广西大青山西南桦人工林林下植物优势种群分布格局
2018-07-04庞圣江杨保国刘士玲贾宏炎
庞圣江 ,张 培 ,杨保国 ,刘士玲,农 友 ,贾宏炎
(1.中国林业科学研究院 热带林业实验中心,广西 凭祥 532600;2.广西友谊关森林生态系统国家定位观测研究站,广西 凭祥 532600)
种群分布格局是植物群落的重要结构特征[1],能够直接反映植物个体在水平空间的分布状况[2],是植物自身生物学特性与自然环境长期相互作用的结果[3]。由于植物对环境长期的选择与适应,群落中优势种或者伴生种分布格局,随着环境条件的改变而变化。因此,对植物群落分布格局的研究,不仅可以了解植物的生物生态学特性,揭示种群的空间分布结构以及种群与群落的动态变化趋势[4-5],亦有助于种内与种间关系的确定、种群与环境互作机制以及理解森林的协同进化过程[6],因此,种群空间分布格局一直是生态领域的研究热点之一[7-9]。
林下植物作为人工林群落的重要组成,不仅促进凋落物的分解,并且具有增强土壤养分归还的能力[10]。由于植被层次现象明显,林下植物群落中幼树、灌木、藤本和草本植物有着各自的优势种群;它们能够迅速地调整个体的形态结构,提高环境的适应能力,为人工林生态系统的长期稳定[11-12]、资源利用的最大化[13-16]等方面起着不可替代的作用。关于林下植物种群分布格局的研究报道,多以优势种群作为对象,开展不同立地、干扰措施对种群空间分布格局的影响及其生态学意义。如,Beatty研究北美糖槭Acer saccharinum-水青冈Fagus grandifolia阔叶林更新树种分布格局发现,加拿大铁杉Tsuga canadensis幼树种群对微地形的长期适应与选择产生特定微生境,导致其呈斑片状的聚集分布[17];Gálhidy等研究发现,欧洲山毛榉Fagus sylvatica林隙环境差异,改变了林内光照、土壤水分,致使草本植物优势种呈不同空间分布格局[18];王成等研究认为,带状间伐有利于促进赤松Pinus densiflora人工林天然更新和幼苗生长,且赤松幼树更新表现为聚集分布[19];樊艳荣等研究不同林下植被干扰对毛竹Phyllostachys edulis林下植物优势种群分布格局的影响,结果表明,多年未除草、劈山除草和除草剂除草对毛竹Phyllostachys edulis林植物优势种及其空间分布格局的影响是有差异的;如杉木Cunninghamia lanceolata前在两种措施中呈聚集分布,而在后者呈随机分布,地菍Melastoma dodecandrum和芒萁Dicranopteris dichotoma在3种毛竹林中均呈强烈聚集分布[3]。目前,关于不同造林模式下植物优势种空间分布格局的研究鲜见报道,由于人工配植树种发育程度的差异,为林下植物群落提供了空间异质性较大的生存环境。同时,林下植物对人工林生态系统稳定性和长期林地肥力的维持作用独特,对不同造林模式的人工林下植物优势种空间分布格局进行研究,掌握林下植物如何充分利用人工配置树种在协同进化过程空缺出的生态资源,从而使整个人工林生态系统的结构与功能趋于稳定。
西南桦Betula alnoides为桦木科Betulaceae桦木属Betula乔木树种,因其具有速生、适应性广、材质优良以及经济价值高等优良特性,已成为我国热带、南亚热带地区林业生产的重要乡土珍贵阔叶树种,现有人工林面积已逾15万hm2[20]。随着西南桦种植业的规模化发展,西南桦人工林对当地林区生态环境的恢复与保护以及木材生产已占有非常重要地位。因此,本研究选取广西大青山林区西南桦纯林、西南桦×灰木莲Manglietia glauca混交林和西南桦×马尾松Pinus massoniana混交林为研究对象,调查林下植物种类、数量和盖度等,揭示不同造林模式的西南桦人工林林下植物优势种分布格局的影响规律,为西南桦人工林的可持续经营提供参考。
1 研究方法
1.1 研究地概况
