右侧背外侧前额叶在视觉工作记忆中的因果性作用*
2018-07-07王思思库逸轩
王思思 库逸轩
(华东师范大学心理与认知科学学院, 上海 200062)
1 引言
视觉工作记忆(visual working memory)是一个能够对环境中的视觉信息进行暂时性地储存和加工的在线工作空间(Baddeley & Hitch, 1974; Cowan,2001), 它在我们脑海中形成暂时性的视觉表征以应对接下来环境中不断出现的刺激。作为连接大脑和外部世界的中继站, 视觉工作记忆对许多高级认知过程如推理、问题解决等具有重要作用(Fukuda,Vogel, Mayr, & Awh, 2010; Alloway & Alloway,2010)。虽然人脑每时每刻都要处理纷繁世界中的海量信息, 大量的行为、电生理和脑成像研究证据表明视觉工作记忆只能表征数量非常有限的物体(3~4个) (Luck & Vogel, 1997; Vogel & Machizawa,2004; Todd & Marois, 2004), 并且不同个体之间的视觉工作记忆容量存在很大差异(Vogel & Machizawa,2004; Vogel, McCollough, & Machizawa, 2005)。
背外侧前额叶皮层(dorsolateral prefrontal cortex,DLPFC)在视觉工作记忆的存储和加工中发挥重要的作用。神经元电生理记录研究表明, DLPFC在视觉工作记忆的延迟阶段出现持续性的、增强的激活(Funahashi, Bruce, & Goldman-Rakic, 1989; Fuster& Alexander, 1971; Kubota & Niki, 1971)。DLPFC主要负责工作记忆空间信息的加工(Courtney, Petit,Maisog, Ungerleider, & Haxby, 1998), 视觉空间工作记忆的维持和操作(Curtis, 2006; D'Esposito,Postle, Ballard, & Lease, 1999), 注意控制和转换(Vanderhasselt, De Raedt, & Baeken, 2009), 以及干扰抑制和信息刷新(Anderson et al., 2004;Toepper et al., 2010)等功能。
此外, 根据Baddeley和Hitch的经典工作记忆模型, 工作记忆的存储和加工器中包含语音缓冲器和视空模板两个子系统, 前者负责对言语信息的加工, 后者负责对视觉和空间刺激的处理(Baddeley& Hitch, 1974)。脑成像研究表明:大脑左侧DLPFC和右侧 DLPFC对加工材料存在偏侧化差异, 二者分别负责言语和视觉空间工作记忆信息的加工(Smith,Jonides, & Koeppe, 1996; Walter et al., 2003)。
然而这些神经电生理和神经影像学的研究都是相关性的研究, 并不能说明 DLPFC在工作记忆加工中的因果性作用。经颅直流电刺激(transcranial direct current stimulation, tDCS)是一种无创性的微弱电流刺激, 通过影响神经元膜电位的变化对大脑皮层的兴奋性产生影响(Priori, 2003)。在调控运动皮层功能的研究中发现, 正性电刺激能够通过增强神经元同步活动使得刺激皮层去极化而兴奋性提升, 而负性电刺激则会通过令神经元超极化而使刺激皮层活动受到抑制。tDCS的效果在电刺激结束后可以持续较长时间, 13 min强度1 mA的电刺激效果可以在运动皮层持续约90 min (Nitsche & Paulus,2001)。这种“后效”很有可能和受NMDA受体调节的长时程增强(LTP)作用相似的神经可塑性变化有关(Nitsche, Fricke, et al., 2003)。作为一种研究大脑皮层和其对应认知功能因果联系的有效手段, 经颅直流电刺激技术已经应用到注意、情绪、决策等大脑功能的各个方面(Coffman, Clark, & Parasuraman,2014; Dedoncker, Brunoni, Baeken, & Vanderhasselt,2016)。
前人通过经颅直流电刺激探究前额叶在工作记忆中的作用的研究中, 比较一致地发现正性电刺激左侧 DLPFC可以显著提升言语工作记忆表现(Fregni et al., 2005; Andrews, Hoy, Enticott, Daskalakis,& Fitzgerald, 2011; Ohn et al., 2008 ), 证明了左侧DLPFC在言语工作记忆中存在加工优势。对于右侧DLPFC, Wu等人发现正性电刺激右侧DLPFC能够显著提升有干扰情况下的空间工作记忆顺序回忆任务的反应速度以及逆序回忆任务的记忆广度,而对无干扰情况下的顺序和逆序回忆任务的表现没有影响(Wu et al., 2014)。Li等人采用颜色的变化检测任务发现, 正性电刺激右侧 DLPFC能够显著提升被试在有干扰情况下的视觉工作记忆表现(Li et al., 2017)。另一项研究使用正性tDCS刺激右侧DLPFC显著提升了视觉空间延迟再认任务的正确率(Giglia et al., 2014)。这些研究结果为右侧DLPFC在视觉空间工作记忆的干扰抑制及信息维持过程中的重要作用提供了有力的证据。但是目前仍缺乏研究结果证明右侧DLPFC在视觉工作记忆中的作用是否受到工作记忆负荷的调节。神经影像学证据表明, DLPFC血氧信号水平随着记忆负荷的增加而增强(Druzgal & D'Esposito, 2003; Rypma, Berger, &D'Esposito, 2002), 因此在不同记忆负荷下, 电刺激DLPFC的效果也可能不同。为了探究这一问题, 本研究设置了不同的工作记忆负荷, 并通过经颅直流电刺激探究右侧DLPFC在不同记忆负荷中的作用。
