近日钟基因Clock单核苷酸多态性与心肌缺血的 相关性*
2018-07-06陈晨江舟王正荣李光明
陈晨 江舟 王正荣 李光明
(1.四川省科学城医院,四川 绵阳621900;2.四川大学华西基础医学院与法医学院生物医学工程研究室,四川 成都 610041; 3. 卫生部时间生物学重点实验室,四川 成都610041;4.南充市中心医院,四川 南充637000)
缺血性心脏病(ischemic heart disease)是指冠状动脉粥样硬化导致血管腔狭窄或闭塞,或(和)因为冠状动脉功能性改变(即血管痉挛)所致心肌缺血缺氧,严重致心肌坏死而引起的心脏疾病[1-2]。流行病学调查发现近年我国缺血性心脏病发病率逐年升高,严重危害人类健康的常见病[3- 4]。本病出现临床症状或致残、致死等多发生于40岁以后,且男性发病明显早于女性患者。缺血性心脏病的发病机制,曾有多种学说从不同的角度加以阐述,从最早的脂质浸润学说,其后的平滑肌细胞克隆学说以及血小板聚集和血栓形成学说。现在学术界倾向于1973年提出的动脉粥样硬化形成的“损伤-反应学说”。
近年随着时间生物学的兴起,众多学者及临床医务工作者在长期的医学实践观察中发现心肌缺血发病具有近日节律,心肌缺血的节律被认为与心肌电生理、血液流变学、自主神经活动和冠脉血栓形成等的近日节律有关[5-7]。Maemura K等[8-9]采用酵母双杂交技术对人脐带血管内皮细胞cDNA进行分析,发现节律基因Clif(aryl hydrocarbon receptor nuclear translocator like 2,CLIF)能与近日钟基因Clock(circadian clock gene)结合形成二聚体,在内皮细胞中Clock、Clif形成的二聚体能与血浆纤溶蛋白溶酶原激活抑制因子-1(serpin family E member 1,Pai-1)基因的E-box结合促进其上调表达产生Pai-1,形成近日节律,这也是心肌梗塞或心绞痛常见于早晨的关键因素。为进一步明确近日钟基因在心肌缺血发生发展中的作用及机制,本研究致力于中国汉族人群中Clock基因多态性与心肌缺血的相关性研究,拟对心肌缺血患者Clock基因多态性进行多位点和多样本综合研究,有助于更好地了解心肌缺血发病的机制,进而为缺血性心脏病的预防提供实验依据,对已患患者群更有针对性地制定个体化治疗方案。
1 资料与方法
1. 1 一般资料 收集2015年1月~2016年1月之间临床确诊的心肌缺血患者120例设为心肌缺血组,年龄38~84岁;同时通过临床体检确定为健康人群78例设为健康对照组,年龄40~75岁(样本例数来自四川省科学城医院、南充市中心医院及四川大学华西基础医学部)。所有实验标本的收集均经本医院的医学伦理委员会认可并获得患者知情同意。分别于清晨7时抽取5ml抗凝血液进行生化指标检测包括甘油三酯、总胆固醇、高密度脂蛋白、低密度脂蛋白等指标,其中3ml留作DNA分离进行基因测定。总胆固醇、甘油三酯及低密度脂蛋白指标心肌缺血组明显高于健康对照组,两者比较差异有统计学意义(P<0.05);而高密度脂蛋白指标在心肌缺血组与健康对照组相比差异无统计学意义(P>0.05)。心肌缺血组与健康对照组临床资料统计分析,见表1。
1.2 试验试剂和相关仪器 普通 PCR 仪、电泳仪及凝胶成像系统购自美国BIO-RAD公司,NanoDrop 2000c 紫外分光光度计购自美国ThermoScientific公司,电热恒温水浴箱购自北京永光明医疗仪器厂。总 RNA 抽提试剂 Trizol 购自 Invitrogen 公司。逆转录试剂盒,普通PCR试剂盒(Takara EmeraldAmp PCR Master Mix)与人全基因组DNA提取试剂盒(Universal Genomic DNA Extraction Kit Ver.3.0)购自日本Takara公司。SNaPshot试剂盒购自美国ABI公司。Pai-1和血栓素(thrombomodulin,Tm)酶联免疫吸附试验试剂盒购自美国Sigma公司。近日钟基因(clock circadian regulator,Clock)干扰试剂盒购自中国上海吉玛公司。

Table2Comparisonofgeneralandbiochemicalindexesofmyocardialischemiagroupandhealthycontrolgroup