大青山林区位于广西凭祥市中国林业科学研究院热带林业实验中心(简称热林中心),地理坐标为 21°58′~ 22°19′N,106°40′~ 106°59′E。该研究区属于南亚热带季风气候区,年均温21 ℃,年均降水量约1 400 mm,主要集中于4—9月。自20世纪70年代末至今,热林中心在国内最先开展西南桦引种、驯化、良种选育和栽培等研究,不同造林模式中,包括有西南桦纯林、西南桦×灰木莲混交林和西南桦×马尾松混交林等;林下植物种类丰富,幼树以西南木荷Schima wallichii、漆树Toxicodendron delavayi、中平树Macaranga denticulata、山杜英Elaeocarpus sylvestris和枫香Liquidambar formosana等为主;常见灌木有银柴Aporosa dioica、琴叶榕Ficus pandurata、九节Psychotria asiatica、粗叶悬钩子Rubus alceifolius和三叉苦Melicope pteleifolia等;藤本植物有玉叶金花Mussaenda pubescens、毒根斑鸠菊Vernonia cumingiana、厚叶玉叶金花Mussaenda erosa和酸藤子Embelia laeta等;草本植物以弓果黍Cyrtococcum patens、五节芒Miscanthus floridulus、蔓生莠竹Microstegium fasciculatum和铁芒萁Dicranopteris linearis等为主。土壤类型为山地红壤。
1.2 样地设置与数据采集
2015年5月以来,根据热林中心营林资料以及在野外勘查现有西南桦林的基础上,采用典型样地调查法,即用森林罗盘仪设置面积为20 m×30 m标准调查样地20个,在每个样地内沿对角线的两个角和中心位置,分别设置3个5 m×5 m的样方,用于林下植被调查,包含幼树、灌木和藤本以及草本植物的名称、数量和盖度等。其中,西南桦纯林8个样地24个草本样方,西南桦×灰木莲混交林6个样地18个草本样方,西南桦×马尾松混交林6个样地18个草本样方。同时,调查记录主林层林木树高、胸径、郁闭度、海拔、坡位和坡度等,详见表1。
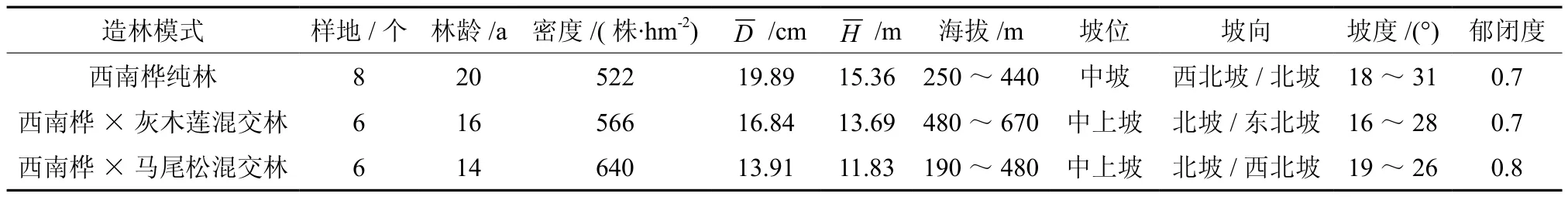
表1 研究地基本概况Table 1 The conditions of experimental sites
1.3 数据分析
本研究重要值和种群分布格局采用的计算公式,具体如下:

式中:s2为种群多度的方差,多度平均值。若DI>1,种群为聚集分布;DI=1,种群为随机分布;DI<1,种群为均匀分布。为了检验种群分布是否偏离Poisson分布,需要进行t检验。

式中:K为聚集强度,K值越大,聚集程度越低;K值越小,聚集程度越高。当K>0时,种群聚集分布;当K<0时,为均匀分布;当K→∞(一般在8以上)时,为随机分布。

式中:m表示生物个体在一个样方中的平均相邻个体数,反映了样方内生物个体的拥挤程度,数值越大聚集强度越大,表示一个个体受其他个体的拥挤效应越大[21]。
应用Excel 2007对原始数据进行初步整理,用SPSS16.0软件对西南桦纯林、西南桦×灰木莲混交林和西南桦×马尾松混交林下植物种类、数量、盖度和重要值以及分布格局等待测指标进行统计分析。
2 结果与分析
2.1 林下植物优势种及其重要值
从表2可以看出,大青山林区3种西南桦林下植物种类较为丰富,调查记录到58科107属126种,限于篇幅,仅列出林下植物优势种。其中,西南桦纯林植物种类最多(93种),西南桦×马尾松混交林次之(78种),西南桦×灰木莲混交林中物种数量最少(52种)。纯林幼树优势种为西南木荷、漆树,西南桦×灰木莲混交林中为山杜英和漆树,西南桦×马尾松混交林中为漆树、枫香和中平树;纯林灌木优势种为银柴和九节,西南桦×灰木莲混交林中为九节、粗叶悬钩子和三叉苦,西南桦×马尾松混交林中为银柴、琴叶榕和九节;纯林藤本植物优势种为玉叶金花和毒根斑鸠菊,西南桦×灰木莲混交林中为玉叶金花、毒根斑鸠菊和厚叶玉叶金花,西南桦×马尾松混交林中为玉叶金花、毒根斑鸠菊和络石;纯林草本优势种为弓果黍和五节芒,西南桦×灰木莲混交林中为弓果黍、五节芒和蔓生莠竹,西南桦×马尾松混交林中为弓果黍、五节芒、蔓生莠竹和铁芒萁。
