洱海骨顶鸡(Fulica atra)的日间越冬行为研究
2018-07-05刘太香施香环张淑霞
徐 芳,刘太香,施香环,张淑霞
(大理大学东喜玛拉雅研究院,云南大理 671003)
活动节律与时间分配是动物行为学研究的重要内容,直接与动物的新陈代谢与能量需求相关,研究动物时间分配与活动节律有助于了解动物的生活习性、生理和生态需求。鸟类的时间分配不仅是对环境条件的一种适应〔1〕,而且是影响其活动全部因素的综合表现〔2〕。
国内外对游禽类水鸟的行为时间分配与活动规律开展了很多研究,例如小䴙䴘(Podicepsruficollis)、凤头䴙䴘(Podiceps cristatus)〔3〕、中华秋沙鸭(Mergus squamatus)〔4〕、鸳鸯(Aix galericulata)〔5〕、白眼潜鸭(Aythya nyroca)〔6-8〕、白头硬尾鸭(Oxyura leucocepha⁃la)〔9〕、小天鹅(Cygnus columbianus)〔10〕等;但有关骨顶鸡的行为数据尚比较缺乏,尤其是自然湿地中自然状态下越冬期的行为活动节律。Irwin和O'Halloran〔11〕研究了爱尔兰城市公园中人工喂食条件下的骨顶鸡越冬行为,发现游泳是人工喂食状态下的骨顶鸡日间最主要的行为类型,占全天行为的38%;桑莉莉等〔12〕对崇明东滩人工湿地(鱼蟹塘)中越冬的骨顶鸡行为进行观察,发现取食行为是主要的行为类型,占32.58%。为了增加对骨顶鸡这一物种自然状态下的生态学行为理解,我们对处于中国西南云南省大理州的洱海湿地中越冬的骨顶鸡群体日活动节律和行为时间分配进行了研究,并对其取食行为进行了重点研究,以揭示骨顶鸡这一物种对不同外界环境条件的适应机制。
骨顶鸡(Fulicaatra)隶属于鹤形目(Gruiformes)秧鸡科(Rallidae)〔13〕。骨顶鸡全身黑色,仅喙和额甲为白色;广布于欧亚大陆、印度次大陆、东南亚和澳大利亚;骨顶鸡为杂食性物种,冬季主要以沉水植物的种子、嫩芽、茎、叶等为食。骨顶鸡具有较强耐人为干扰的能力,甚至在国外公园中习惯了游人喂食面包〔11〕。骨顶鸡为云南省越冬水鸟中数量最多的物种,在各大湖泊、水库湿地多为优势物种〔14〕。大群骨顶鸡潜水觅食带来的食物扰动,可能促进同时混群活动的雁鸭类获得食物〔15〕,因此,研究骨顶鸡越冬行为生态学对云南冬季水鸟群落的保护具有重要作用。
洱海是云南省内骨顶鸡的主要越冬地之一,洱海中的骨顶鸡个体数量较大,可以到达上万只;再加上洱海中还有其他多种越冬水禽,例如赤膀鸭(Anas strepera)、赤麻鸭(Tadorna ferruginea)等物种,来自种内和种间的竞争压力较大,因此预期洱海自然状态中的骨顶鸡由于食物相对匮乏,将在取食行为时间分配上可能比人工喂食状态下的骨顶鸡占用更多的时间。本研究打算解决以下问题:(1)洱海越冬骨顶鸡的日间行为时间分配;(2)洱海越冬骨顶鸡的日间行为活动节律;(3)洱海越冬骨顶鸡的取食行为策略。
1 研究方法