除此之外, 电刺激右侧 DLPFC影响视觉工作记忆表现的神经机制的研究仍较少, 电刺激右侧DLPFC影响的是视觉工作记忆的编码、保持还是提取过程仍不清楚, 本研究采用经典的视觉工作记忆变化检测范式(Vogel & Machizawa, 2004; Tseng et al., 2012), 这一范式的优点是能够通过示例序列呈现, 消失和探测序列呈现将工作记忆的编码、保持和提取过程分离开来。通过这种能够明确区分工作记忆不同加工阶段的范式结合脑电信号记录的方式, 探究右侧 DLPFC在视觉工作记忆不同负荷任务中发挥作用的阶段。
N2pc (Negativity posterior contralateral, Luck &Hillyard, 1994b)是视觉目标对侧视野的电极波幅与目标同侧视野电极波幅的差值波, 它是主要出现在大脑后部电极的负波, 一般出现在刺激呈现之后约180~280 ms, 并且在PO7/PO8电极周围波幅最负。N2pc主要反映了对目标的注意选择(Mazza & Caramazza,2012)。SPCN (Sustained posterior contralateral negativity,Jolicœur, Sessa, Dell’Acqua, & Robitaille, 2006) 同N2pc一样, 也是一个在目标呈现对侧更负的偏侧化事件相关电位成分。也称作CNSW (Contralateral negative slow wave, Klaver, Talsma, Wijers, Heinze,& Mulder, 1999)或CDA (Contralateral delay activity,Vogel & Machizawa, 2004)。它主要出现在刺激呈现之后 300 ms以后, 并随着视觉工作记忆中信息的保持持续。SPCN的波幅也是在PO7/PO8电极周围最强。SPCN反映了视觉工作记忆中信息的维持,它的波幅会随着信息存储数量的增加而增加, 直到到达被试的最大记忆容量后不再变化(Robitaille et al., 2010; Vogel & Machizawa, 2004)。本研究旨在通过这两个电生理指标, 探究右侧 DLPFC在视觉工作记忆提取阶段中发挥作用的机制, 究竟是通过影响注意过程还是信息维持过程来影响工作记忆表现的。
2 方法
2.1 被试
40名华东师范大学本科生或研究生参加了实验, 年龄在19~24岁之间, 平均年龄21.9岁, 标准差1.8, 其中男生19名。所有被试的视力或矫正视力正常, 无色觉障碍, 均为右利手, 经被试口头报告, 身体健康无神经系统疾病, 没有脑部损伤史,体内无金属植入。所有被试均为自愿参加实验, 在参加实验之前都签署被试知情同意书, 实验完成后获得少量报酬。
2.2 实验范式
本研究采用了改进的视觉工作记忆变化检测任务(Vogel & Machizawa, 2004) (图1), 首先会在灰色的屏幕上呈现一个向左或向右的箭头加注视点, 被试的眼睛需要始终盯着注视点, 同时用余光注意箭头的指向。接下来示例序列呈现100 ms, 在注视点的左右两侧出现相同数量的不同颜色的小方块, 被试需要眼球始终盯住中间的注视点, 同时用余光记忆箭头指向一侧屏幕上小方块的颜色(4个或者 6个)。随后示例序列消失, 在注视点呈现900 ms后出现探测序列, 在注视点两侧出现与示例序列位置和数量相同的色块, 要求被试判断箭头指向一侧屏幕上小方块的颜色相对于示例序列有没有发生变化, 如发生变化, 只会有一个小方块的颜色发生变化。变化按F键, 不变按J键, 变化与不变化的概率各为50%, 按键在被试间平衡。
所有的小方块都呈现在两个4.3° × 8°的长方形区域内, 两个长方形区域的边缘间距 0.7°。每个区域内每次呈现4或者6个0.9° × 1.1°大小的色块, 色块的颜色从蓝、绿、红、紫、黄、白、黑七种颜色中选取, 所有颜色在示例序列里出现不超过两次。两两小方块中心之间的水平间距为 1.6°, 垂直间距为 1.4°, 位置不会发生重叠。被试眼睛和屏幕的距离是100 cm。
被试需要分两次(电刺激和虚假刺激, 分在两天)完成两个系列的任务, 每个系列包括 4小节任务, 每小节任务需要完成72个试次, 每个系列任务中被试需要完成负荷 4(每侧 4个色块)和负荷 6任务(每侧6个色块)各144个试次。每个被试共需要完成576个试次。
2.3 经颅直流电刺激