上述3种造林模式的西南桦人工林中,林下植物优势种的重要值分别占西南桦纯林、西南桦×灰木莲混交林和西南桦×马尾松混交林的56%,67%,74.5%,表明林下植物优势种在西南桦人工林群落结构与发展过程中的重要作用和地位。3种西南桦人工林林下共有幼树优势种为漆树,灌木优势种为九节,藤本植物为玉叶金花和毒根斑鸠菊,草本植物为弓果黍和五节芒。
2.2 林下植物优势种分布格局
由表3可知,3种造林模式的西南桦人工林中,林下共有幼树优势种漆树,在西南桦×马尾松混交林的扩散系数(DI)>1,负二项参数(K=5.64)>0,聚块性指标(PAI)>1,t<t(17)呈聚集分布,但聚集程度不够明显;在纯林和西南桦×灰木莲
混交林中为均匀分布。共有灌木优势种九节和藤本植物玉叶金花,在西南桦×灰木莲混交林和西南桦×马尾松混交林中K均小于2,远远小于8,呈强烈聚集分布;二者在纯林中则表现为强度较弱的聚集分布。共有草本优势种弓果黍和五节芒均呈强烈聚集分布,K值均小于2.5。在纯林中聚集强度五节芒高于弓果黍,在西南桦×灰木莲混交林和西南桦×马尾松混交林则是弓果黍高于五节芒。说明五节芒在纯林中对空间的占有程度较高,种群处于扩张阶段;而弓果黍在后二者亦处于侵占和定居阶段。

表2 西南桦人工林下植物主要物种组成†Table 2 Species composition of the understory vegetation under Betula alnoides forests
其他非共有林下植物优势种中,纯林幼树西南木荷为均匀分布,灌木银柴为聚集分布。在西南桦×灰木莲混交林,植物优势种山杜英、粗叶悬钩子、三叉苦和叶玉叶金花均为聚集分布,但聚集程度较低;蔓生莠竹则呈强烈聚集分布。在西南桦×马尾松混交林,植物优势种铁芒萁和蔓生莠竹为强烈聚集分布,枫香和琴叶榕为聚集分布,中平树、银柴和络石均为强度较弱的聚集分布。
3种造林模式的西南桦人工林中,林下植物优势种聚集分布占所有优势种,比例分别为62.5%,81.8%,92.3%;随机或趋向随机分布的植物优势种,比例分为12.5%,9.1%,7.7%;均匀分布的植物优势种,比例分别为25%,9.1%,0%,即西南桦×马尾松混交林无均匀分布的植物优势种。

表3 西南桦人工林下植物优势种分布格局†Table 3 Distribution patterns of the understory plant species in Betula alnoides forests
3 结论与讨论
本研究结果显示,3种造林模式的西南桦人工林中,西南桦纯林植物种类最多,西南桦×马尾松混交林次之,西南桦×灰木莲混交林中物种数量最少,这与以往学者对西南桦纯林[22]、西南桦×马尾松混交林[23]以及西南桦×山桂花混交林[24]林下植物种类数量的研究结果是一致的。原因可能与西南桦为落叶乔木树种,营造纯林时,冠幅较小且不连续,冬春落叶期间,林内光照条件较好,为植物生长与繁殖提供了良好的生存空间,林下植物种类最为丰富;西南桦×马尾松混交造林,与西南桦×速生阔叶树种(灰木莲、山桂花等)比较,林冠层盖度相对较小,形成大量的林隙环境,为土壤种子库和地上植被的天然更新以及更多物种的入侵和定居创造了良好的条件,致使林下植物种类相对较多,物种多样性恢复的效果较好。由于发育时间较短,3种西南桦人工林群落结构层次尚未明显,特别是主林层均为造林树种,但林下幼树、灌木、藤本和草本植物组成极为丰富。在近自然经营的理念下,采取合理的干扰措施,促进林下幼树、灌木生长发育,将使主林层结构进一步分化,有助于形成西南桦人工林复层群落结构。