1.1 研究地概况 洱海(25°36′~25°58′N,100°06′~100°18′E)位于云南省大理白族自治州境内,是一个典型的内陆断陷淡水湖泊。水位1 973.66 m时,长42.58 km,最大宽8.0 km,湖面面积249.0 km2,最大水深20.7 m,平均水深10.17 m〔16〕。洱海湖滨带全长128 km,其中西部48 km,南部8 km,东部51 km,北部11 km〔17〕。湖区属中亚热带高原季风气候,年平均气温15.0℃,1月平均气温8.5℃,极端最低气温-4.2℃,7月平均气温20.1℃,极端最高气温34.0℃。多年平均日照时数2 472 h,无霜期305 d,年均降水量1 056.6 mm,最大年降水量1 456.5 mm,最小年降水量650.2 mm,5~10月雨季降水量占全年的85%~96%〔16〕。近50年来,随着洱海流域社会经济的快速发展,进入洱海的氮、磷负荷持续增加,洱海水质持续下降,同时水位大幅波动,从而导致洱海水生态系统发生退化,沉水植物多样性下降且群落结构趋于简单化〔18〕。观察地点选择在洱海西岸大理古城附近的才村码头。
1.2 调查方法 2016年12月至2017年1月,借助Bushnell(20~20倍)单筒望远镜和奥林巴斯双筒望远镜(8倍),采用瞬时扫描法,按照从左到右、从近到远的“之”字形顺序对栖息于才村码头(25°42′54.65″N,100°11′45.05″E)附近水域的骨顶鸡群的所有可见个体进行观察,每15 min记录一次视野内所有可见个体的行为类别,调查中一人专注于扫描,另一人负责记录。在昼间(9:00—17:00时)进行观察。通过实际观察,结合已开展的游禽相关研究,将洱海越冬骨顶鸡的行为分为取食、游泳、理羽、警戒、休息及其他等6类,其中取食行为又分为表面进食、半表面觅食、表面觅食和潜水觅食4个小类。见表1。

表1 越冬骨顶鸡行为分类与定义
1.3 数据分析 将所有的扫描数据输入Excel,计算各类行为占比均值和标准差、4小类取食行为分别占取食行为总数的占比均值和标准差;从9:00到17:00以小时为单位,统计每个小时内的各类行为占比均值和标准差。所有数据分析在Excel中完成。
2 结果
于2016年的12月3日和31日,2017年的1月21日、22日和24日、3月18日共搜集6天数据,完成扫描177次,每次扫描个体数在61~253只。
2.1 日间越冬行为时间分配 洱海越冬期骨顶鸡的各类越冬行为中,有4种所占比例较高的行为:取食是最主要的行为,其次是休息、游泳和理羽,而警戒和其他行为占比很小。见图1。
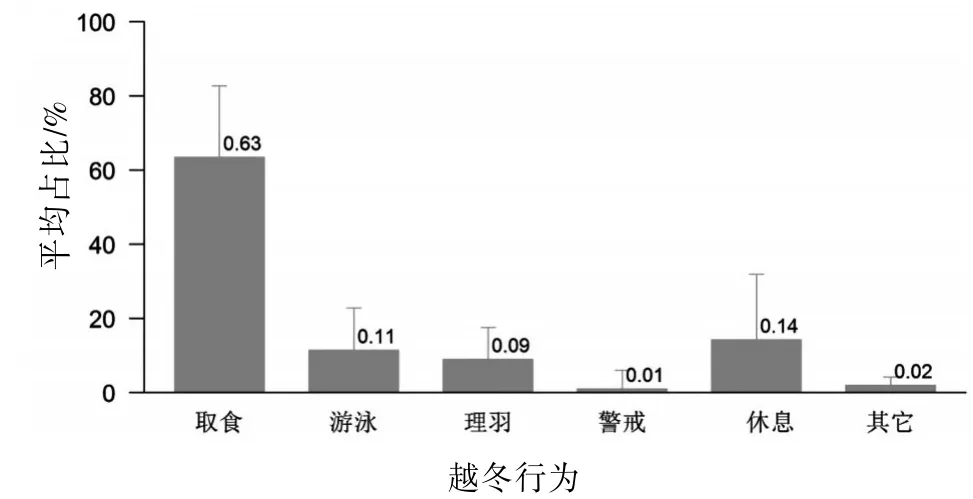
图1 洱海骨顶鸡日间各类越冬行为的占比(均值±标准差,177次扫描)
2.2 日间活动节律 虽然取食行为占时间分配比例最大,但并没有形成明显的节律(图2(a))。休息行为具有较明显的节律性,表现为两次休息小高峰期,一次在中午12点前后,另一次则在黄昏前(图2(b))。而游泳行为(图2(c))和理羽行为(图2(d))亦没有形成明显的节律。
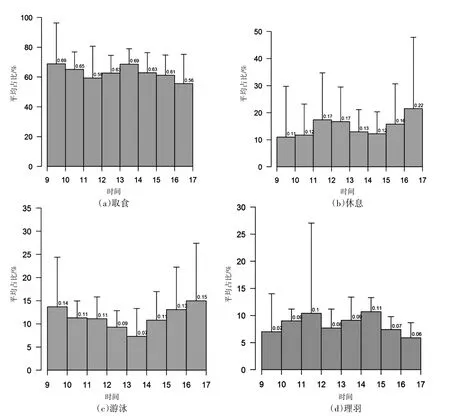
图2 洱海骨顶鸡各类越冬行为的日间变化
