0.05),比根长则T1处理与T2处理及网隔模式下3种氮素水平间差异不显著。降香黄檀的根系生长规律与尾叶桉有所不同,同一氮素水平下,其根系生长总体表现为膜隔>不隔>网隔,各分隔模式间差异达显著水平。同时发现,T9根系的各生长指标在9个处理中均达到最大值,而T4处理下的根系生长指标则相对较小(除根尖数外)。根总长度、根总表面积、根总体积、平均直径、根尖数、比根长最大值比最小值分别高54.75%、50.24%、64.47%、58.25%、45.52%、68.98%。
表3 不同处理对尾叶桉根系生长的影响

表4 不同处理对降香黄檀根系生长的影响
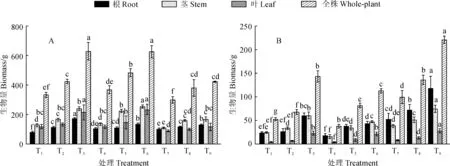
图1 不同处理对尾叶桉(A)和降香黄檀(B)各器官以及总生物量的影响Fig.1 Effect of different treatments on the organs and total biomass of E. urophylla and D. odorifera
2.4 施氮和根系分隔对幼苗生物量积累及其分配的影响
2.4.1施氮和根系分隔对尾叶桉幼苗生物量的影响如图1所示,同一分隔模式下,不同氮素水平间尾叶桉各器官生物量及总生物量的表现均为6 g·株-1>3 g·株-1>0 g·株-1。其中,对根系生物量而言,生物量最大的处理T3(173.60 g)高出最小处理T1(80.73 g)达92.87 g,不同氮素水平下根生物量均存在显著差异(P<0.05)。茎的生物量同样随着氮素水平的增加而增加,施氮6 g·株-1比不施氮在3种分隔模式下分别增加了44.8%、44.1%、35.8%,且除T8与T9无显著差异外,其他氮素水平间茎生物量差异均达显著水平(P<0.05)。而对叶生物量而言,其总体大小顺序为T6>T3>T5>T2>T4>T1>T9>T8>T7,施氮0 g·株-1水平和膜隔模式下生物量差异不明显。不同分隔模式下,尾叶桉膜隔模式下的总生物量最小,除T1与T4、T2与T8及T3与T6及间差异不显著外,其他同一氮素水平下不同分隔模式间均存在显著差异(P<0.05)。其中T3的总生物量达到最大值628.57 g,是最小值T7(299.51 g)的2.1倍。
2.4.2施氮和根系分隔对降香黄檀幼苗生物量的影响施氮能够有效促进降香黄檀苗高和地径的生长,显然其生物量也随着氮素施入而增加(图1)。同一分隔模式下,降香黄檀幼苗的根系生物量、茎生物量、叶生物量以及全株生物量均表现为6 g·株-1>3 g·株-1>0 g·株-1,除T1和T2的根、叶生物量以及T5与T6根生物量不存在显著差异外,其他器官以及总生物量各水平间均存在显著差异。同一氮素水平下,各器官以及总生物量均为在膜隔模式下达到最大,而分别在施氮3 g·株-1状态下不隔模式及施氮0 g·株-1、6 g·株-1水平网隔模式下达到最小,不同分隔模式间的全株生物量存在显著差异(P<0.05)。无论是根、茎、叶生物量还是全株总生物量,施氮6 g·株-1水平膜隔模式(T9)显著大于其他处理,且比最小处理的生物量(T4)分别多84.89%、78.24%、87.38%和82.97%。
3 讨 论
3.1 施氮及根系互作对尾叶桉/降香黄檀幼苗地上部生长及叶片生理特性的影响
植物生长除了受自身遗传物质的影响外,还会受外界环境因子如水分、光照、土壤等的作用[23]。氮素营养是影响植物生长发育的重要因子,其在一定供应范围内,对植物苗高、地径的生长均有明显的促进作用[24-25]。本研究中,施氮对尾叶桉和降香黄檀的苗高、地径生长具有明显的促进作用,且随着施氮水平的增加,尾叶桉和降香黄檀叶片超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性及可溶性蛋白含量显著升高,而丙二醛(MDA)含量则呈降低趋势。由此可知,与不施氮相比,施氮3、6 g·株-1能够提高两植株幼苗叶片SOD、POD的活性,较大程度上减缓了膜脂过氧化反应对植株造成的伤害;同时,叶片可溶性蛋白的含量显著提高,有效增加其细胞的渗透调节能力,为其生长提供更多的能量来源,而MDA含量降低则提高了植株细胞的生理功能,进一步使其维持较高的生命活力[26]。同时发现,不隔模式显著提高了尾叶桉的苗高、地径,而降香黄檀苗高、地径增长却在根系膜隔下表现最好,这可能是由于植物的生长除了受N元素供应影响外,还会受到生长空间和种间根系的相互作用,从而影响养分的吸收利用。根系不分隔处理下,尾叶桉因其速生特性,根系相互作用优势明显,其可以占据降香黄檀地下部空间,但降香黄檀的根却较少到混交尾叶桉的地下部空间。本研究发现,尾叶桉根系总长度及根总表面积均高于降香黄檀,侧面反映了混交扩展了尾叶桉根系水平尺度的养分生态位,使得其通过根系吸收更多土壤养分,从而在3种分隔模式中生长表现最优。而非固氮树种与固氮树种混交后,能够提高非固氮树种营养水平改善土壤养分[27]。