组别心肌缺血组(n=120)健康对照组(n=78)P年龄 (岁)67.2 ± 8.959.8 ± 10.00.022①性别 (女/男)49/7145/330.021①总胆固醇(mmol/L)4.49 ± 1.032.84 ± 0.760.005①甘油三酯(mmol/L)1.92 ± 1.561.23 ± 0.660.002①高密度脂蛋白(mmol/L)1.91 ± 0.851.36 ± 0.310.170低密度脂蛋白(mmol/L)3.75 ± 0.882.78 ± 0.63<0.001①
注:与健康对照组比较,①P<0.05
1.3 方法
1.3.1 人全基因组DNA的提取 选取-80℃保存的血细胞,依照Universal Genomic DNA Extraction Kit Ver.3.0试剂盒说明书,提取各标本中的全基因组DNA,之后使用NanoDrop 2000c 紫外分光光度计测定所提取每个样本的全基因组浓度,并放于-80℃超低温冰箱保存备用。
1.3.2 小鼠总RNA提取及逆转录 收集各组小鼠的血浆,按照 RNA 抽提试剂盒说明书抽提总 RNA。用NanoDrop 2000c 紫外分光光度计对提取的总RNA 进行定量后,取1 ug总RNA按照试剂盒说明书进行逆转录反应(reverse transcription,RT )。
1.3.3 普通PCR反应 按照Takara EmeraldAmp PCR Master Mix试剂盒进行普通PCR反应。反应体系如下:PCR Master Mix 10μL,正、反向引物(10 μmol )共2.0μL,模板1μL,灭菌水7μL,引物序列(见表2)。PCR 扩增条件如下:98℃预变性2 min,98℃变性10 s,60℃退火30 s,72℃延伸60 s,共37个循环,72℃延伸5 min,4℃冷却30 min,反应结束后置4℃保存。随后采用1.5 %琼脂糖电泳检测PCR产物大小,取出凝胶在凝胶成像系统记录并鉴定提取结果。
1.3.4 酶联免疫吸附试验(ELISA) 采用双抗体夹心法测定人群和小鼠血浆中PAI-1和血栓素(TM)蛋白含量的变化表达水平。主要原理如下:用纯化的一抗包被微孔板,制成固相抗体,往微孔中加入血清,再加入与酶标记过的二抗结合,形成抗体-抗原-酶标抗体复合物,经过彻底洗涤后加显色剂显色。

表2 基因引物序列信息 Table 2 Sequence information of gene primers
1.3.5 RNA干扰技术 C57BL/6J 小鼠购自南京动物模式研究,参照Clock干扰试剂盒,采用小鼠尾静脉注射干扰物,沉默小鼠Clock基因的表达,并采用RT-PCR测定小鼠血浆Clock mRNA的表达水平变化。
1.3.6 单碱基引物延伸法基因分型(SnapShot) 按照SnapShot试剂盒进行反应。反应体系如下:PCR产物总体积9μL,Exol酶0.1μL,SAP buffer 0.1μL,SAP 2μL,加灭菌水至总体积12μL。反应条件:37℃ 1 h;75℃ 15min。随后进行单碱基引物延伸,反应体系:纯化后产物1μL,SNapShot Multiplex 1μL,SnapShot primer 0.2μL,加灭菌水至至总体积5μL。反应条件:96℃ 预变性1min;96℃变性10 sec,50℃退火5 sec,60℃延伸30 sec,共25个循环。随后采用1 %琼脂糖电泳检测PCR产物大小,取出凝胶在凝胶成像系统记录并鉴定提取结果。

2 结果
2.1 两组患者Clock基因多态性位点基因型及等位基因频率分布比较Clock基因rs3840267位点的基因型及等位基因频率分布,在心肌缺血组和健康对照组中差异有统计学意义(P<0.05)。而rs3749474和rs1402963位点基因型及等位基因频率分布,在心肌缺血组和健康对照组比较,差异无统计学意义(P>0.05),见表3。

表3 两组患者Clock基因多态性位点基因型及等位基因频率分布比较[n(×10-2 )] Table 3 Comparison of genotype and allele frequency distribution of Clock gene polymorphic loci in myocardial ischemia group and healthy control group
注:与健康对照组相比,①P<0.05
2.2 两组Clock基因rs3840267位点基因型及等位基因频率分布在不同性别之间的比较 心肌缺血组与健康对照组中Clock基因rs3840267位点的基因表型和等位基因分频率分布在心肌缺血组与健康对照组中不同性别之间的比较,发现,在女性组中心肌缺血组AA和A型显著性的低于健康对照组(P<0.05),见表4、表5。
表4Clock基因rs3840267位点基因型及等位基因频率分布在两组中男性人群的比较[n(×10-2)]