2.3 取食行为分析 总体来说,洱海越冬骨顶鸡在水表面进食在各种取食行为中占比最高,为其最主要的取食行为(图3)。在觅食策略上,潜水觅食比表面觅食和半表面觅食更为常见。
3 讨论
3.1 日活动时间分配 本研究中,洱海骨顶鸡在越冬期间取食行为占日活动的60%以上,是日间活动中占比最大的行为类型,这也与中国多数关于游禽的相关研究一致,但与北非越冬的白眼潜鸭表现不同〔8〕,这是由于后者具有夜间取食、昼间休息的行为特点。与爱尔兰城市公园中处于人工喂食面包状态下的越冬骨顶鸡有所区别,在城市公园人工喂食状态下,取食行为并不是日间活动的主要行为类型〔11〕,可能人类喂食的食物资源较丰富,亦可能是

图3 洱海骨顶鸡各种取食行为比较(均值±标准差,177次扫描)
人类喂食的面包提供的能量高于自然环境中的水草所造成。与人工环境相比,自然环境中的食物品质和数量相对来说可能比较缺乏,骨顶鸡需要花费较多的时间来寻找并取得食物以满足能量需求。
3.2 日间活动节律 本研究中自然湿地自然状态下的骨顶鸡具有明显的日间休息节律,即中午和黄昏时。这与中华秋沙鸭的行为节律类似,在低温环境中,中华秋沙鸭亦是从下午就开始增加休息行为〔19〕。在本研究中,骨顶鸡从14:00—15:00开始增加休息,至日落前一直处于休息状态。鸟类一般在低温环境中通过减少行为活动以降低能量消耗和热量损失〔20〕,这种节律的出现是否与低温有关,有待于未来更长越冬时间尺度的观察。
3.3 觅食和进食策略 本研究中骨顶鸡的取食行为以表面进食为主,与城市公园中骨顶鸡的取食行为一致,可能是骨顶鸡具有将潜水后啄食的植物体带至水表面进食的行为习性〔11〕,也可能是由于生理结构决定其无法完成水下进食〔21〕。潜水觅食行为多于表面觅食,可能与洱海观察点缺乏湖滩地、沿岸水较深有关:因为骨顶鸡通常根据觅食地的水深和食物分布选择不同的觅食策略,例如在浅水区域通常采取仅头部进入水中的半表面觅食策略,在深水区则多采取完全潜水的方式觅食〔22〕。本研究中洱海骨顶鸡觅食行为上表现出与前研究地点不同的特点,洱海的骨顶鸡未发现有上岸取食的现象,这可能与研究地点的湖岸带土地利用不同有关,在国外城市公园中天气不好、游人不多导致喂食量不足时,骨顶鸡会上岸取食草地上的食物,虽然这种行为发生的时候并不多;在上海崇明东滩人工湿地中,由于具备浅水区的沼泽地这一生境,骨顶鸡也会上岸取食〔12〕。而在我们的研究地点洱海才村码头,缺乏浅水湖滩地,且湖岸带已经基本被水泥硬化了,水泥路上有行人、机动车等频繁通过,并不适宜骨顶鸡上岸取食。
4 结论
在洱海自然状态下的越冬骨顶鸡,取食行为为主要日间行为类型,其次是休息;在取食行为上,以表面进食为主,没有上岸觅食行为。在日间行为节律上,本研究中骨顶鸡具有一天两次的休息高峰期,即中午和黄昏时分。
致谢:感谢中国三江并流区域生物多样性协同创新中心和云南省高校洱海流域保护与可持续发展研究重点实验室对本文提供的资助。
〔1〕ORIANS G H.The ecology of blackbird social systems〔J〕.Ecological Monographs,1961,31:285-312.