本实验使用德国 NeuroConn GmbH公司生产的 DC-STIMULATOR PLUS经颅直流电刺激仪施加电刺激。将两个塑胶电极片(5 cm × 7cm)用浸过盐水的海绵片包裹并贴于刺激的大脑位置。在正性电刺激右侧DLPFC情况下, 正极电极放置于10-20脑电系统的 F4电极位置, 参考电极放置在被试的左侧脸颊以避免电刺激对其他脑区的影响(Jones &Berryhill, 2012)。真刺激情形下, 采用强度为1.5 mA的直流电刺激持续刺激 15 min; 虚假刺激情形下,电极的正负极放置位置与真刺激相同, 但是电流刺激仅在开始时持续30 s, 被试佩戴电极片共15 min后取下, 以使被试感受到和持续电刺激相同的轻微刺痛感, 并且难以区分接受的是哪种电刺激。由于虚假刺激情况下电流时间非常短, 因此将虚假刺激作为电刺激右侧DLPFC的控制情况。为了消除电刺激的后效, 每名被试需要分两次(分别在两天,间隔 24 h以上)参加实验。被试接受电刺激右侧DLPFC和虚假刺激的顺序也在被试间进行平衡。被试在接受 15 min电刺激之后, 立刻开始视觉工作记忆任务, 并同时记录脑电信号。
2.4 脑电记录参数
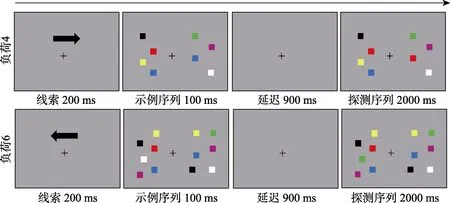
图1 负荷4和负荷6变化检测任务实验流程
脑电数据使用德国 Brain Product公司生产的BrainAmp 32导脑电系统采集。分别放置电极在右眼外侧记录水平眼电, 左眼下侧记录垂直眼电。头皮与电极之间的阻抗小于 10 kΩ, 在线滤波带通为0.05~100 Hz, 采样频率为1000 Hz/导。以FCz电极作为在线参考, 离线分析以左右两耳乳突电极的平均电位为参考电极, 离线低通滤波带宽 40 Hz。分别以示例序列和探测序列开始的时刻作为事件相关电位的零点, 示例序列和探测序列对应的分段时间分别为−200~800 ms 和−100~600 ms, 并分别取零点前的200 ms和100 ms脑电波作为基线。运动伪迹去除阈限为±100 μV, 水平眼电 HEOG的去除阈限为±70 μV。所有事件相关电位的分析只选取被试判断正确试次的脑电数据进行叠加平均。
不区分箭头朝向, 将所有试次记忆目标对侧和目标同侧的电极波幅的差值叠加平均(Hopf, Boelmans,Schoenfeld, Luck, & Heinze, 2004; Eimer & Kiss,2010)。N2pc是在刺激出现之后约180~280 ms出现在顶枕皮层的负波, 本研究中示例序列和探测序列N2pc的时间窗取得分别是示例序列和探测序列开始之后的180~250 ms和250~320 ms (Luck & Hillyard,1994a; Eimer, 1996; Woodman & Luck, 2003; Jolicœur,Brisson, & Robitaille, 2008; Woodman, Arita, &Luck, 2009; Woodman, 2010)。SPCN的时间窗取得分别是示例序列和探测序列开始后的320~800 ms和350~600 ms (Jolicœur et al., 2008; Eimer & Kiss,2010)。根据前人研究, N2pc和SPCN一般在PO7/PO8电极附近波幅最强, 由于本实验中的 32导电极帽不包括PO7/PO8, 因此选取这两个电极周围的三对电极CP5/6, P7/8, O1/2的对侧与同侧电极波幅差值的均值计算N2pc和 SPCN的幅值, 本研究并没有对N2pc和SPCN的潜伏期进行分析。
2.5 数据测量与分析
被试反应的辨别力通过信号检测论中的 d’表示, d’是击中率的标准分减去虚报率的标准分的差值, d’越大, 个体对刺激信号的辨别力越高。视觉工作记忆容量通过Cowan’s K (Cowan, 2001)系数表示, 其计算公式为:K = 记忆负荷 × (击中率 − 虚报率), 表示呈现特定数量的项目数时个体所能记忆的项目个数。对 K值, d’值以及事件相关电位 N2pc和SPCN的波幅值均采用2(刺激类型:正性电刺激右侧DLPFC, 虚假刺激) × 2(记忆负荷:负荷4, 负荷6)两因素重复测量方差分析, 采用SPSS 19.0进行数据处理。p值的多重比较校正均采用Bonferroni校正方法。
前人研究发现, 不同特质的个体对于电刺激的敏感性存在较大的差异, 个体本身工作记忆容量大小(Tseng et al., 2012)、性别(Meiron & Lavidor, 2013)甚至受教育程度(Berryhill & Jones, 2012)都会影响个体接受tDCS之后受益的大小。本研究设置了负荷4和负荷6两种任务, 由于人类视觉工作记忆容量大约为3~5个, 对有些被试来说, 负荷4任务已经接近他们的记忆限制, 负荷6任务则超出了他们的能力范围, 过多的记忆项目反而会干扰他们的记忆, 因此从负荷 4到负荷 6, 记忆容量不会提升甚至会降低。而对另一些个体来说, 负荷4还没有到达他们的记忆限制, 到负荷6时其记忆的项目数量会继续增加。虽然前人研究中关于这种记忆增长潜力的分组方式的描述较少, 但本研究中这种从负荷4到负荷6记忆容量提升的潜力大小与被试本身的平均记忆容量K (虚假刺激情况下负荷4和负荷6 K值的均值)也有相关,r= 0.65,p< 0.001。被试接受tDCS之后的增益量很有可能受到这种潜力的调节,因此我们根据被试在负荷4到负荷6任务情况下记忆容量的增加量大小(K6−K4)的中位数将所有被试分为两组:增加量大的高记忆增长潜力组(以下简称高潜力组)和增加量小甚至为负的低记忆增长潜力组(以下简称低潜力组)以探究工作记忆增长潜力对tDCS刺激右侧DLPFC作用的影响。
3 结果
3.1 行为结果
