水稻根系生长素抗性突变体的筛选及表型鉴定
2018-07-05张志昌潘伟槐严旭尹守鹏程祝宽潘建伟5
张志昌,潘伟槐,严旭,尹守鹏,程祝宽,潘建伟5*
(1.浙江师范大学化学与生命科学学院,浙江金华321004;2.铜仁职业技术学院农学院,贵州铜仁554300;3.绍兴文理学院生命科学学院,浙江绍兴312000;4.中国科学院遗传与发育生物学研究所,北京100101;5.兰州大学生命科学学院,兰州730000)
植物根系是植物体吸收水分和养分的主要器官,同时还具有固定地上部分、分解有毒物质和感知环境刺激或胁迫等生物学功能。水稻(Oryza sativa L.)是重要的单子叶模式作物,其根系主要是由初生根、不定根及其侧根组成的须根系,生长发育调控比双子叶模式植物拟南芥(Arabidopsis thaliana)的直根系复杂得多[1]。水稻生长发育受内源激素调控,主要包括生长素代谢(合成、降解)途径、生长素运输途径、生长素信号转导途径及其他激素调控[2]。生长素合成途径中的限速基因YUCCA1的超表达可以引起吲哚乙酸水平提高,促进水稻不定根的形成[3]。OsPIN1基因属于生长素极性运输蛋白PIN家族,其沉默后会抑制不定根的发育[4]。OsRAA1(Oryza sativa root architecture associated 1)能够通过泛素蛋白酶途径进行降解,OsRAA1的超表达会抑制主根生长和使不定根数目增加[5];水稻Aux/IAA基因家族OsIAA11(indole-3-acetic acids 11)基因突变能够抑制水稻侧根原基的形成[6]。
植物根系生长于土层中,其生长发育表型分析远比地上部分困难,因此在水稻中鲜有与生长素相关突变体的报道,而现有成果主要是通过拟南芥研究取得。本研究拟通过人工合成的生长素类似物2,4-二氯苯氧乙酸(2,4-dichlorophenoxyacetic acid,2,4-D)处理,从辐射诱变的水稻突变体库中筛选水稻根系生长素抗性突变体,并分析突变体的主根、不定根和侧根形成与生长对2,4-D处理的响应,鉴定其表型,以期为后续目的基因的克隆和功能鉴定奠定理论基础,以及为揭示水稻根系生长发育调控机制提供遗传学研究材料;同时,水稻作为重要的粮食作物,发育良好的根系是农作物高产、稳产的重要保障,因此,深入展开水稻根系发育调控研究具有重要的实践指导意义。
1 材料与方法
1.1 材料
为建水稻突变体库,于2012年在江苏省农业科学院原子能研究所,利用60Co-γ(诱变剂量400 Gy)辐射诱变野生型黄华占(Oryza sativa L.ssp.indica)水稻种子。经繁殖,在M2代共获得3 805个独立株系。
1.2 水稻初生根伸长对外源2,4-D处理的响应测定
参照PAN等[7]的方法,并略作改进。经ddH2O浸泡后的野生型黄华占种子4℃低温处理5 d后,于37℃条件下萌发24 h,播种于96孔板上,在0.1 mmol/LCaC12水溶液中,光照培养72 h(光照16 h/黑暗8 h,光强3×104lx,温度32℃/28℃,湿度75%)。将萌发4 d后的水稻幼苗进行生长素类似物2,4-D浓度梯度试验。选取长势一致的幼苗,每个处理组10株。2,4-D浓度梯度(0、0.01、0.05、0.1、0.5、1.0和5.0 μmol/L)用0.1 mmol/L CaCl2水溶液进行配制,分别处理水稻幼苗72 h。处理前后分别测量水稻初生根根长(root length,RL),以分析不同生长素浓度对水稻初生根伸长的抑制作用。生长素对初生根伸长的抑制率计算公式为:抑制率=1-(RL2,4-D-72h-RLCK-0h)/(RLCK-72h-RLCK-0h)。本试验至少独立重复3次,结果取平均值,用t-检验(Student’s t-test)进行显著性分析。