在本研究中,林下共有幼树优势种漆树,在西南桦×马尾松混交林呈聚集分布,但聚集程度不够明显,在纯林和西南桦×灰木莲混交林为均匀分布;灌木九节和藤本植物玉叶金花,在西南桦×灰木莲混交林和西南桦×马尾松混交林呈强烈聚集分布,二者在纯林聚集程度不明显;弓果黍和五节芒均在3种西南桦林内均呈强烈聚集分布。说明3种造林模式的西南桦人工林中,林下植物优势种有随机、均匀和聚集分布3种类型;其中,林下植物优势种聚集分布占所有优势种的比例达60%以上,与上官铁梁等对山西绵山植被优势种群分布格局的研究[25]以及庞圣江等对细叶云南松种群空间分布格局的研究结论基本一致[26],他们研究认为,绝大多数植物种群的分布格局类型为聚集分布。究其原因,可能与植物种群个体的生物和生态学特性密切相关,由于植物种群在自然环境分布的偶然性、地形差异和空间距离等因素,致使植物种群侵入某一领地的定居过程是非同步的点状或者非均匀的斑块状,造成植物种群的空间分布异质性。也有学者研究认为,植物种群的空间分布格局,一方面取决于植物个体的生物学特性和种间竞争关系等一系列生理生态过程,另一方面与植物群落环境,如林窗大小、土壤养分和水分资源分布等密切相关[27-28]。有关西南桦林下植物优势种分布格局的影响,尚待从植物生物学特性、种间竞争、土壤肥力以及林分环境异质性等方面开展更深层次的研究。
以往研究发现,植物种群分布格局聚集强度的变化,在某种程度上可用于表征种群的动态变化,同时,与植物群落结构的变化和演替发展相关联[29]。3种造林模式的西南桦人工林中,林下植物共有优势种漆树、九节、玉叶金花、毒根斑鸠菊、弓果黍和五节芒,在不同群落的空间分布格局以及聚集强度方面的差异;造成上述原因,与植物群落的自身特性、干扰特征、资源和生境状况的差异等有关[30-32]。由于西南桦与不同伴生树种造林时,改变了西南桦林内植物种类组成、光照、土壤肥力和水分等生境状况,致使种内以及种间竞争关系发生变化[22]。西南桦纯林呈均匀分布的林下植物优势种占所有优势种的比例最高,表明其生境相对均匀,一些植物优势种已经完成对部分空间的侵占,能够使种群长期保持稳定的状态。而西南桦×灰木莲混交林与西南桦×马尾松混交林中,强烈聚集的植物优势种占所有优势种的比例较高,说明大多数植物优势种群处于侵入与定居或者扩张阶段。聚集强度差异的结果表明,针对不同西南桦造林模式,林下植物优势种为了获取充足的资源,已经进化出不同的生存策略或者适应机制。有学者研究指出,植物优势种群处于侵入与定居或者得到进一步扩散的阶段时,具有极高聚集程度;当种群定居成功并不断地繁衍增长,侵占大部分空间以后,聚集程度就会相应地下降[29]。
综上所述,3种造林模式对西南桦林下植物优势种及其分布格局的影响存在差异;我们建议适当立地选择、减少西南桦纯林比例,宜选取马尾松作为伴生树种混交造林,采取合理的营林措施,改善林分环境,提高林分异质性,从而促进西南桦人工林健康、高效的可持续经营。
[1]康永祥,康博文,刘建军,等.陕北黄土高原文冠果群落结构及物种多样性[J].生态学报,2010, 30(16):4328-4339.
[2]樊艳荣,陈双林,林 华,等.不同林下植被干扰措施对毛竹林下植物种群分布格局的影响[J].生物多样性, 2013,21(6):709-714.
[3]彭少麟.南亚热带森林群落动态学[M].科学出版社,1996:84-89.
[4]Brown J R, Stuth J W.How herbivory affects grazing tolerant and sensitive grasses in a central Texas grassland: integrating plant response across hierarchical levels [J].Oikos, 1993, 67(2):291-297.
[5]刘振国, 李镇清.不同放牧强度下冷蒿种群小尺度空间格局[J].生态学报, 2004, 24(2):227-234.