〔2〕杨晓君,杨岚.笼养绿孔雀行为时间分配的初步观察〔J〕.动物学报,1996,42(S):106-111.
〔3〕陈斌,蒋剑虹,邵明勤.小䴙䴘和风头䴙䴘越冬行为的昼间时间分配及活动节律〔J〕.湿地科学,2015,13(5):587-592.
〔4〕易国栋,杨志杰,刘宇,等.中华秋沙鸭越冬行为时间分配及日活动节律〔J〕.生态学报,2010,30(8):2228-2234.
〔5〕阮云秋.鸳鸯越冬期日活动行为时间分配的研究〔J〕.野生动物,1995,88:19-23.
〔6〕赵序茅,马鸣,张同.白眼潜鸭秋季行为时间分配及活动节律〔J〕.动物学杂志,2013,48(6):942-946.
〔7〕MUZAFFAR S B.Diurnal time-activity budgets in winter⁃ing Ferruginous PochardAythya nyrocain Tanguar Haor,Bangladesh〔J〕.Forktail,2004,20:25-27.
〔8〕HOUHAMDI M,SAMRAOUI R.Diurnal and nocturnal behaviour of Ferruginous DuckAythya nyrocaat Lac DesOiseaux,northern Algeria〔J〕.Ardeola,2008,55(1):59-69.
〔9〕赵序茅,马鸣,张同,等.白头硬尾鸭行为时间分配及日活动节律〔J〕.生态学杂志,2013,32(9):2439-2443.
〔10〕李言阔,黄建刚,李凤山,等.鄱阳湖越冬小天鹅在高水位年份的昼间时间分配和活动节律〔J〕.四川动物,2013,32(4):498-503.
〔11〕IRWIN S,O’HALLORAN J.The wintering behaviour of CootFulica AtraL.at Cork Lough,south-west Ireland〔J〕.Proceedings of the Royal Irish Academy,1997,97B(2):157-162.
〔12〕桑莉莉,葛振鸣,裴恩乐,等.崇明东滩人工湿地越冬水禽行为观察〔J〕.生态学杂志,2008,27(6):940-945.
〔13〕杨岚,文贤继,韩联宪,等.云南鸟类志:上卷(非雀形目)〔M〕.昆明:云南科技出版社,1995.
〔14〕杨岚,李恒.云南湿地〔M〕.北京:中国林业出版社,2010:533.
〔15〕EDDLEMAN W R,PATTERSON C T,KNOPF F.Inter⁃specific relationships between american coots and water⁃fowl during fall migration〔J〕.The Wilson Bulletin,1985,97(4):463-472.
〔16〕王苏民,窦鸿身.中国湖泊志〔M〕.北京:科学出版社,1998:371.
〔17〕尹延震,储昭升,赵明,等.洱海湖滨带水质的时空变化规律〔J〕.中国环境科学,2011,31(7):1192-1196.
〔18〕符辉,袁桂香,曹特,等.洱海近50a来沉水植被演替及其主要驱动因素〔J〕.湖泊科学,2013,25(6):854-861.
〔19〕曾宾宾,邵明勤,赖宏清,等.性别和温度对中华秋沙鸭越冬行为的影响〔J〕.生态学报,2013,33(12):3712-3721.
〔20〕CARACO T.Time budgeting and group size:a test of theo⁃ry〔J〕.Ecology,1979,60(3):611-617.
〔21〕郑光美.鸟类学〔M〕.北京:北京师范大学出版社,1995:55.
〔22〕CONIGLIARO M,BATTISTI C,AMORI G,et al.Diving times and pecking rates of the Eurasian Coot(Fulica atra)in different habitat types:a pilot study〔J〕.Rend Fis Acc Lincei,2011,22:47-53.