1.3 水稻突变体生长素抗性表型的初筛方法
本研究所用水稻突变体库共有3 805个独立株系,每个株系取25~30粒种子,经ddH2O浸泡后,4℃低温处理5 d,再于37℃条件下萌发24 h,播于96孔板中,在 0.5 μmol/L 2,4-D 水溶液(含有0.1 mmol/L CaCl2)中,光照培养 72 h(光照16 h/黑暗8 h,光强3×104lx,温度32 ℃/28 ℃,湿度75%)。以野生型黄华占为对照,测量每个株系初生根在2,4-D处理前后的根长。每个处理重复3次,结果取平均值,用t-检验进行显著性分析。在本研究中,将初生根伸长长度为野生型2倍或2倍以上的突变体株系初步定为候选生长素抗性突变体。
1.4 候选突变体的生长素抗性表型的鉴定
对经初筛后的候选突变体对生长素抗性表型进行二次鉴定,以确定是否为真正的生长素抗性突变体。从每个候选突变体株系中选取100粒饱满籽粒,经ddH2O浸泡后4℃低温处理5 d,再置于37℃条件下萌发24 h,播于96孔板中,2,4-D浓度梯度为 0、0.01、0.05、0.1、0.5、1.0 和 5.0 μmol/L(含有0.1 mmol/LCaCl2),光照培养72 h(光照16 h/黑暗8 h,光强3×104lx,温度32℃/28℃,湿度75%)。处理前后分别测量水稻初生根根长,以分析不同生长素浓度对水稻初生根伸长的抑制作用。每个处理重复3次,结果取平均值,用t-检验进行显著性分析。
2 结果与分析
2.1 水稻生长素抗性突变体筛选体系的建立
在对水稻突变体库进行生长素抗性突变体筛选之前,需要确定生长素最佳筛选浓度。本研究利用人工合成生长素类似物2,4-D不同浓度梯度对野生型水稻黄华占初生根伸长的影响进行了分析,结果如图1A所示。与空白对照相比,0.01 μmol/L 2,4-D明显促进了根的伸长,表明低浓度生长素能够促进水稻初生根的伸长。然而,从图1B可以看出,0.05、0.1、0.5、1.0和5.0 μmol/L的2,4-D均抑制了幼苗初生根的伸长,其抑制率分别为15%、33%、85%、94%和99%。这说明随着2,4-D浓度的增加,生长素对初生根伸长的影响表现为抑制作用。因此,为筛选高抗生长素突变体,本研究将抑制率为85%、浓度为0.5 μmol/L的2,4-D(P<0.001)作为筛选生长素抗性突变体的初筛浓度。

图1 不同浓度外源2,4-D处理对水稻初生根伸长的影响Fig.1 Effects of exogenous 2,4-D treatment with different concentrations on primary root elongation in rice
2.2 水稻生长素抗性突变体的初筛和二次鉴定
本研究利用0.5 μmol/L的2,4-D筛选浓度,对水稻突变体库中的3 805个突变体株系进行了初筛,共获得188个候选突变体株系,为二次筛选奠定了基础。
为验证这188个候选突变体株系是否为真正的高抗生长素突变体,利用2,4-D不同浓度梯度对其进行二次鉴定。结果表明,在188个株系中,仅4个株系真正表现为根系生长素高抗表型,分别被命名为Osarr1-1(Oryza sativa auxin resistant root 1-1)、Osarr2-1、Osarr3-1和Osarr3-2突变体。在0.01、0.1、0.5和 1.0 μmol/L 2,4-D 处理 72 h后,突变体Osarr1-1初生根伸长量高于野生型对照组(图2A和E),表明Osarr1-1初生根伸长对生长素具有较强抗性。突变体株系Osarr2-1在0.1和0.5 μmol/L 2,4-D处理组中,其初生根伸长对2,4-D处理表现出较强的生长素抗性(图2B和F)。突变体株系Osarr3-1初生根伸长对 0.01、0.05、0.1、0.5和 1.0 μmol/L 2,4-D处理均表现出极强的生长素抗性(图2C和G)。此外,由图2A~D可知,在无2,4-D处理的情况下,Osarr1-1、Osarr2-1和Osarr3-1突变体与野生型的初生根伸长完全相似(P>0.05)。