有2名被试由于在负荷6情况下反应正确率低于 50%被剔除, 对剩余 38名被试的实验数据进行了后续的分析。d’的分析结果表明:记忆负荷的主效应显著,F(1, 38) = 186.36,p< 0.001, ηp2= 0.83。刺激类型的主效应、刺激类型和记忆负荷的交互作用均不显著(ps > 0.05)。K的分析结果同d’结果相似, 记忆负荷的主效应显著,F(1, 38) = 18.83,p<0.001, ηp2= 0.34。刺激类型的主效应、刺激类型和记忆负荷的交互作用均不显著(ps> 0.05)。
所有被试的K值和d’分析结果表明, 视觉工作记忆的辨别力 d’会随着记忆负荷的提升显著降低(ps < 0.001), 而工作记忆的容量K则随着记忆负荷的提升显著提高(ps < 0.05)。但是正性tDCS刺激右侧DLPFC对视觉工作记忆的表现并没有明显的影响。为了进一步探究不同记忆增长潜力对电刺激效果的影响, 将组别变量:高潜力组(19人,MK6-K4=0.74,SDK6-K4= 0.40)、低潜力组(19人,MK6-K4=−0.10,SDK6-K4= 0.37)加入进行2(刺激类型:正性电刺激右侧DLPFC, 虚假刺激) × 2(记忆负荷:负荷4, 负荷6) × 2(组别:高潜力组, 低潜力组)重复测量方差分析发现, 刺激类型 × 记忆负荷 × 组别的交互作用显著, 无论是d’,F(2, 38) = 18.16,p<0.001, ηp2= 0.36; 还是 K,F(2, 38) = 10.30,p< 0.005,ηp2= 0.22。
分别对高、低潜力组被试的d’进行2刺激类型× 2记忆负荷重复测量方差分析表明(图2):低潜力组被试的记忆负荷主效应显著,F(1, 19) = 107.42,p<0.001, η2p= 0.86, 反应的辨别力随着记忆负荷的增加而降低。刺激类型的主效应、刺激类型和记忆负荷的交互作用均不显著(ps> 0.05), 电刺激并没有对低潜力组被试的表现产生影响。高潜力组被试d’的记忆负荷主效应显著,F(1, 19) = 80.59,p< 0.001,ηp2= 0.82; 刺激类型的主效应不显著,F(1, 19) =1.10,p> 0.05; 刺激类型和记忆负荷的交互作用显著,F(1, 19) = 19.39,p< 0.001, ηp2= 0.52。事后检验结果表明这种交互作用是由在负荷4的情况下正性电刺激右侧DLPFC相比于虚假刺激显著提升了被试的辨别力 d’导致,t(19) = 3.15,p< 0.01, Cohen’sd=0.58。而在负荷6时高潜力组被试在两种刺激情况下辨别力差异不显著,t(19) = 1.05,p> 0.05。两种刺激情况下, 两组在负荷4任务的辨别力都显著高于负荷 6 任务(ps < 0.005)。
高、低潜力组K的分析结果表明(图2):低潜力组被试的所有效应均不显著(ps > 0.05)。而高潜力组被试记忆负荷的主效应显著,F(1, 19) = 34.98,p< 0.001, ηp2= 0.66。刺激类型的主效应不显著,F(1,19) = 0.12,p> 0.05。刺激类型和记忆负荷的交互作用显著,F(1, 19) = 6.87,p< 0.05, ηp2= 0.28。事后检验结果表明这种交互作用是由在负荷4情况下正性电刺激右侧DLPFC相比于虚假刺激显著提升了高潜力组被试的记忆容量 K导致,t(19) = 2.13,p<0.05, Cohen’sd= 0.41。而在负荷6时两种刺激情况下的记忆容量差异不显著,t(19) = 1.38,p> 0.05。此外, 高潜力组被试在两种刺激情况下负荷6的记忆容量都显著高于负荷4的记忆容量(ps < 0.05)。
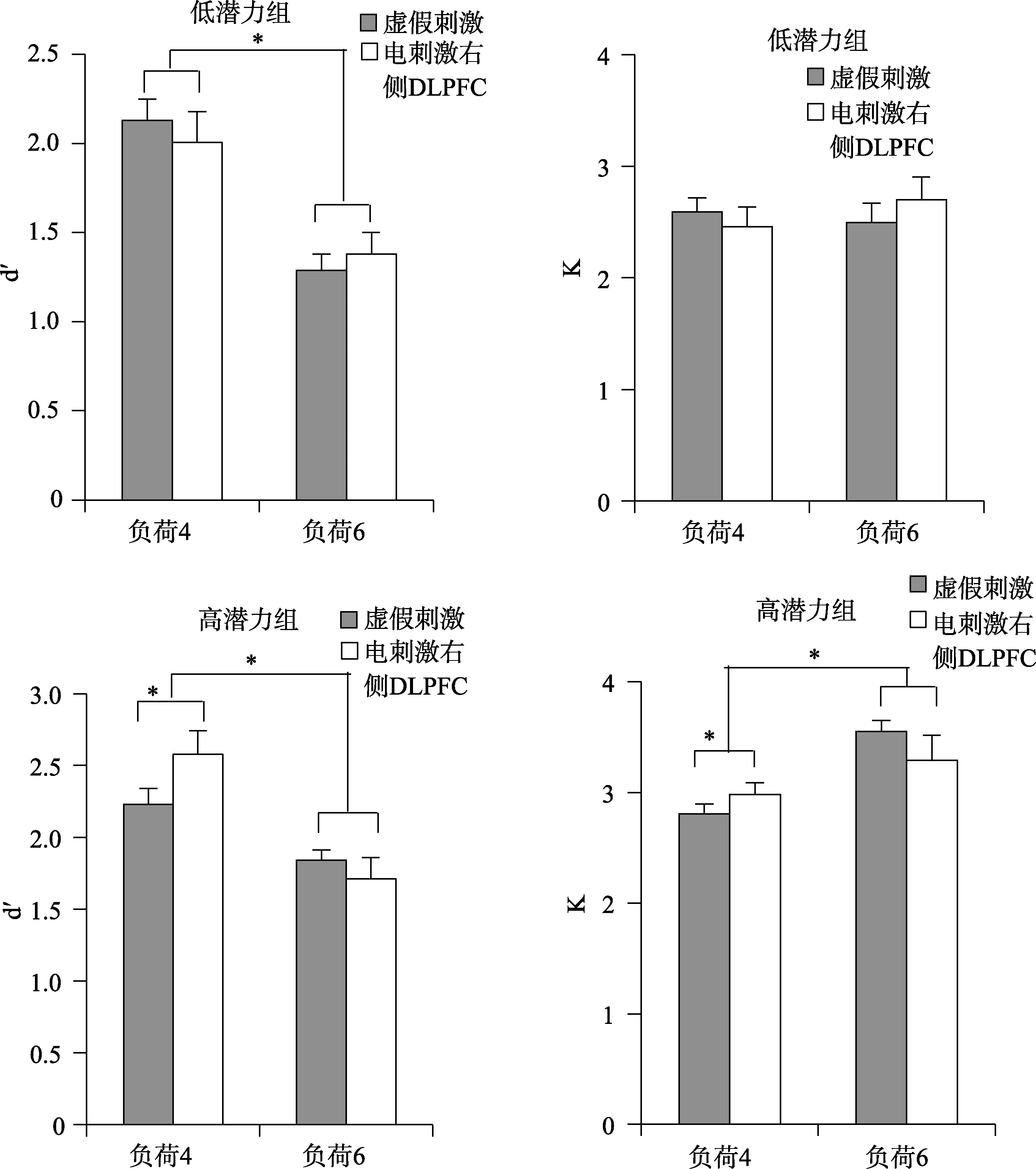
图2 高低潜力组行为表现
以上这些结果表明, 正性电刺激右侧 DLPFC对低潜力组被试的表现并没有显著影响, 但是会显著提升高潜力组被试在低记忆负荷下的辨别力和记忆容量。对被试在负荷4到负荷6记忆容量增长的潜力大小与正性电刺激对工作记忆表现 d’和 K的提升效果进行相关分析发现:被试的记忆容量提升潜力越大, 其在负荷4任务情况下行为表现获得电刺激的提升效果越大,rd’= 0.48,p< 0.01;rK=0.40,p< 0.05 (图 3)。
3.2 ERP 结果
一名被试由于眼动伪迹过多在后续的脑电数据分析中被剔除, 对剩余的 37名被试的脑电数据进行分析。选取CP5/CP6, P7/P8, O1/O2三对电极的对侧电极减同侧电极幅值的差值均值进行分析。对 37名被试在示例序列出现后的 N2pc波幅和SPCN波幅进行2刺激类型 × 2记忆负荷重复测量方差分析结果表明(图4):N2pc波幅和SPCN波幅的所有效应均不显著(ps > 0.05)。对37名被试在探测序列出现后的N2pc波幅和SPCN波幅进行2 × 2重复测量方差分析结果表明(图 5, 图 6):N2pc波幅的所有效应均不显著(ps > 0.05)。SPCN波幅的刺激类型主效应显著,F(1, 37) = 6.45,p< 0.05, ηp2=0.15, 记忆负荷的主效应、刺激类型和记忆负荷的交互作用均不显著(ps > 0.05)。对不同刺激情况下SPCN的波幅进行配对样本t检验, 结果表明, 负荷4情况下, 电刺激右侧DLPFC之后SPCN的幅值比虚假刺激情况下更负,t(37) = −0.23,p= 0.04,Cohen’sd= 0.74。而负荷6情况下二者差异不显著。加入组别因素对N2pc波幅和SPCN波幅进行2刺激类型 × 2记忆负荷 × 2组别重复测量方差分析,结果表明:N2pc波幅的各类效应均不显著; SPCN波幅的刺激类型主效应显著,F(1, 37) = 7.01,p<0.05, ηp2= 0.167, 其他效应均不显著。