Table4ComparisonofgenotypeandallelefrequencydistributionofClockgeners3840267lociinthemalegroupofmyocardialischemiagroupandhealthycontrolgroup

组别心肌缺血组健康对照组P风险比(95%可信区间)AA54(87.10)27(87.10)11A-8(12.90)4(12.90)-0(0.00)0(0.00)A116(93.55)58(93.55)11-8(6.45)4(6.45)
表5Clock基因rs3840267位点基因型及等位基因频率分布在两组中女性人群的比较[n(×10-2)]
Table5ComparisonofgenotypeandallelefrequencydistributionofClockgeners3840267lociinwomenwithmyocardialischemiaandhealthycontrols
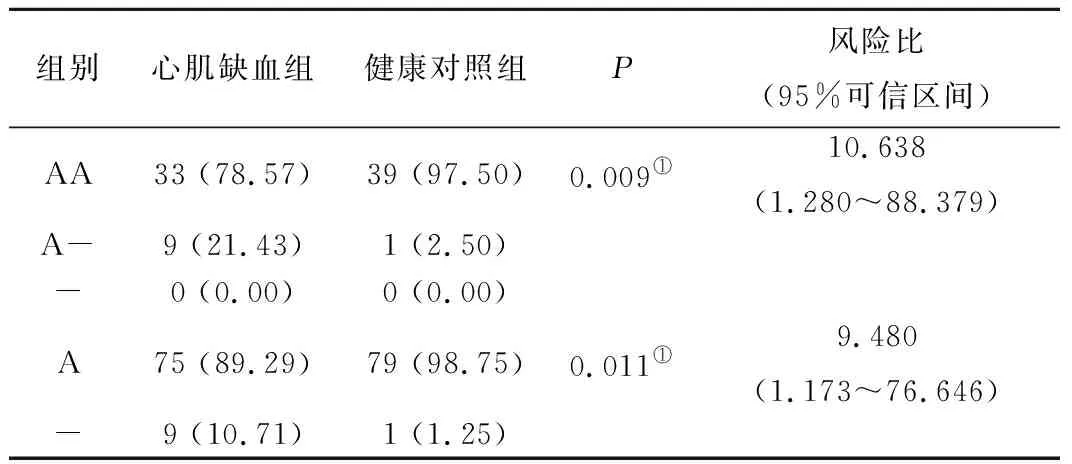
组别心肌缺血组健康对照组P风险比(95%可信区间)AA33 (78.57)39 (97.50)0.009①10.638(1.280~88.379)A-9 (21.43)1 (2.50)-0 (0.00)0 (0.00)A75 (89.29)79 (98.75)0.011①9.480(1.173~76.646)-9 (10.71)1 (1.25)
注:与健康对照组相比,①P<0.05
3 讨论
心血管系统广泛存在着近日节律的变化,无论是心率、血压、心功能、心肌收缩力等均呈现出近日节律变化的规律,这些节律存在是由近日节律钟基因的存在通过其自激振荡而调控心血管系统而形成的心血管系统的近日节律[10-12]。通常心血管系统的近日节律变化的规律可反映出心血管系统正常与否。近日钟基因的表达正常与否可直接影响心血管系统的生物节律,如clock基因敲除小鼠不但引起自发活动的近日节律的变化,亦导致代谢和心率等变化,甚至造成生长发育和肿瘤等疾病的发生[13-15]。研究发现节律基因Clock可能为中国汉族人群高脂血症相关基因[16]。而导致高脂血症冠状动脉硬化出现心肌缺血症状。
本研究提示Clock基因rs3840267位点与中国汉族人群心肌缺血存在关联性。通过细胞水平和动物实验研究,采用RNA干扰技术沉默小鼠体内clock基因表达,发现其血浆中Pai-1和Tm表达明显下调。PAI-1 和TM 是在凝血机制和血栓形成中发挥着重要作用的因子。PAI-1是一种单链糖蛋白,当其以活性状态存在于血浆中时与血浆波基结合素结合,形成复合物提高其稳定性,当PAI-1活性存在时能有效的发挥其生理机能,降低血液的凝血倾向及血栓形成,防止血液在血管内形成血栓[17-18]。TM是位于血管内皮细胞膜表面的糖蛋白,它具有清除凝血酶、活化血液中抗凝因子蛋白C 的作用,在抗凝因子蛋白C 等的协同作用下能促进纤维蛋白的溶解从而降低血栓形成等作用[19-20]。当PAI-1和TM二者的功能失调时,将导致血凝和抗血栓功能失调,特别是PAI-1和TM的基因表达下调时,引起表达产生的PAI-1和TM 减少,会造成抗凝血功能减弱,在血管中易形成血栓。正常情况下,PAI-1和TM的基因属于钟控基因,即二者的表达受到Clock的调控。当Clock基因表达异常时,特别是基因发生改变,如SNP等变异后,表达产生的Clock对PAI-1和TM基因调控功能受到影响,主要是造成Pai-1和Tm表达下调,生成的PAI-1和TM量下降,从而引起抗凝血功能下降,血管中的血液容易形成血栓。
通常情况下,Clock基因SNP变化是不足以引起近日节律的完全消失,但是能造成如PAI-1和TM等钟控基因的表达下调,造成血管中的血液易形成血栓;而Clock基因近日节律仍存在在凌晨自身表达下将形成的CLOCK亦下降,结果造成患者在清晨形成血栓倾向升高,导致冠脉血管狭窄,易出现清晨心绞痛发作,甚至发生血管完全被血栓阻塞致心肌梗塞等近日节律特征的变化。
4 结论
本研究显示,通过对Clock基因多态性与心肌缺血的关联性研究,发现Clock基因rs3840267位点多态性与心肌缺血存在关联。该研究为缺血性心脏病的防治提供了更进一步的实验依据。
【参考文献】
[1]Haude M.Management of Valvular Heart Disease: Esc/Eacts Guidelines 2017[J]. Herz,2017,42(8): 715-720.