虽不隔模式下尾叶桉或降香黄檀地下部养分和水分的竞争作用和促进作用并存且相互作用强烈,但树种间的相互作用改变其植株体内的养分释放过程[28],提高了降香黄檀的固氮能力,从而弥补了养分部分流失的不足,使得降香黄檀在不隔模式生长较网隔模式好。这与苗锐等[29]研究发现3种间作体系中不隔处理的蚕豆结瘤数目均多于膜隔处理的结果相似;根系用尼龙网分隔,则使尾叶桉或降香黄檀其中一种植物根系不能进入另一植物根区竞争养分,地下部根系间没有直接接触,尾叶桉根系生长虽受到一定程度限制,但由于可移动的养分及水分仍可以通过,尾叶桉从中吸收部分养分,从而使其生长略低于不隔模式而优于膜隔模式。而此模式下的降香黄檀生物固氮量主要由其地上部库强决定,缺少尾叶桉地下部的互作作用致其地下部固氮能力有限,且土壤中的养分交换过程存在部分流失,养分不足是造成网隔模式下降香黄檀生长相对较差的重要原因;根系用塑料膜分隔,该处理下植物根系、养分和水分不存在物质交换,地下部的种间相互作用减至最小,尾叶桉无法获取更多养分且根系生长受到一定程度限制,因而其在膜隔模式下生长最差。降香黄檀则正好相反,其通过共生固氮将大气中的氮素转化为有机氮素固定于植物体内,且缺少了尾叶桉对其地下部水分与营养元素的竞争后,充足的养分及自身固氮能力令其表现出膜隔模式下生长最好。
3.2 施氮及根系互作对尾叶桉/降香黄檀根系形态的影响
植物所需的氮素绝大部分来自于土壤,而植物对土壤氮素的竞争互补利用、氮阻遏消减和氮转移等调控氮素利用的生理、生态学过程,很大程度上是依赖于根间互作产生的[30-31]。随着土壤环境的改变,根系往往会通过改变自身的生长以及形态等来适应外界环境的变化[32]。当土壤氮供应充足时,植物的根总长度、表面积和平均直径均会增加,同时促进植物根系根尖数和侧根的形成,但均在一定氮素范围内表现出此现象[33-34]。本研究结果与前人一致,随着氮素施入浓度的增加,无论是尾叶桉还是降香黄檀其根系总长度、总表面积、总体积、平均直径以及根尖数和比根长均显著的提高。根系总长度、根系表面积、体积等根系参数是决定根系养分吸收范围、吸收强度的重要指标,通过延伸根长、扩大根系表面积、增加根系总表面积有助于水肥的获取,促进整株植物的生长[35-36]。植物根系的比根长增大有效加快对养分和水分的利用效率,从而提高自身竞争力[37],氮素水平为6 g时,不论是尾叶桉还是降香黄檀的比根长都最大,更有利于对水肥的吸收。尾叶桉在混交中具有较强的水肥竞争力,竞争优势明显高于降香黄檀,在根系不分隔时,尾叶桉占据更多的养分资源,根系生长得到显著提高。而在根系膜隔条件下,降香黄檀根系生长不受尾叶桉根系以及养分竞争的影响,表现出生长最好,这与前人研究大豆与小麦的间作体系得出的结果一致[38]。通常比根长较大的植物有较多细根、较大根表面积[39],其在相同养分及水分条件下对地下资源获取能力更强,有助于地上部分的生长,提高植物光合效率,进一步向根系分配更多的物质促进根系发育[40],从而解释了降香黄檀苗高、地径和生物量均在膜隔条件下生长相对其他2种模式更好,而尾叶桉的苗高、地径、生物量在不隔模式下达到最好。
3.3 施氮及根系互作对尾叶桉/降香黄檀生物量的影响
生物量可作为植物对环境因子响应最直观的指标,研究其变化对了解植物对氮素的响应具有重要的意义[41]。本研究中,无论是何种分隔模式,各器官以及全株生物量积累均随着氮素水平的增加而增加。这可能是由于盆栽试验土壤营养元素有限,通常在这种情况下,随着供氮的增加,植物地上部分的生物量以及全株生物量也会随之增加。而在所有处理中,尾叶桉茎所占的生物量比例最大,其次是N0和N3水平下的叶生物量,这主要是因为当土壤氮素营养较低时,尾叶桉需要将生物量更多地分配到地上以占据高度优势和生产更多光合产物以供生长[40]。而在N6水平下氮素营养充足,不但显著增加地上部分的生长,且将更多的光合产物分配至地下,促进根生物量的增长。这可能与尾叶桉的速生特性有关,在氮素营养充足情况下尾叶桉会向根系分配更多的生物量,促进其根系生长,从而扩大其获取水分和养分的空间。这与部分学者的得出的在氮不足的情况下,根系生物量累积较多的结果相反[42-44]。安慧等[45]在分析根域限制对连翘幼苗生长的影响发现,根域体积减小后,根系的伸长会受阻,根系活力下降,从而导致根的重量减小。也有研究表明,根域限制减少了地上部分细胞分裂素以及叶面积和抑制侧枝生长而阻碍地上部生长[46],以致同时影响了尾叶桉的地上部分与地下部分的生长,造成膜隔模式下尾叶桉的总生物量最小。就降香黄檀而言,9个处理中根系都占有较大比例,根系在分隔模式下的总生物量显著高于网隔和不隔模式,可能是由于无论是氮素较少还是充足条件下,降香黄檀首先促进根的生长以获取更多的水分与营养,再进一步调节地上与地下部分生物量的分配,而没有根系的相互作用,使得降香黄檀的生物量在膜隔下表现累积最多。