虽然刺激类型 × 记忆负荷 × 组别的交互作用并不显著, 为了进一步探究电刺激对工作记忆神经指标的影响是否受到被试个体差异的调节, 分别对高潜力组(18人)和低潜力组(19人)被试的 N2pc波幅和SPCN波幅进行2刺激类型 × 2记忆负荷重复测量方差分析。结果表明:高、低潜力组被试N2pc波幅的重复测量方差分析结果均不显著。低潜力组被试 SPCN波幅的各种效应均不显著(ps >0.05) (图 5, 图6)。高潜力组被试的SPCN波幅的刺激类型主效应显著,F(1, 18) = 8.23,p< 0.05, ηp2=0.33)。但是记忆负荷的主效应、刺激类型与记忆负荷的交互作用均不显著(ps > 0.05)。配对样本t检验结果表明, 高潜力组被试在负荷 4任务情况下, 正性电刺激右侧 DLPFC相对于虚假刺激情况下的SPCN波幅获得显著提升,t(18) = −3.27,p< 0.01,Cohen’sd= 0.78。而在负荷6的情况下二者差异不显著,t(18) = −1.42,p> 0.05。
对被试在负荷4到负荷6记忆容量增长的潜力大小与正性电刺激对SPCN波幅的提升效果进行相关分析发现:被试的记忆容量提升潜力越大, 其在负荷4任务情况下SPCN波幅获得电刺激的提升效果越大,r= −0.39,p= 0.017 (图7), 与行为结果一致。
4 讨论
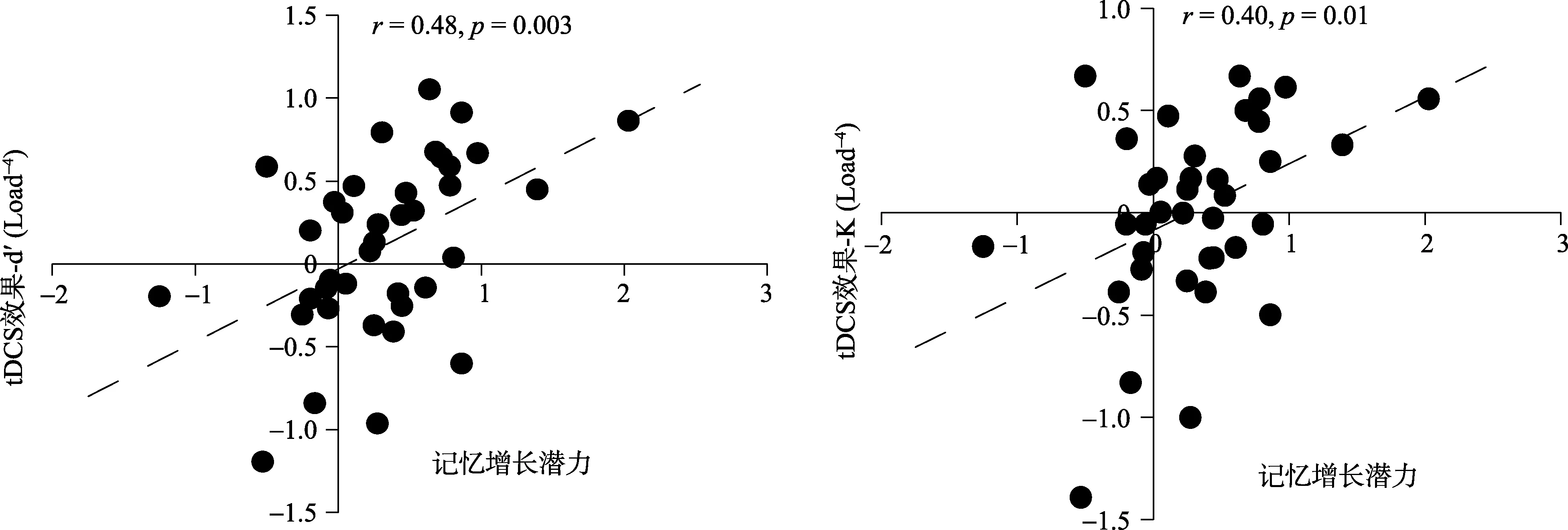
图3 记忆增长潜力和正性电刺激右侧DLPFC效果的相关
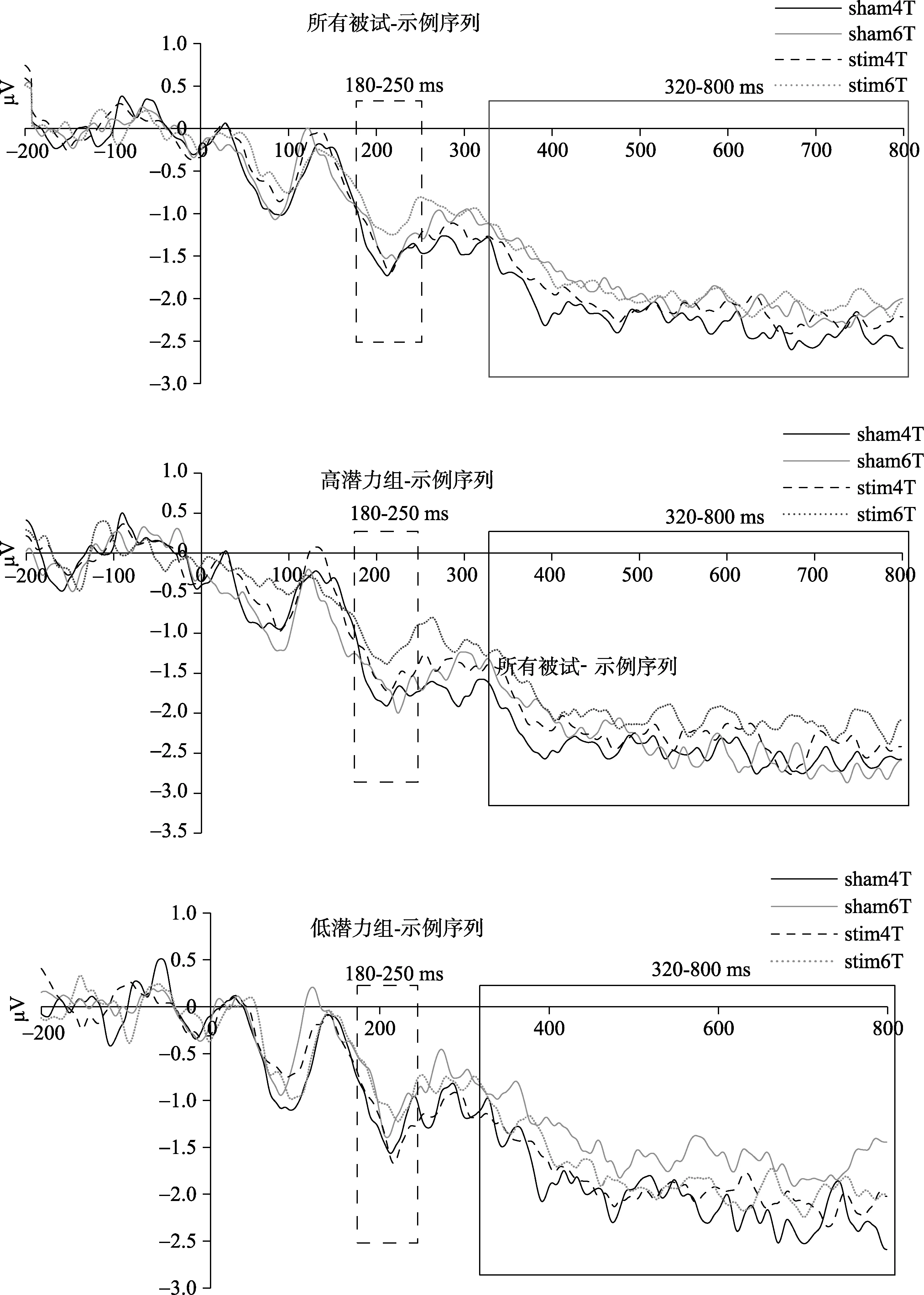
图4 所有被试以及高低潜力组被试在示例序列出现后的ERP成分
本研究通过正性电刺激右侧DLPFC提升其兴奋性以实现对视觉工作记忆行为表现和神经活动的调控, 从而验证右侧 DLPFC在不同负荷视觉工作记忆加工中的因果性作用。根据被试在控制条件(虚假刺激)下从负荷4到负荷6任务时记忆容量的增长潜力将被试分为高增长潜力组和低增长潜力组, 发现正性电刺激刺激右侧 DLPFC能够选择性地提升高增长潜力组个体在低记忆负荷下的视觉工作记忆表现以及信息提取阶段的SPCN波幅。证明了右侧DLPFC在视觉工作记忆的提取阶段发挥重要作用。
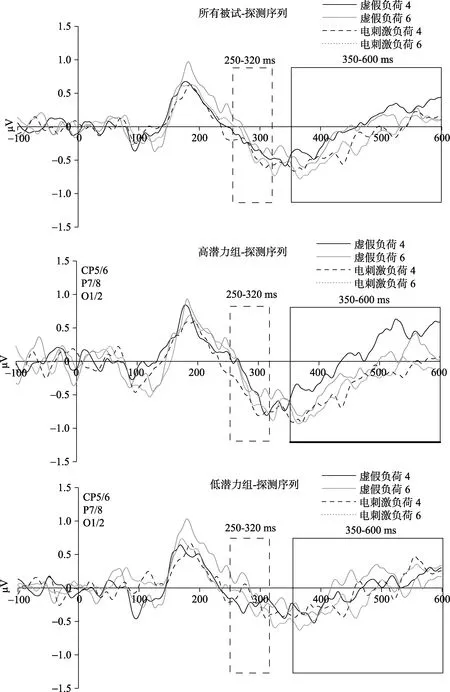
图5 所有被试以及高低潜力组被试在探测序列出现后的ERP成分
同前人研究结果相一致, 本研究发现正性经颅直流电刺激的刺激效果与个体本身的特质存在交互作用。Tseng等人(2012)的研究使用正性经颅直流电刺激右侧后顶叶皮层(posterior parietal cortex)发现本身工作记忆容量较低的个体相对于工作记忆容量较高的个体更易获得电刺激的提升作用。Berryhill等人(2012)分别电刺激左侧和右侧DLPFC发现受教育程度较高的老年人相比于受教育程度较低的老年人更易获得电刺激提升效果。Meiron等人(2013)的研究结果表明女性更容易从电刺激右侧DLPFC中受益, 而男性更易从电刺激左侧 DLPFC中受益。因此, 从电刺激中受益的程度与很多因素有关。虽然Tseng等人的研究发现工作记忆容量较低的个体更易从电刺激中获益, 但是我们的研究却发现工作记忆容量增长潜力较高的个体从tDCS中获得了更多提升。其原因可能在于, 尽管我们采用了相似的任务(Vogel & Machizawa, 2004), 但是我们电刺激的脑区并不相同, Tseng等人刺激的是右侧PPC (Posterior Parietal Cortex), 该脑区更多的在工作记忆的信息维持阶段发挥作用, 主要负责视觉空间注意和记忆维持。而本研究刺激的脑区是右侧DLPFC, 则更多的发挥执行控制的作用, 主要负责目标维持和干扰抑制。这两个脑区兴奋性的变化可能对视觉工作记忆的表现造成不同的影响。而且最新的研究发现电刺激的效果并不受到个体的工作记忆容量大小的影响(Li et al., 2017), 这表明电刺激的效果并不受单一因素的影响。一项关于 tDCS对运动和认知表现调控的元分析研究表明, tDCS对行为表现的影响效果确实很难预测(Jacobson,Koslowsky, & Lavidor, 2012)。同 Berryhill等人(2012)的研究结果发现受教育程度高的群体电刺激获益大相类似, 本研究发现tDCS刺激右侧DLPFC提升了高潜力组的表现。本研究发现电刺激的效果也会受到个体本身从较低负荷到较高负荷工作记忆容量的增长潜力的影响, 同前人研究结果共同证明了个体对于电刺激的敏感性可能受到个体本身神经系统特性的调控。Nitsche, Liebetanz等人(2003)提出经颅直流电刺激的长时效果依赖于大脑中NMDA谷氨酸受体介导的长时程增强/抑制作用。他们还发现经颅直流电刺激的神经可塑性效果受到脑源性神经营养因子(BDNF)基因型多样性的影响。因此个体电刺激效果的差异很有可能是不同的受体和基因型调控导致的。
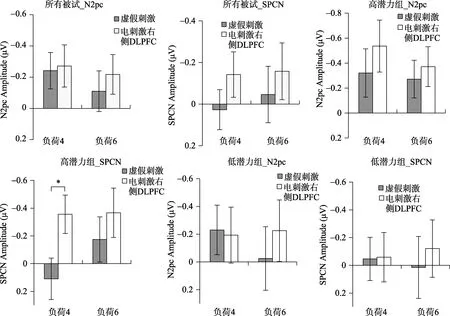
图6 所有被试、高低潜力组被试探测序列出现后N2pc (250~320 ms)和SPCN (350~600 ms)的均值