图2 水稻生长素抗性突变体的表型鉴定Fig.2 Phenotypic identification of auxin-resistant mutants in rice
2.3 外源2,4-D处理对Osarr1-1和Osarr2-1突变体根系的影响
为进一步分析2,4-D对突变体不定根和侧根的影响,本研究测定了经生长素处理后野生型和突变体不定根和侧根的数目和伸长。由图3I可知:在无2,4-D存在的情况下,Osarr1-1和Osarr2-1的不定根数目与野生型相比差异无统计学意义(P>0.05),Osarr1-1不定根伸长小于野生型,而Osarr2-1不定根伸长与野生型相似;在0.5 μmol/L 2,4-D处理下,Osarr1-1和Osarr2-1的不定根数目明显多于野生型。从图3A~D中可知:尽管2,4-D处理没有明显影响野生型不定根的形成和数目,但明显抑制了其伸长;相反,与野生型相比,2,4-D处理对Osarr1-1和Osarr2-1不定根伸长的影响相对较小,表明Osarr1-1和Osarr2-1不定根伸长对2,4-D处理不敏感。
此外,本研究还分析了2,4-D处理对侧根的影响。结果(图4A和B)表明:在无2,4-D的情况下,Osarr1-1的侧根长度明显比野生型短,但其侧根数目无明显差异(P>0.05);而Osarr2-1的侧根数目和长度与野生型相比差异均无统计学意义(P>0.05)。在0.5 μmol/L2,4-D处理后,Osarr1-1和Osarr2-1侧根长度和数目均明显高于野生型。这表明,Osarr1-1和Osarr2-1突变体根系包括初生根、不定根和侧根的形成与伸长对2,4-D处理表现为不敏感。
2.4 外源2,4-D处理对Osarr3突变体根系的影响
在对Osarr3突变体株系进行鉴定的过程中,发现有绿苗(Osarr3-1)与白化苗(Osarr3-2)分离,并且呈3∶1(35∶11≈3.18∶1)的分离比例,暗示白化苗性状由单基因突变引起。因此,我们将这些分离出来的白化苗突变体命名为Osarr3-2。由于该白化苗突变体在三叶期时的叶色可以由白转绿并继续存活繁殖,最终能获得种子,因此,本研究对Osarr3-1和Osarr3-2突变体不定根和侧根的形成与伸长同时进行了分析。
由图3I可知:在无2,4-D的情况下,与野生型相比,Osarr3-1不定根数目明显增加,但Osarr3-2则无明显差异(P>0.05);在0.5 μmol/L 2,4-D处理下,Osarr3-1和Osarr3-2不定根数目明显高于野生型。与Osarr1-1和Osarr2-1相似,Osarr3-1和Osarr3-2不定根伸长也表现为对2,4-D处理不敏感(图3F和H)。对Osarr3-1和Osarr3-2侧根的分析结果(图4)表明:在无2,4-D的情况下,Osarr3-1侧根数目和伸长与野生型相比没有差异(图4A,B和G;P>0.05),而Osarr3-2侧根形成和伸长明显受抑制(图4A,B和I)。在0.5 μmol/L 2,4-D处理组中,Osarr3-1侧根形成与伸长均高于野生型,表现出对2,4-D处理不敏感(图4A,B和H);而Osarr3-2侧根形成对2,4-D处理的敏感性与野生型相似(图4B和J;P>0.05),但其侧根伸长明显高于野生型(图4A和J)。Osarr3-1和Osarr3-2突变体在不同发育时期的表型见图5。
3 讨论