[2]Maheshwari V, Barr B, and Srivastava M. Acute Valvular Heart Disease[J]. Cardiol Clin,2018,36(1): 115-127.
[3]Timmis A, Townsend N, Gale C,etal. European Society of Cardiology: Cardiovascular Disease Statistics 2017[J]. Eur Heart J,2017,38(2): 74-98.
[4]Auer R, Sidney S, Goff D,etal. Lifetime Marijuana Use and Subclinical Atherosclerosis: The Coronary Artery Risk Development in Young Adults (Cardia) Study[J]. Addiction,2017,112(11): 1904-1912.
[5]von Schantz M. Phenotypic Effects of Genetic Variability in Human Clock Genes on Circadian and Sleep Parameters[J]. J Genet,2008,87(5): 513-519.
[6]Hardin PE. The Circadian Timekeeping System of Drosophila[J]. Curr Biol,2005,15(17): 714-722.
[7]Mignot E, Takahashi JS. A Circadian Sleep Disorder Reveals a Complex Clock[J]. Cell,2007,128(1): 22-23.
[8]Maemura K, de la Monte SM, Chin MT,etal. Clif, a Novel Cycle-Like Factor, Regulates the Circadian Oscillation of Plasminogen Activator Inhibitor-1 Gene Expression[J]. J Biol Chem,2000,275(47): 36847-36851.
[9]Maemura K, Layne MD, Watanabe M,etal. Molecular Mechanisms of Morning Onset of Myocardial Infarction[J]. Ann N Y Acad Sci,2001,947(12): 398-402.
[10] Fontenot MR, Berto S, Liu Y,etal. Novel Transcriptional Networks Regulated by Clock in Human Neurons[J]. Genes Dev,2017,31(21): 2121-2135.
[11] Wang Y, Ni X, Yan J,etal. Modeling Transcriptional Co-Regulation of Mammalian Circadian Clock[J]. Math Biosci Eng,2017,14(5-6): 1447-1462.
[12] Olliac B, Ouss L, Charrier A. Suicide Attempts in Children and Adolescents: The Place of Clock Genes and Early Rhythm Dysfunction[J]. J Physiol Paris,2017,110(4): 461-466.
[13] El-Athman R, Genov NN, Mazuch J,etal. The Ink4a/Arf Locus Operates as a Regulator of the Circadian Clock Modulating Ras Activity[J]. PLoS Biol,2017,15(12): 2002940.
[14] Bering T, Carstensen MB, Rath MF. Deleting the Arntl Clock Gene in the Granular Layer of the Mouse Cerebellum: Impact on the Molecular Circadian Clockwork[J]. J Neurochem,2017,142(7): 841-856.
[15] Sun Q, Zhao Y, Yang Y,etal. Loss of the Clock Protein Per2 Shortens the Erythrocyte Life Span in Mice[J]. J Biol Chem,2017,292(30): 12679-12690.
[16] 邹晏,刘阳,鲁芳,等. 中国汉族人群近日钟基因与高脂血症的关联[J]. 西部医学,2014,26(12): 1574-1580.
[17] Oishi K, Ohkura N, Yasumoto Y,etal. Circadian Fluctuations in Circulating Plasminogen Activator Inhibitor-1 Are Independent of Feeding Cycles in Mice[J]. Chronobiol Int,2017,34(2): 254-259.
[18] Oishi K, and Ohkura N. Chronic Circadian Clock Disruption Induces Expression of the Cardiovascular Risk Factor Plasminogen Activator Inhibitor-1 in Mice[J]. Blood Coagul Fibrinolysis,2013,24(1): 106-108.
[19] Cheng S, Jiang Z, Zou Y,etal. Downregulation of Clock in Circulatory System Leads to an Enhancement of Fibrinolysis in Mice[J]. Exp Biol Med (Maywood),2011,236(9): 1078-1084.
[20] Takeda N, Maemura K, Horie S,etal. Thrombomodulin Is a Clock-Controlled Gene in Vascular Endothelial Cells[J]. J Biol Chem,2007,282(45): 32561-32567.