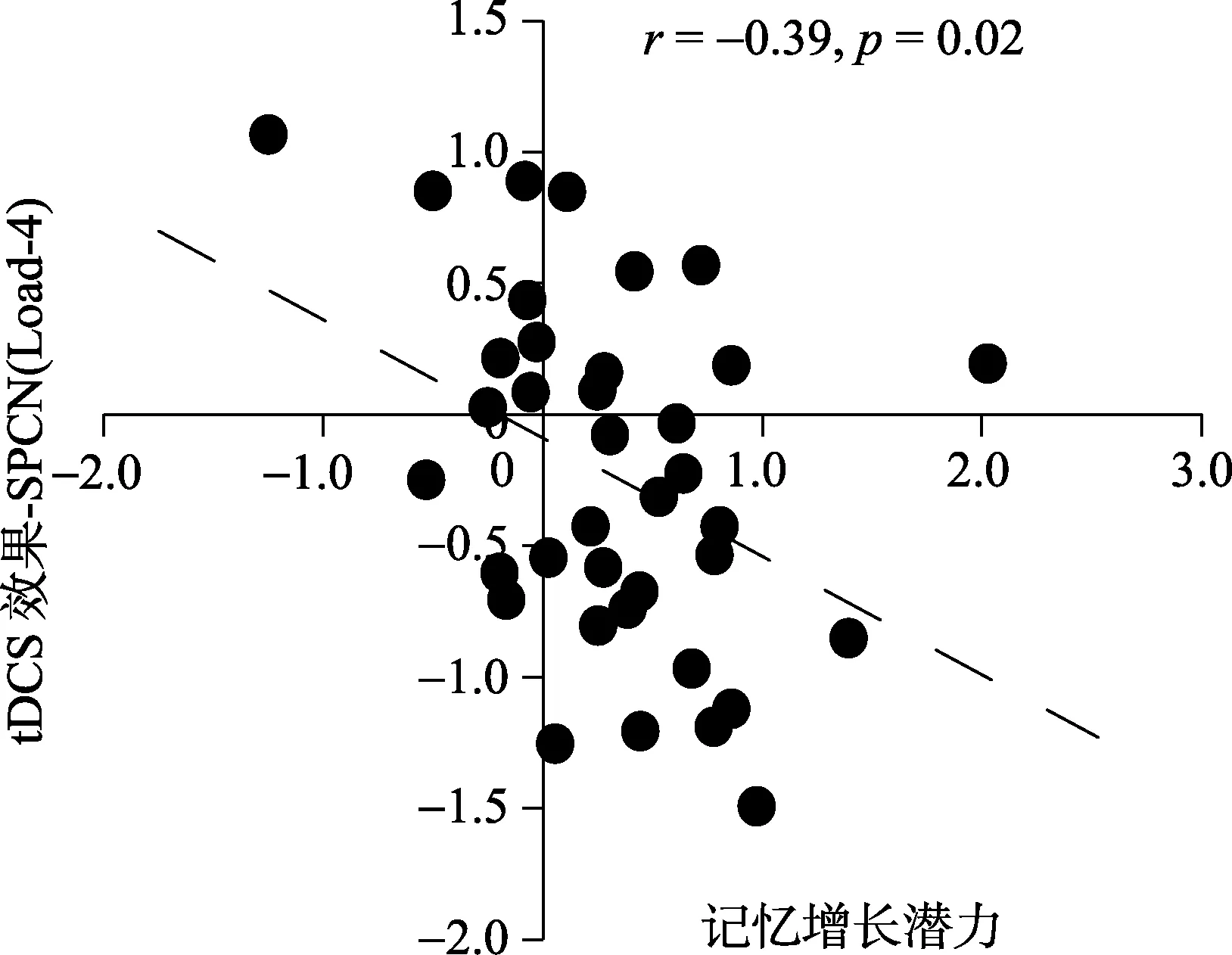
图7 被试的记忆增长潜力与正性电刺激右侧DLPFC在SPCN幅值上效果的皮尔逊相关
本研究发现, 正性电刺激右侧 DLPFC选择性地提升了高潜力组被试在低负荷任务下的视觉工作记忆辨别力和容量, 而对于高负荷任务的表现没有影响。对于低潜力组被试来说, 负荷4已经到达他们的记忆限制, 即使通过正性电刺激提升右侧DLPFC的兴奋性也无法使他们的记忆表现获得提升; 而对于高潜力组被试, 在负荷 4的情况下他们的工作记忆容量还具有较高的提升潜力, 提高DLPFC活动水平能够促进他们的记忆表现; 当负荷达到6时, 高潜力组被试也到达了自己的记忆限制, 因此即使 DLPFC活动水平增强也无法对他们的工作记忆表现起到促进作用。这些结果表明:电刺激右侧DLPFC对工作记忆表现的影响效果受到记忆负荷和难度的调节, 证明了 DLPFC的活动具有记忆负荷依赖的特性(Druzgal & D'Esposito, 2003;Rypma et al., 2002)。并且同Tseng等人(2012)的研究结果:tDCS没有对高表现组的视觉工作记忆产生影响一致证明, DLPFC的活动水平对工作记忆表现的调控作用存在一个限制阈限, 当它的活动达到这一限制后, 即使通过外界调控(电刺激等)增强其活动水平, 也不会对工作记忆的表现产生影响。
前人的研究虽然在一定程度上证明了右侧DLPFC在视觉空间工作记忆中的作用(Wu et al.,2014; Giglia et al., 2014), 但缺乏对其具体神经机制的探究, 本研究采用能够区分不同加工阶段的范式, 不同的ERP成分N2pc和SPCN对电刺激右侧DLPFC调控视觉工作记忆表现的神经机制进行深入探索。前人研究表明, SPCN反映的是视觉工作记忆的信息维持, 但是这种信息维持可能发生在工作记忆的延迟保持阶段(Vogel & Machizawa, 2004),也可能发生在提取阶段(Tseng et al., 2012)。本研究并未发现延迟和提取过程中SPCN的负荷效应, 这一结果与前人研究相符(Robitaille et al., 2010; Vogel& Machizawa, 2004), 超过记忆容量(4)之后的顶叶对侧−同侧差异波的波幅维持不变。有意思的是,本研究发现延迟过程中的SPCN不受到刺激的影响,而提取过程的 SPCN在特定情况下受到刺激的影响。这表明相对于工作记忆的编码和保持阶段, 右侧DLPFC在提取阶段的作用更加显著。而提取过程中 N2pc没有发现显著结果, 从神经生理学角度证明在视觉工作记忆的提取阶段, 相比于早期注意,右侧DLPFC可能在晚期比较过程的信息维持中发挥更显著的作用。
最后, 我们的研究为大脑右半球在视觉空间工作记忆加工中具有加工优势提供了进一步的证据。前人研究证明了右侧DLPFC与工作记忆的干扰抑制(Wu et al., 2014; Li et al., 2017), 和延迟再认(Giglia et al., 2014)的因果联系, 通过采用事件相关电位的分析方法, 本研究为 DLPFC在工作记忆的提取过程中的重要作用提供了因果联系的证据。除此之外, 我们的研究发现也为找到提升个体工作记忆的表现的快速有效方式提供了良好的启示。
参 考 文 献
Alloway, T. P., & Alloway, R. G. (2010). Investigating the predictive roles of working memory and IQ in academic attainment.Journal of Experimental Child Psychology,106(1), 20–29.
Anderson, M. C., Ochsner, K. N., Kuhl, B., Cooper, J.,Robertson, E., Gabrieli, S. W., ... Gabrieli, J. D. E. (2004).Neural systems underlying the suppression of unwanted memories.Science,303(5655), 232–235.
Andrews, S. C., Hoy, K. E., Enticott, P. G., Daskalakis, Z. J.,& Fitzgerald, P. B. (2011). Improving working memory:The effect of combining cognitive activity and anodal transcranial direct current stimulation to the left dorsolateral prefrontal cortex.Brain Stimulation, 4(2), 84–89.
Baddeley, A. D., & Hitch, G. J. (1974). Working memory.Psychology of Learning and Motivation,8, 47–89.
Berryhill, M. E., & Jones, K. T. (2012). tDCSselectivelyimproves working memory in older adults with more education.Neuroscience Letters,521(2), 148–151.