由于不同水稻品种对2,4-D的敏感性差异较大,因此,在筛选水稻突变体库之前,本研究首先测定了野生型水稻品种黄华占对2,4-D的敏感性。然后,对由60Co-γ辐射诱变的水稻突变体库进行生长素抗性初筛,共筛选到188个候选抗性株系。经生长素抗性二次鉴定后,最终获得4个生长素抗性稳定的突变株系。这一结果暗示,利用单一生长素浓度的初筛,其假阳性结果非常高;因此,二次鉴定十分必要。

图3 外源2,4-D处理对水稻不定根的影响Fig.3 Effects of exogenous 2,4-D treatment on adventitious roots of rice
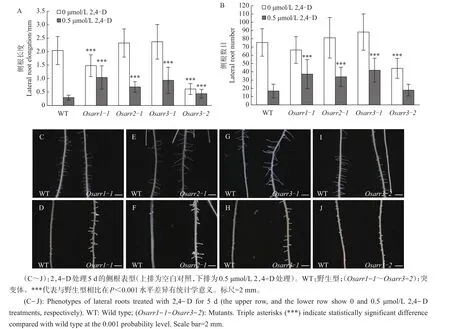
图4 外源2,4-D处理对水稻侧根的影响Fig.4 Effects of exogenous 2,4-D treatment on lateral roots of rice

图5 Osarr3-1和Osarr3-2在不同发育时期的表型Fig.5 Phenotypes of Osarr3-1 and Osarr3-2 at different developmental stages
已知生长素调控植物向性生长、逆境胁迫响应和侧生器官的形成等均是通过生长素信号途径调控下游有关基因的表达来实现的[8-9]。拟南芥生长素受体TIR1/AFB或生长素响应因子ARF的功能缺陷都会引起植株对外源生长素敏感性的改变[10-11]。在水稻中,目前已经报道的水稻生长素信号传导突变体大多与水稻不定根和侧根形成相关。如超表达OsRAA1基因能够抑制水稻初生根的伸长,但同时促进不定根的形成,其植株对外源生长素处理超敏感,以及向性生长迟缓等[5,12]。对水稻根系的遗传学分析表明,水稻生长素相关突变体如crl1/arl1表现出不定根的缺陷,OsCRL1/ARL1表达受生长素响应因子OsARF的调控,表明生长素信号转导途径在水稻不定根形成调控中具有关键性作用[13-14]。同样,OsIAA1和OsIAA11功能缺陷抑制了侧根的形成和生长,表明OsIAA1和OsIAA11参与调控水稻侧根的形成与发育[6,15]。进一步研究发现,OsIAA11通过与OsIPK2互作调控侧根的形成,OsIPK2的超表达促进了OsIAA11蛋白的稳定性,从而导致其在植株内的积累[16]。
本研究通过2,4-D处理筛选获得4个高抗生长素水稻突变体株系Osarr1-1、Osarr2-1、Osarr3-1和Osarr3-2。这些突变体根系(主根、侧根和不定根)的形成与生长对2,4-D处理均表现为不敏感,与已报道的拟南芥和水稻抗生长素突变体的表型相似。但在无2,4-D情况下,这4个突变体的不定根和侧根表型不完全相似;因此,这4个突变体很可能与生长素信号转导途径中的不同组分有关。当然,也不能完全排除与生长素生物合成途径相关[17],因为2,4-D处理对这4个突变体的不定根形成和生长有一定程度的促进作用。此外,本研究发现,Osarr3-2白化苗突变体的白化苗表型在三叶期后自动消失,推测其突变位点很可能推迟了叶绿体的发育。然而,要进一步揭示其内在调控机制,必须首先分离克隆这些突变基因。因此,今后的研究工作重点是利用图位克隆技术分离克隆这些目标基因,为解析水稻根系形成与生长调控机制提供新的观点。
:
[1] CUI P,LIU H B,RUAN S L,et al.A zinc finger protein,interacted with cyclophilin,affects root development via IAA pathway in rice.Journal of Integrative Plant Biology,2017,59(7):496-505.