[6]封 磊,洪 伟,吴承祯,等.杉木-拟赤杨人工混交林种内、种间竞争强度研究[J].热带亚热带植物学报,2004,12(1):46-50.
[7]玉 宝,乌吉斯古楞,王百田,等.兴安落叶松天然林不同林分结构林木水平分布格局特征研究[J].林业科学研究, 2010,23(1): 83-88.
[8]柴 勇,朱 华,孟广涛,等.云南哀牢山古茶树群落优势树种的种群结构与分布格局[J].林业科学研究, 2011, 24(3):277-284.
[9]赵中华,惠刚盈,胡艳波,等.2种类型阔叶红松林优势种群空间分布格局及其关联性[J].林业科学研究, 2011, 24(5):554-562.
[10]郭 琦,王新杰.不同混交模式杉木人工林林下植被生物量与土壤物理性质研究[J].中南林业科技大学学报, 2014, 34(5):70-74.
[11]姚茂和,盛炜彤,熊有强.杉木林下植被对立地的指示意义[J].林业科学,1992,28(3):208-212.
[12]袁正科,田育新,李锡泉,等.缓坡梯土幼林林下植被覆盖与水土流失[J].中南林业科技大学学报,2002, 22(2):21-24.
[13]Huffman D W, Moore M M.Responses of Fendler ceanothus to overstory thinning, prescribed fire, and drought in an Arizona ponderosa pine forest [J].Forest Ecology and Management,2004, 198(1-3):105-115.
[14]Taylor A H, Jang S W, Zhao L J,et al.Regeneration patterns and tree species coexistence in old-growthAbies-Piceaforests in southwestern China [J].Forest Ecology and Management, 2006,223(1-3):303-317.
[15]褚建民,卢 琦,崔向慧,等.人工林林下植被多样性研究进展[J].世界林业研究,2007,20(3):9-12.
[16]尹伟伦.全球森林与环境关系研究进展[J].森林与环境学报,2015,35(1):1-7.
[17]Beatty S W.Influence of microtopography and canopy species on spatial patterns of forest understory plants [J].Ecology, 1984,65(5):1406-1419.
[18]Gálhidy L, Mihók B, Hagyó A,et al.Effects of gap size and associated changes in light and soil moisture on the understorey vegetation of a Hungarian beech forest [J].Plant Ecology, 2006,183(1):133-145.
[19]王 成,刘继生,张美淑,等.带状间伐促进赤松人工林天然更新效果的研究——天然更新的综合效果与分布格局[J].延边大学农学学报,2004,26(2):77-80.
[20]曾 杰,郭文福,赵志刚,等.我国西南桦研究的回顾与展望[J].林业科学研究,2006,19(3):379-384.
[21]张金屯.植物种群空间分布的点格局分析[J].植物生态学报,1998,22(4):344-349.
[22]李品荣,曾觉民,陈 强,等.西南桦人工纯林与混交林群落学特征比较[J].东北林业大学学报,2007, 35(5):14-16.
[23]杨保国,刘士玲,贾宏炎,等.广西大青山西南桦人工林群落结构特征[J].广西林业科学,2016, 45(4):414-418.
[24]李莲芳,刘永刚,孟 梦,等.热带山桂花、西南桦人工幼林植物区系成分分析[J].广西植物,2007,27(3): 414-419.
[25]上官铁梁,张 峰.山西绵山植被优势种群的分布格局与种间联结的研究[J].植物科学学报,1988, 6(4):357-364.
[26]庞圣江,张 培,贾宏炎,等.细叶云南松群落物种多样性与种群空间分布格局[J].南方农业学报,2015, 46(4):645-651.
[27]刘 聪,朱教君,吴祥云,等.辽东山区次生林不同大小林窗土壤养分特征[J].东北林业大学学报,2011, 39(1):79-81.
[28]张金屯.植物种群空间分布的点格局分析[J].植物生态学报,1998,22(4):344-349.
[29]张继义,赵哈林.科尔沁沙地草地植被恢复演替进程中群落优势种群空间分布格局研究[J].生态学杂志, 2004,23(2):1-6.
[30]罗菊春,王庆锁.干扰对天然红松林植物多样性的影响[J].林业科学,1997,33(6):498-503.
[31]马云波,牛聪傑,许中旗,等.不同造林模式对铁尾矿地土壤性质的影响[J].水土保持学报,2015(3):242-248.
[32]邓娇娇,周永斌,杨立新,等.落叶松和水曲柳带状混交对土壤微生物群落功能多样性的影响[J].生态学杂志,2016,35(10): 2684-2691.