Coffman, B. A., Clark, V. P., & Parasuraman, R. (2014). Battery powered thought: Enhancement of attention, learning, and memory in healthy adults using transcranial direct current stimulation.NeuroImage,85, 895–908.
Courtney, S. M., Petit, L., Maisog, J. M., Ungerleider, L. G.,& Haxby, J. V. (1998). An area specialized for spatial working memory in human frontal cortex.Science,279(5355),1347–1351.
Cowan, N. (2001). Metatheory of storage capacity limits.Behavioral and Brain Sciences, 24(1), 154–176.
Curtis, C. E. (2006). Prefrontal and parietal contributions to spatial working memory.Neuroscience,139(1), 173–180.
Dedoncker, J., Brunoni, A. R., Baeken, C., & Vanderhasselt,M. A. (2016). A systematic review and meta-analysis of the effects of transcranial direct current stimulation (tDCS)over the dorsolateral prefrontal cortex in healthy and neuropsychiatric samples: Influence of stimulation parameters.Brain Stimulation,9(4), 501–517.
D'Esposito, M., Postle, B. R., Ballard, D., & Lease, J. (1999).Maintenance versus manipulation of information held in working memory: An event-related fMRI study.Brain and Cognition,41(1), 66–86.
Druzgal, T. J., & D'Esposito, M. (2003). Dissecting contributions of prefrontal cortex and fusiform face area to face working memory.Journal of Cognitive Neuroscience,15(6), 771–784.
Eimer, M. (1996). The N2pc component as an indicator of attentional selectivity.Electroencephalography and Clinical Neurophysiology,99(3), 225–234.
Eimer, M., & Kiss, M. (2010). An electrophysiological measure of access to representations in visual working memory.Psychophysiology,47(1), 197–200.
Fregni, F., Boggio, P. S., Nitsche, M., Bermpohl, F., Antal, A.,Feredoes, E., ... Pascual-Leone, A. (2005). Anodal transcranial direct current stimulation of prefrontal cortex enhances working memory.Experimental Brain Research,166(1),23–30.
Fukuda, K., Vogel, E., Mayr, U., & Awh, E. (2010). Quantity,not quality: The relationship between fluid intelligence and working memory capacity.Psychonomic Bulletin & Review,17(5), 673–679.
Funahashi, S., Bruce, C. J., & Goldman-Rakic, P. S. (1989).Mnemonic coding of visual space in the monkey's dorsolateral prefrontal cortex.Journal of Neurophysiology,61(2), 331–349.