[2]WANG Y D,ZHANG T,WANG R C,et al.Recent advances in auxin research in rice and their implications for crop improvement.Journal of Experimental Botany,2018,69(2/3/4):255-263.
[3]YUKO Y,NORIKO K,YOICHIM,etal.Auxin biosynthesis by the YUCCA genes in rice.Plant Physiology,2007,143(3):1362-1371.
[4] XU M,ZHU L,SHOU H X,et al.A PIN1 family gene,OsPIN1,involved in auxin-dependent adventitious root emergence and tillering in rice.Plant and Cell Physiology,2005,46(10):1674-1681.
[5] HAN Y,WANG X,JIANG J F,et al.Biochemical character of the purified OsRAA1,a novel rice protein with GTP-binding activity,and its expression pattern in Oryza sativa.Journal of Plant Physiology,2005,162(9):1057-1063.
[6] ZHU Z X,LIU Y,LIU S J,et al.A gain-of-function mutation in OsIAA11 affects lateral root development in rice.Molecular Plant,2012,5(1):154-161.
[7] PAN W H,SHOU J X,ZHOU X R,et al.Al-induced cell wall hydroxyproline-rich glycoprotein accumulation is involved in alleviating Al toxicity in rice.Acta Physiologiae Plantarum,2011,33(2):601-608.
[8] KEPINSKI S,LEYSER O.The Arabidopsis F-box protein TIR1 is an auxin receptor.Nature,2005,435(7041):446-451.
[9]DHARMASIRI N,DHARMASIRI S,ESTELLE M,et al.The F-box protein TIR1 is an auxin receptor.Nature,2005,435(7041):441-445.
[10]CALDERON VILLALOBOS L I,LEE S,DE OLIVEIRA C,et al.A combinatorial TIR1/AFB-Aux/IAA co-receptor system for differential sensing of auxin.Nature Chemical Biology,2012,8(5):477-485.
[11]KATIE G,AARON S,CRISTINA C,et al.Retraction notice to:The AFB4 auxin receptor is a negative regulator of auxin signaling in seedlings.Current Biology,2015,25(6):520-525.
[12]XU YY,CAO H,CHONG K.APC-targeted RAA1 degradation mediates the cell cycle and root development in plants.Plant Signaling&Behavior,2010,5(3):218-223.
[13]COUDERT Y,LE V A T,ADAM H,et al.Identification of crown rootless1-regulated genes in rice reveals specific and conserved elements of postembryonic root formation.New Phytologist,2015,206(1):243-254.
[14]COUDERT Y,BES M,LE T V A,et al.Transcript profiling of crown rootless1 mutant stem base reveals new elements associated with crown root development in rice.BMC Genomics,2011,12(1):387.
[15]SONG Y L,YOU J,XIONG L Z,et al.Characterization of OsIAA1 gene,a member of rice Aux/IAA family involved in auxin and brassinosteroid hormone responses and plant morphogenesis.Plant Molecular Biology,2009,70(3):297-309.
[16]CHEN Y,YANG Q F,SANG S H,et al.Rice inositol polyphosphate kinase(OsIPK2)directly interactswith OsIAA11 to regulate lateral root formation.Plant and Cell Physiology,2017,58(11):1891-1900.
[17]QIN H,ZHANG Z J,WANG J,et al.The activation of OsEIL1 on YUC8 transcription and auxin biosynthesis is required for ethylene-inhibited root elongation in rice early seedling development.PLoS Genetics,2017,13(8):e1006955.