Fuster, J. M., & Alexander, G. E. (1971). Neuron activity related to short-term memory.Science,173(3997), 652–654.
Giglia, G., Brighina, F., Rizzo, S., Puma, A., Indovino, S.,Maccora, S., ... Fierro, B. (2014). Anodal transcranial direct current stimulation of the right dorsolateral prefrontal cortex enhances memory-guided responses in a visuospatial working memory task.Functional Neurology, 29(3),189–193.
Hopf, J. M., Boelmans, K., Schoenfeld, M. A., Luck, S. J., &Heinze, H. J. (2004). Attention to features precedes attention to locations in visual search: Evidence from electromagnetic brain responses in humans.Journal of Neuroscience,24(8), 1822–1832.
Jacobson, L., Koslowsky, M., & Lavidor, M. (2012). tDCS polarity effects in motor and cognitive domains: A metaanalytical review.Experimental Brain Research,216(1),1–10.
Jolicœur, P., Brisson, B., & Robitaille, N. (2008). Dissociation of the N2pc and sustained posterior contralateral negativity in a choice response task.Brain Research,1215, 160–172.
Jolicœur, P., Sessa, P., Dell’Acqua, R., & Robitaille, N.(2006). On the control of visual spatial attention: Evidence from human electrophysiology.Psychological Research,70(6), 414–424.
Jones, K. T., & Berryhill, M. E. (2012). Parietal contributions to visual working memory depend on task difficulty.Frontiers in Psychiatry,3, 81.
Klaver, P., Talsma, D., Wijers, A. A., Heinze, H. J., & Mulder,G. (1999). An event-related brain potential correlate of visual short-term memory.NeuroReport,10(10), 2001–2005.
Kubota, K., & Niki, H. (1971). Prefrontal cortical unit activity and delayed alternation performance in monkeys.Journal of Neurophysiology,34(3), 337–347.
Li, S. Y., Cai, Y., Liu, J., Li, D. W., Feng, Z. F., Chen, C. S.,& Xue, G. (2017). Dissociated roles of the parietal and frontal cortices in the scope and control of attention during visual working memory.NeuroImage,149, 210–219.
Luck, S. J., & Hillyard, S. A. (1994a). Spatial filtering during visual search: Evidence from human electrophysiology.Journal of Experimental Psychology: Human Perception and Performance,20(5), 1000–1014.
Luck, S. J., & Hillyard, S. A. (1994b). Electrophysiological correlates of feature analysis during visual search.Psychophysiology,31(3), 291–308.
Luck, S. J., & Vogel, E. K. (1997). The capacity of visual working memory for features and conjunctions.Nature,390(6657), 279–281.
Mazza, V., & Caramazza, A. (2012). Perceptual grouping and visual enumeration.PLoS One,7(11), e50862.
Meiron, O., & Lavidor, M. (2013). Unilateral prefrontal direct current stimulation effects are modulated by working memory load and gender.Brain Stimulation,6(3), 440–447.
Nitsche, M. A., & Paulus, W. (2001). Sustained excitability elevations induced by transcranial DC motor cortex stimulation in humans.Neurology,57(10), 1899–1901.
Nitsche, M. A., Fricke, K., Henschke, U., Schlitterlau, A.,Liebetanz, D., Lang, N., ... Paulus, W. (2003).Pharmacological modulation of cortical excitability shifts induced by transcranial direct current stimulation in humans.The Journal of Physiology,553(1), 293–301.
Nitsche, M. A., Liebetanz, D., Lang, N., Antal, A., Tergau, F.,& Paulus, W. (2003). Safety criteria for transcranial direct current stimulation (tDCS) in humans.Clinical Neurophysiology,114(11), 2220–2222.
Ohn, S. H., Park, C. I., Yoo, W. K., Ko, M. H., Choi, K. P.,Kim, G. M., ... Kim, Y. H. (2008). Time-dependent effect of transcranial direct current stimulation on the enhancement of working memory.NeuroReport,19(1),43–47.
Priori, A. (2003). Brain polarization in humans: A reappraisal of an old tool for prolonged non-invasive modulation of brain excitability.Clinical Neurophysiology,114(4), 589–595.
Robitaille, N., Marois, R., Todd, J., Grimault, S., Cheyne, D.,& Jolicœur, P. (2010). Distinguishing between lateralized and nonlateralized brain activity associated with visual short-term memory: fMRI, MEG, and EEG evidence from the same observers.NeuroImage,53(4), 1334–1345.
Rypma, B., Berger, J. S., & D'Esposito, M. (2002). The influence of working-memory demand and subject performance on prefrontal cortical activity.Journal of Cognitive Neuroscience,14(5), 721–731.
Smith, E. E., Jonides, J., & Koeppe, R. A. (1996). Dissociating verbal and spatial working memory using PET.Cerebral Cortex,6(1), 11–20.
Todd, J. J., & Marois, R. (2004). Capacity limit of visual short-term memory in human posterior parietal cortex.Nature,428(6984), 751–754.
Toepper, M., Gebhardt, H., Beblo, T., Thomas, C., Driessen,M., Bischoff, M., ... & Sammer, G. (2010). Functional correlates of distractor suppression during spatial working memory encoding.Neuroscience,165(4), 1244–1253.
Tseng, P., Hsu, T. Y., Chang, C. F., Tzeng, O. J. L., Hung, D.L., Muggleton, N. G., ... & Juan, C. H. (2012). Unleashing potential: Transcranial direct current stimulation over the right posterior parietal cortex improves change detection in low-performing individuals.Journal of Neuroscience,32(31),10554–10561.
Vanderhasselt, M. A., De Raedt, R., & Baeken, C. (2009).Dorsolateral prefrontal cortex and Stroop performance:Tackling the lateralization.Psychonomic Bulletin & Review,16(3), 609–612.
Vogel, E. K., & Machizawa, M. G. (2004). Neural activity predicts individual differences in visual working memory capacity.Nature,428(6984), 748–751.
Vogel, E. K., McCollough, A. W., & Machizawa, M. G.(2005). Neural measures reveal individual differences in controlling access to working memory.Nature,438(7067),500–503.
Walter, H., Bretschneider, V., Grön, G., Zurowski, B.,Wunderlich, A. P., Tomczak, R., & Spitzer, M. (2003).Evidence for quantitative domain dominance for verbal and spatial working memory in frontal and parietal cortex.Cortex,39(4-5), 897–911.
Woodman, G. F. (2010). A brief introduction to the use of event-related potentials in studies of perception and attention.Attention, Perception, & Psychophysics,72(8),2031–2046.
Woodman, G. F., Arita, J. T., & Luck, S. J. (2009). A cuing study of the N2pc component: An index of attentional deployment to objects rather than spatial locations.Brain Research,1297, 101–111.
Woodman, G. F., & Luck, S. J. (2003). Serial deployment of attention during visual search.Journal of Experimental Psychology: Human Perception and Performance,29(1),121–138.
Wu, Y. J., Tseng, P., Chang, C. F., Pai, M. C., Hsu, K. S., Lin,C. C., & Juan, C. H. (2014). Modulating the interference effect on spatial working memory by applying transcranial direct current stimulation over the right dorsolateral prefrontal cortex.Brain and Cognition,91, 87–94.
